
Abstract
How do ageing bacterial colonies generate adaptive mutants? Over a period of two months, we isolated on ageing colonies outgrowing mutants able to use a new carbon source, and sequenced their genomes. This allowed us to uncover exquisite details on the molecular mechanism behind their adaptation: most mutations were located in just a few hotspots in the genome, and over time, mutations increasingly were consistent with the involvement of 8-oxo-guanosine, formed exclusively on the transcribed strand. This work provides strong support for retromutagenesis as a general process creating adaptive mutations during ageing.
Similar content being viewed by others
Introduction
Bacteria constitute a precious biological model system for studying the molecular details of ageing and evolution. Bacterial cells defective in the MutY enzyme, responsible for removing adenine nucleotides pairing with the 8-oxo oxidized variant of guanosine (8-oxo-G), exhibit a dramatic increase in the number of adaptive mutants and Bridges proposed a model to explain this phenomenon that was later termed retromutagenesis1, 2. In this model, the process of transcription opens up the DNA double helix, enhancing the probability of mutation in the transcribed strand, but only mutations on the transcribed strand are transferred to mRNA and translate into mutant proteins that explore novel activities. Subsequently, the activity of the mutant protein may enable the cell to replicate and thereby to fix the initial adaptive mutation on both DNA strands. In agreement with the retromutagenesis model, lacZ amber mutations on the transcribed strand were recently isolated in approximately 10-fold excess over mutations on the non-transcribed strand upon treatment with the mutagen nitrous acid3. However, the prevalence of this molecular mechanism has not been studied in a non-mutator background and has not been validated in a whole genome context.
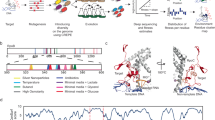
To gain new, in-depth knowledge of the mechanisms behind adaptive mutation we studied the genetic changes in a background as undisturbed as possible. For this purpose, we designed an E. coli strain incapable of fermenting maltose, plated it on rich medium with maltose, and subsequently collected all mutants starting to outgrow on colonies over the course of two months (Fig. 1).

Schematic illustration of the experimental set-up. The parental strain is incapable of using the carbon source available in the plates, but after some days on plates, mutants adapted to their environment appear. These mutants are collected and subjected to next-generation genome sequencing (NGS).
Results
Ageing colonies give rise to mutants in a non-mutator-based experimental system
Our starter strain is a derivative of the model bacterium Escherichia coli K12 MG1655 with its cyaA gene deleted, precluding synthesis of the signalling molecule cyclic AMP. As a consequence, a great many genes involved in carbon source catabolism cannot be expressed because they belong to operons requiring the cAMP Receptor Protein (CRP) complex to be activated4. These cells can grow for some time on rich media, but, after having used the accessible sources present in the available medium (maltose-MacConkey), the cells cannot grow further but remain as small colonies caught in a quiescent state.
Around four days after plating, mutants capable of fermenting maltose started to appear as red papillae-like structures on the white colonies, and these mutants continued to appear over the next two months as some cells adapted to their environment. Mutant papillae outgrew on approximately one in 200 colonies, and progressively invaded the surface of the plate (Fig. 2a) making it necessary to start with a large number of plates to be able to sample mutants at the late time points. All mutant strains were purified. They displayed a variety of phenotypes on different carbon sources (Supplementary Table 1).

When subjected to prolonged incubation, Escherichia coli K12 MG1655 cyaA- cells adapt to their environment. (a) E. coli cyaA- cease to grow after forming small white colonies, when maltose is the major carbon source. After 4–5 days of incubation, mutants start occurring as red papillae on MacConkey agar (inserted image) and continue to form over approximately 6 weeks. Mutant papillae outgrew on approximately one in 200 colonies. (b) 96 mutant papillae were isolated, phenotypically characterized and genome sequenced. Loci with less than three mutations are uniformly distributed on the circular genome as illustrated with black lines. Hotspots of genes with more than two mutations are indicated with double-sized black lines and the total number of mutations in each gene is stated within brackets.
