
Abstract
The history of the life–Earth system can be divided into five ‘energetic’ epochs, each featuring the evolution of life forms that can exploit a new source of energy. These sources are: geochemical energy, sunlight, oxygen, flesh and fire. The first two were present at the start, but oxygen, flesh and fire are all consequences of evolutionary events. Since no category of energy source has disappeared, this has, over time, resulted in an expanding realm of the sources of energy available to living organisms and a concomitant increase in the diversity and complexity of ecosystems. These energy expansions have also mediated the transformation of key aspects of the planetary environment, which have in turn mediated the future course of evolutionary change. Using energy as a lens thus illuminates patterns in the entwined histories of life and Earth, and may also provide a framework for considering the potential trajectories of life–planet systems elsewhere.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout

Similar content being viewed by others
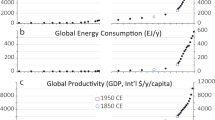
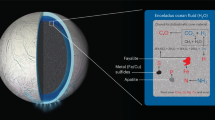

References
Thauer, R. K., Jungermann, K. & Decker, K. Energy conservation in chemotrophic anaerobic bacteria. Bacteriol. Rev. 41, 100–180 (1977).
Amend, J. P. & Shock, E. L. Energetics of overall metabolic reactions of thermophilic and hyperthermophilic Archaea and Bacteria. FEMS Microbiol. Rev. 25, 175–243 (2001).
Vernadsky, W. La Biosphère (Félix Alcan, 1929).
Cloud, P. E. Atmospheric and hydrospheric evolution on the primitive earth. Science 160, 729–736 (1968).
Smil, V. General Energetics: Energy in the Biosphere and Civilization (John Wiley and Sons, 1991).
Lenton, T. & Watson, A. Revolutions that Made the Earth (Oxford Univ. Press, 2011).
Lenton, T. M., Pichler, P.-P. & Weisz, H. Revolutions in energy input and material cycling in Earth history and human history. Earth Syst. Dynam. 7, 353–370 (2016).
Amend, J. P., McCollom, T. M., Hentscher, M. & Bach, W. Catabolic and anabolic energy for chemolithoautotrophs in deep-sea hydrothermal systems hosted in different rock types. Geochim. Cosmochim. Acta 75, 5736–5748 (2011).
Mayhew, L. E., Ellison, E. T., McCollom, T. M., Trainor, T. P. & Templeton, A. S. Hydrogen generation from low-temperature water–rock reactions. Nat. Geosci. 6, 478–484 (2013).
McCollom, T. M. & Seewald, J. S. Serpentinites, hydrogen, and life. Elements 9, 129–134 (2013).
McDermott, J. M., Seewald, J. S., German, C. R. & Sylva, S. P. Pathways for abiotic organic synthesis at submarine hydrothermal fields. Proc. Natl Acad. Sci. USA 112, 7668–7672 (2015).
Russell, M. J., Hall, A. J. & Martin, W. Serpentinization as a source of energy at the origin of life. Geobiology 8, 355–371 (2010).
Lane, N., Allen, J. F. & Martin, W. How did LUCA make a living? Chemiosmosis in the origin of life. BioEssays 32, 271–280 (2010).
Dibrova, D. V., Chudetsky, M. Y., Galperin, M. Y., Koonin, E. V. & Mulkidjanian, A. Y. The role of energy in the emergence of biology from chemistry. Orig. Life Evol. Biosph. 42, 459–468 (2012).
Sousa, F. L. et al. Early bioenergetic evolution. Phil. Trans. R. Soc. Lond. B 368, 20130088 (2013).
Weiss, M. C. et al. The physiology and habitat of the last universal common ancestor. Nat. Microbiol. 1, 16116 (2016).
Ferry, J. G. & House, C. H. The stepwise evolution of early life driven by energy conservation. Mol. Biol. Evol. 23, 1286–1292 (2006).
Rosing, M. 13C-depleted carbon microparticles in >3700-Ma sea-floor sedimentary rocks from west Greenland. Science 283, 674–676 (1999).
Pecoits, E. et al. Atmospheric hydrogen peroxide and Eoarchean iron formations. Geobiology 13, 1–14 (2015).