Adaptive mutations are located in specific hotspot genes
We sequenced the genomes of 96 mutants, spanning the whole two-month period, and identified an average of four to five mutations per genome, the majority localized in a few hotspots only (Fig. 2b and Supplementary Table 2). The hotspots are located in the following genes (Fig. 2b): cmk (cytidylate kinase), crp (cyclic AMP receptor protein), malT (transcription activator of the maltose regulon transcription), rhlE (ATP-dependent DNA helicase), rpoS (sigma factor sigma 38), xseA (large subunit of exonuclease VII), and two loci that are known to be unstable in laboratory strains; e14-icd (e14 prophage inserted in the NADP-dependent isocitrate dehydrogenase gene5) and intR (Rac prophage integrase6).
From the way the experiment was constructed, it could be expected that mutations in the crp gene would account for growth on maltose in a ΔcyaA background. However, the difficulty of obtaining cAMP-independent (crp*) mutants witnessed by Jon Beckwith’s laboratory suggested that under exponential growth such mutations were very rare (of the order of 1 in 109 cells7). In light of this, the ease with which we obtained such mutations in resting colonies was utterly unexpected.
It has been repeatedly observed that in E. coli laboratory strains, the rpoS gene is often inactivated8, 9. This is also what we observed at the rpoS hotspot (point mutations in general, but also frameshifts and a deletion of the region, see Supplementary Table 2). Inactivation of the RpoS protein may have triggered an increase in age-related mutagenesis, as this transcription factor is involved in oxidative stress response during stationary phase10. However, before we understand the molecular mechanism, we can only speculate why the remaining hotspot genes are targeted.
The majority of adaptive mutations take place on the transcribed strand
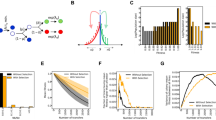
Theoretically, 12 different types of mutations are possible in DNA (A-C/G/T, C-A/G/T, G-A/C/T or T-A/C/G), but it is established that, due to respiration under stationary phase conditions, 8-oxo-G induced mutations are the most frequent to occur in the absence of an additional mutagenic process11, 12. In line with this, a dominant proportion of the mutations that were observed during the generation of the carbon-positive papillae is consistent with the involvement of 8-oxo-G (G-T and C-A transversions, 69% of the total number of missense mutations, Fig. 3a). A large proportion of the remaining mutations (28% of the total number of missense mutations identified) are likely the result of cytosine deamination (G-C to A-T transitions, Fig. 3a), another common mutational event in non-dividing cells11, 13.

8-oxoguanine formation and cytosine deamination are highly dominating on the transcribed strand in isolated papillae. (a) Assuming that all identified G-T and C-A transversions are the result of 8-oxoguanine formation, and all G-C to A-T transitions are the result of cytosine deamination, we analysed the localisation of the corresponding G- and C- residues in the 96 sequenced mutant genomes and in (b) the 594 sequenced crp sequences. Only missense mutations were analysed. (c) Upper panel: Illustration of the mechanism of retromutagenesis after 8-oxoguanine (encircled G) formation on the transcribed strand (ts, dark grey line) that enables insertion of an adenine ribonucleotide in the mRNA (green waved line). The mutated mRNA enables growth and a round of replication that permanently fixes the mutation on the coding strand (cs, light blue line) in daughter cells. Lower panel: 8-oxoguanine formation on the coding strand does not transfer to daughter cells. (d) Illustration of retromutagenesis after cytosine nucleotide deamination into uracil (encircled U), enabling base pairing with A. Upper and lower panels show result of cytosine deamination on transcribed and coding strand respectively, as detailed in (c).
Remarkably, the mutated base is highly dominantly present on the transcribed strand within gene coding sequences (84% for G transversions and 93% for C deaminations, Fig. 3a). Even more extremely, when counting in crp variants we Sanger sequenced from 594 different papillae (Supplementary Table 3), 99% of the G transversion and C deamination events had taken place on the transcribed strand (Fig. 3b). These observations are consistent with an increased mutation rate in transcribed regions (see Supplementary Table 2) and strongly support the retromutagenesis model (Fig. 3c,d)1, 3, 14 – here in the absence of a mutagen. Importantly, the extreme strand bias considerably reduces the relevance of potential residual replication and growth in the aging colonies – a heavily debated, but hard to test, part of the controversy of adaptive mutations15.
In the retromutagenesis model, mutations generated on the transcribed strand are selected for. In contrast, a so-called hitchhiking mutation is per definition not selected. Are the other hotspots selected for or hitchhikers? It is interesting to compare the specific nucleotide changes in crp that occur alone (singles) with those found only in combination (paired) with a crp* mutation: 92% (486 out of 528) of single mutations were G transversion and C deamination events on the transcribed strand, whereas only 59% (48 out of 81) of the paired mutations were of this type (Supplementary Table 3). Furthermore, the 1% G transversions and C transitions found on the non-transcribed strand (seven events in total) were all in the paired positions. Thus, in contrast to the crp* mutations, it is not possible to decipher the mechanism leading to these extra mutations in crp.
C deamination events outside crp are evenly distributed (50% on the transcribed and the non-transcribed strand). This indicates that the other C deamination events are not caused by retromutagenesis. Apart from events in crp, G transversion mutations were identified in the xseA, cmk, malT and rpoS hotspots, and 20 out of 21 cmk and 5 out of 5 xseA G transversions had taken place on the transcribed strand. This may indicate a selective advantage of these mutants, but hitchhiking is still theoretically possible because the xseA and cmk genes are transcribed from the same (+) strand as crp on the genome and thus may be fixed in the same round of replication as the crp* mutants are. In contrast, rpoS mutations must have a selective advantage: 16 out of 17 rpoS G transversion missense mutations occur on the transcribed strand, but the rpoS gene is placed on the complement (−) strand and thus retromutagenesis must have taken place independently of crp* generation. Three out of four identified G transversion mutations in the malT hotspot were placed on the non-transcribed strand and thus cannot have been selected by retromutagenesis.
Phenotypes of selected mutations created by recombineering
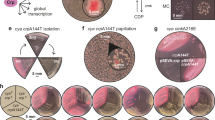
Because selection is an important aspect of retromutagenesis3 it was interesting to study the phenotypes of the hotspot mutations in isolation and in different combinations. To this end, using oligonucleotide-based recombineering16, we re-created the most frequently occurring crp (A145T and A145E), rpoS (N98K), cmk (A216E) and xseA (H456N) mutations; as well as the crp (A145E) rpos (N98K), crp (A145T) rpos (N98K), crp (A145T) cmk (A216E) and crp (A145T) xseA (H456N) double mutants. The two crp mutants showed a crp* phenotype (red on mannitol-MacConkey) in isolation and in all combinations, but were not sufficient to enable fermentation of maltose (Table 1). In contrast, mutations in the hotspots genes rpoS or xseA showed no positive impact on the fermentation of different carbon sources tested except for a slightly increased sorbitol fermentation when combining the rpoS mutation with one of the crp mutations. The crp (A145T) cmk (A216E) double mutant was able to ferment both maltose and the other tested carbon sources - in agreement with the observation that the majority of cmk mutants (23/24) isolated in the papillae experiment all co-occurred with crp mutations and exhibited a red phenotype on maltose-MacConkey. Curiously, we were unable to isolate the cmk (A216) version alone as it always co-occurred with a (spontaneous) crp mutation – possibly indicating a strongly increased selection pressure for creating a crp mutant in the presence of this cmk mutation.
Formation of adaptive mutations shows age-dependence
Time is an important factor in the development of mutant papillae and the sampling of adaptive mutants over two months enabled us to observe interesting age-related trends. Firstly, a minimum of four days of incubation is required for adaptive mutations to occur in our experimental setup. Secondly, the frequency of papillae with rpoS mutations increases over time, suggesting an increasingly selective advantage with age (Fig. 4a). Alternatively, the mechanism(s) leading to rpoS mutations are becoming more prominent over time. Thirdly, G transversion mutations in crp increases over time, paralleled by a decrease in C-deaminations (Fig. 4b).

Time-dependent trends in mutational mechanisms. (a) The frequency of rpoS mutations in isolated mutants is increasing with time. The x-axis shows days after plating. (b) Isolated mutants show a distinct mutation pattern in the crp gene. Over time, there is a steady increase in mutations originating from guanosine (likely 8-oxo-G) whereas the C-deamination associated mutations decrease.
Our observations are not consistent with a general increase in mutagenesis, as the frequency of rifampicin resistance is of the order of four mutants per 108 cells as generally observed17. Furthermore, removal of RecA, a previously suggested key actor in adaptive mutations18, displayed only a marginally negative effect on the total number of papillae formed (Supplementary Figure 1).
Discussion
In the present study, we sequenced 96 adaptive mutants isolated during a two-month period, after having arisen from a minimally perturbed genetic background. To our knowledge, a similar experiment has not been performed at this time scale previously.
Within the paradigm of our selection procedure, the identification of mutations in genes crp and rpoS was not unexpected. CRP is known to control carbon catabolism and Sabouring and Beckwith isolated the first CRP* mutant in a cya background as a strain growing on minimum maltose plates7. In the same way, RpoS controls expression of genes important for stationary phase survival. Furthermore mutations in the cognate genes have been repeatedly identified in carbon-limitation evolution experiments19,20,21. Remarkably, besides genes relevant to the process of carbon source usage, the sequencing of 96 papillae genomes revealed several hotspots and several regions of instability. The latter are all related to mobile elements. In particular the icd gene is the place of prophage e14 attachment site5, and it appears from our data that this region remains unstable in our experimental conditions. A similar observation was made when investigators explored the GASP phenotype22. In fact, the whole region is unstable as witnessed by mutations in genes ymfE, ymfI and ymfJ (see Supplementary Table 2). In addition to mutations in crp, rpoS and malT, gene cmk, coding for cytidylate kinase, emerged as a mutational hotspot. In the cmk rpsA operon23, the latter gene coding for ribosomal protein S1 in Proteobacteria and an mRNA binding protein in Firmicutes, is widely conserved in Bacteria24. Cytidylate kinase is critical for the production of CDP, the immediate precursor of dCDP that is an absolute requirement for the presence of cytosine in DNA25, and a major source of thymine as well26. The metabolism during stationary phase may require maintenance of a stable CTP pool, asking for the presence of both CMK and nucleoside diphosphokinase, but the reason for an increased mutation number in the cmk gene is unknown. However, an unexpected phenotype of the cmk gene revealed that it was involved in the control of chromosome maintenance (cmk was originally named mssA, suppressor of a cold sensitive growth of mutants in the uridylate kinase gene27), suggesting that it may belong to some important role in this domain.
The present work resulted in the selection of a very large number of CRP variants. It seems that despite the number of mutants explored (ca. 600), there is still room for further mutants to appear. The phenotype of the papillae was assayed on MacConkey medium supplemented with maltose, mannitol, glycerol or sorbitol, as well as on EMB maltose plates (Supplementary Table 1). The corresponding phenotypes are driven by the sequence of the CRP protein, but mutations elsewhere in the genome can also impact the phenotype (see for example in Supplementary Table 1 papilla 1 compared to papilla 3). The present work could be a first step in the understanding of functional networks directing carbon source utilization in stationary phase. Remarkably, previous knowledge was far from covering the whole set of what we observed. This highlights a rarely underlined process of evolution that can be explored using experiments similar to the one presented in this article.
The two maltose-positive clones that carried a wild-type crp had a weak positive phenotype and a mutation in rpoS, besides other mutations. These mutants, in contrast to the mutants in the crp gene, did not grow on minimal plates supplemented with maltose. They may have uncovered pathways allowing for growth on components of the rich MacConkey medium that the starter strain could not use.
Finally, noting that we did not find any hint of RecA involvement in the genomes we sequenced, and to validate a possible role of RecA in adaptive mutation as previously suggested18, we introduced a recA mutation in our root strain and monitored the appearance of positive papillae. Interestingly, while, overall, there does not seem to be a large decrease of the total number of papillae in a recA- background, the time course of their appearance is considerably retarded as compared to the situation observed in the cyaA- background (Supplementary Figure 1). Indeed, it takes a long time for the first papillae to appear in the recA- background, but their number keeps increasing, for up to two months, with the total number observed similar to that found in the cyaA- reference strain. In our context, recA mutants are growing more slowly than their parent counterpart. Whether this growth rate can account for the delay in papillae appearance warrants further studies.
Age was an essential factor not only for the development of adaptive mutation, but also seemingly for the mutational mechanism and we show that a highly dominating proportion of the adaptive mutations originate from guanosine on the transcribed strand - likely due to 8-oxo-G modification. This strongly supports the retromutagenesis model as a major mechanism behind adaptive mutations. Were this process generalised to multicellular organisms it would be a fertile contribution to the initiation of cancer that parallels ageing.
Methods
Bacterial strains
Because the fnr gene is a homologue of crp, we used a starter strain previously identified as MG1655 at the ECGSC1 shown to harbour a ca. 40 kb deletion in the fnr region28. The cyaA deletion was introduced with lambda Red-induced recombination29. In pilot experiments, we had noticed that the presence of a pTrc99A plasmid (Pharmacia Biotech) carrying the E. coli tig gene (PCR amplified with the oligonucleotides CATGCCATGGTGAGGTAACAAGATGCAAGTTTC 3′ and 5′ CGCGGATCCAATTACGCCTGCTGGTTCATC 3′ and cloned into the NcoI and BamHI restriction sites) produced twice as many mutants that in absence of the tig plasmid, typically allowing us to recover around 200 mutants in each two months experiment, a figure quite convenient to get significant observations. Thus K12 Mg1655 (cyaA::cat delta fnr, pTrctig) is our starter strain. The recA deletion was from the Keio collections30 that was used to prepare a P1 lysate by standard procedures.
Papillae formation assay
For growth on plates the lactose-free MacConkey medium was used31 (Difco MacConkey Agar Base) supplemented with 1% carbohydrate (maltose in most experiments) or glycerol and chloramphenicol, 5 mg/litre, ampicillin, 100 mg/litre, and 1 mM IPTG. For subsequent 3-times purification of papillae, chloramphenicol, ampicillin and IPTG were omitted from the plates. To obtain isolated colonies on the plates, early stationary-phase grown bacteria from the LB liquid culture (37 °C) were diluted in sterile water containing 9 g/l sodium chloride, to the concentration of 2.5 × 104 bacteria/ml and 100 µl of the bacterial suspension was spread onto MacConkey plates. The plates were subsequently placed in plastic boxes containing beakers with water to ensure constant humidity and placed for the duration of the experiment in an incubator at 37 °C. To test for mutator phenotypes, rifampicin agar plates were prepared using the LB medium agar supplemented with 100 mg/litre rifampicin prepared in methanol. At all times the plates were wrapped in aluminium foil for protection from light.
Genome sequencing
The genomic libraries were generated using the TruSeq®Nano DNA LT Sample Preparation Kit (Illumina Inc.). 498 independent papillae were selected for amplification of the crp region by direct colony PCR. The PCR reaction was made using Red-Taq polymerase (Sigma-Aldrich) according to the manufacturer’s instructions (hybridisation at 55 °C) with the following primers: forward primer 5′TTTCGGCAATCCAGAGACAGC3′ and reverse primer 5′AACATAGCACCAGCGTTTGTCG3′. The amplified crp regions were sequenced by Sanger method with two primers: forward 5′TTATCTGGCTCTGGAGAAAGC3′ and reverse primer 5′TCGAAGTGCATAGTTGATATCGG3′.
Genome engineering
K12 Mg1655 (cyaA::cat delta fnr) was transformed with plasmid pMA716 and recombineering performed using the oligonucleotides 1–5 (Supplementary Methods) was carried out as described by Sandberg et al.32 with three exceptions: (1) overnight LB liquid cultures were supplemented with 0.4% glucose to preclude starvation, (2) 1 mM cAMP (Sigma-Aldrich A9501) was supplemented at time of induction, and (3) cells were spread on MacConkey-mannitol plates (1% mannitol, 5 ug/ml chloramphenicol, 50 ug/ml ampicillin). Two rounds of recombineering were applied for crp A145T, crp A145E, and xseA H456N and four rounds for cmk A216E and rpoS N98K. Colonies were screened for the desired point mutations by sequencing. Positive clones were either streaked repeatedly on MacConkey-mannitol plates without ampicillin until the pMA7 plasmid was lost or inoculated in liquid culture to carry out recombineering of new targets as above (2 rounds for both crp A145E into rpoS N98K background and for crp A145T into xseA H456N background). The MacConkey medium was applied to ensure that only the dominant phenotype was propagated into later experiments and thus to prevent development of and contamination by unwanted mutations in the background. The crp A145T mutations occurred spontaneously in rpoS N98K cells and in cmk A216E. We were unable to obtain the cmk A216E background without spontaneous crp and other mutations.
References
Bridges, B. A. mutY ‘directs’ mutation? Nature 375, 741 (1995).
Doetsch, P. W. Translesion synthesis by RNA polymerases: occurrence and biological implications for transcriptional mutagenesis. Mutat. Res. 510, 131–140 (2002).
Morreall, J. et al. Evidence for Retromutagenesis as a Mechanism for Adaptive Mutation in Escherichia coli. Plos Genet 11, 1–12 (2015).
You, C. et al. Coordination of bacterial proteome with metabolism by cyclic AMP signalling. Nature 500, 301–306 (2013).
Hill, C. W., Gray, J. A. & Brody, H. Use of the isocitrate dehydrogenase structural gene for attachment of e14 in Escherichia coli K-12. J Bacteriol 171, 4083–4084 (1989).
Casjens, S. Prophages and bacterial genomics: What have we learned so far? Mol. Microbiol. 49, 277–300 (2003).
Sabourin, D. & Beckwith, J. Deletion of the Escherichia coli crp gene. J Bacteriol 122, 338–340 (1975).
Saxer, G. et al. Mutations in global regulators lead to metabolic selection during adaptation to complex environments. Plos Genet 10, e1004872 (2014).
Bleibtreu, A. et al. The rpoS gene is predominantly inactivated during laboratory storage and undergoes source-sink evolution in Escherichia coli species. J Bacteriol 196, 4276–4284 (2014).
Nyström, T. The free-radical hypothesis of aging goes prokaryotic. Cell. Mol. Life Sci. 60, 1333–1341 (2003).
Hall, B. G. Spectrum of mutations that occur under selective and non-selective conditions in E. coli. Genetica 84, 73–76 (1991).
Bridges, B. A. Mutation in resting cells: the role of endogenous DNA damage. Cancer Surv 28, 155–167 (1996).
Brégeon, D., Doddridge, Z. A., You, H. J., Weiss, B. & Doetsch, P. W. Transcriptional mutagenesis induced by uracil and 8-oxoguanine in Escherichia coli. Mol. Cell 12, 959–970 (2003).
Davis, B. D. Transcriptional bias: a non-Lamarckian mechanism for substrate-induced mutations. Proc Natl Acad Sci USA 86, 5005–5009 (1989).
Maisnier-Patin, S. & Roth, J. R. The origin of mutants under selection: How natural selection mimics mutability (adaptive mutation). Cold Spring Harb Perspect Biol 2015;7:a018176 (2015).
Lennen, R. M. et al. Transient overexpression of DNA adenine methylase enables efficient and mobile genome engineering with reduced off-target effects. Nucleic Acids Res, doi: gkv1090[pii]10.1093/nar/gkv1090 (2015).
Jin, D. J. & Gross, C. A. Mapping and sequencing of mutations in the Escherichia coli rpoB gene that lead to rifampicin resistance. J. Mol. Biol. 202, 45–58 (1988).
Taddei, F., Halliday, J. A., Matic, I. & Radman, M. Genetic analysis of mutagenesis in aging Escherichia coli colonies. Mol Gen Genet 256, 277–281 (1997).
Lombardo, M.-J., Aponyi, I. & Rosenberg, S. M. General stress response regulator RpoS in adaptive mutation and amplification in Escherichia coli. Genetics 166, 669–680 (2004).
Kinnersley, M. A., Holben, W. E. & Rosenzweig, F. E unibus plurum: Genomic analysis of an experimentally evolved polymorphism in Escherichia coli. PLoS Genet. 5 (2009).
Adams, J. & Rosenzweig, F. Experimental microbial evolution: history and conceptual underpinnings. Genomics 104, 393–398 (2014).
Zhang, Q. et al. You cannot tell a book by looking at the cover: Cryptic complexity in bacterial evolution. Biomicrofluidics 8, 52004 (2014).
Fricke, J., Neuhard, J., Kelln, R. A. & Pedersen, S. The cmk gene encoding cytidine monophosphate kinase is located in the rpsA operon and is required for normal replication rate in Escherichia coli. J Bacteriol 177, 517–523 (1995).
Nitschke, P. et al. Indigo: a World-Wide-Web review of genomes and gene functions. FEMS Microbiol Rev 22, 207–227 (1998).
Danchin, A. Comparison between the Escherichia coli and Bacillus subtilis genomes suggests that a major function of polynucleotide phosphorylase is to synthesize CDP. DNA Res 4, 9–18 (1997).
Weiss, B. & Wang, L. De novo synthesis of thymidylate via deoxycytidine in dcd (dCTP deaminase) mutants of Escherichia coli. J Bacteriol 176, 2194–2199 (1994).
Yamanaka, K., Ogura, T., Koonin, E. V., Niki, H. & Hiraga, S. Multicopy suppressors, mssA and mssB, of an smbA mutation of Escherichia coli. Mol Gen Genet 243, 9–16 (1994).
Soupene, E. et al. Physiological studies of Escherichia coli strain MG1655: growth defects and apparent cross-regulation of gene expression. J Bacteriol 185, 5611–5626 (2003).
Datta, S., Costantino, N. & Court, D. L. A set of recombineering plasmids for gram-negative bacteria. Gene 379, 109–115 (2006).
Baba, T. et al. Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: the Keio collection. Mol Syst Biol 2, 2006 0008 (2006).
MacConkey, A. Lactose-fermenting bacteria in feces. J Hyg 5, 333–378 (1905).
Sandberg, T. E. et al. Evolution of Escherichia coli to 42 °C and subsequent genetic engineering reveals adaptive mechanisms and novel mutations. Mol. Biol. Evol. 31, 2647–2662 (2014).
Acknowledgements
This work was supported by The Novo Nordisk Foundation, AMAbiotics SAS and a PhD grant from the People Programme (Marie Curie Actions) of the European Union’s Seventh Framework Programme [FP7-People-2012-ITN], under grant agreement No. 317058, “BACTORY”. We thank Ida Lauritsen for technical assistance, and Anna Koza and Ida Bonde for their assistance with NGS.
Author Contributions
A.D. and M.H.H.N. designed the experiments, supervised the work and wrote the manuscript. A.S., S.W. and E.C.F. carried out the experiments, contributed to the manuscript and prepared the figures.
Competing Interests
The authors declare no competing financial interests.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Sekowska, A., Wendel, S., Fischer, E.C. et al. Generation of mutation hotspots in ageing bacterial colonies. Sci Rep 6, 2 (2016). https://doi.org/10.1038/s41598-016-0005-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-016-0005-4
This article is cited by
-
Simple solution processed spin switching in FeCo/Rubrene/NiFe spin valve device
Indian Journal of Physics (2024)
-
cAMP Activation of the cAMP Receptor Protein, a Model Bacterial Transcription Factor
Journal of Microbiology (2023)
-
Liquid Phase Exfoliation and Characterization of Few Layer MoS2 and WS2 Nanosheets as Channel Material in Field Effect Transistor
Transactions on Electrical and Electronic Materials (2023)
-
Slaked lime improves growth, antioxidant capacity and reduces Cd accumulation of peanut (Arachis hypogaea L.) under Cd stress
Scientific Reports (2022)
-
Multi-dimensional-double-spiral (MDDS) inertial microfluidic platform for sperm isolation directly from the raw semen sample
Scientific Reports (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