Bell, E. A., Boehnke, P., Harrison, T. M. & Mao, W. L. Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon. Proc. Natl Acad. Sci. USA 112, 14518–14521 (2015).
Nutman, A. P., Bennett, V. C., Friend, C. R. L., Van Kranendonk, M. J. & Chivas, A. R. Rapid emergence of life shown by discovery of 3,700-million-year-old microbial structures. Nature 537, 535–538 (2016).
Dodd, M. S. et al. Evidence for early life in Earth's oldest hydrothermal vent precipitates. Nature 543, 60–64 (2017).
Papineau, D. Mineral environments on the earliest Earth. Elements 6, 25–30 (2010).
Schönheit, P., Buckel, W. & Martin, W. F. On the origin of heterotrophy. Trends Microbiol. 24, 12–25 (2016).
Catling, D. C. in Treatise on Geochemistry 2nd edn (eds Holland, H. & Turekian, K. ) 6, 177–195 (Elsevier, 2014).
Nisbet, E. G. & Fowler, C. M. R. Archaean metabolic evolution of microbial mats. Proc. R. Soc. Lond. B 266, 2375–2382 (1999).
Brazelton, W. J., Mehta, M. P., Kelley, D. S. & Baross, J. A. Physiological differentiation within a single-species biofilm fueled by serpentinization. mBio 2, e00127–11 (2011).
Schink, B. Energetics of syntrophic cooperation in methanogenic degradation. Microbiol. Mol. Biol. Rev. 61, 262–280 (1997).
Nealson, K. H. & Conrad, P. G. Life: past, present and future. Phil. Trans. R. Soc. Lond. B 354, 1923–1939 (1999).
Fenchel, T. & Finlay, B. J. Ecology and Evolution in Anoxic Worlds (Oxford Univ. Press, 1995).
Forterre, P. & Prangishvili, D. The origin of viruses. Res. Microbiol. 160, 466–472 (2009).
Kharecha, P., Kasting, J. & Siefert, J. A coupled atmosphere–ecosystem model of the early Archean Earth. Geobiology 3, 53–76 (2005).
Canfield, D. E., Rosing, M. T. & Bjerrum, C. Early anaerobic metabolisms. Phil. Trans. R. Soc. Lond. B 361, 1819–1836 (2006).
Sleep, N. H. & Bird, D. K. Niches of the pre-photosynthetic biosphere and geologic preservation of Earth's earliest ecology. Geobiology 5, 101–117 (2007).
Fischer, W. W., Hemp, J. & Johnson, J. E. Evolution of oxygenic photosynthesis. Annu. Rev. Earth Planet. Sci. 44, 647–683 (2016).
Tice, M. M. & Lowe, D. R. Hydrogen-based carbon fixation in the earliest known photosynthetic organisms. Geology 34, 37–40 (2006).
Hohmann-Marriott, M. F. & Blankenship, R. E. Evolution of photosynthesis. Annu. Rev. Plant Biol. 62, 515–548 (2011).
Zubkov, M. V. Photoheterotrophy in marine prokaryotes. J. Plankton Res. 31, 933–938 (2009).
Bosak, T., Greene, S. E. & Newman, D. K. A likely role for anoxygenic photosynthetic microbes in the formation of ancient stromatolites. Geobiology 5, 119–126 (2007).
Ueno, Y., Yamada, K., Yoshida, N., Maruyama, S. & Isozaki, Y. Evidence from fluid inclusions for microbial methanogenesis in the early Archaean era. Nature 440, 516–519 (2006).
Arndt, N. T. & Nisbet, E. G. Processes on the young Earth and the habitats of early Life. Annu. Rev. Earth Planet. Sci. 40, 521–549 (2012).
Sagan, C. & Mullen, G. Earth and Mars: evolution of atmospheres and surface temperatures. Science 177, 52–56 (1972).
Haqq-Misra, J. D., Domagal-Goldman, S. D., Kasting, P. J. & Kasting, J. F. A revised, hazy methane greenhouse for the Archean Earth. Astrobiology 8, 1127–1137 (2008).
Catling, D. C. & Claire, M. W. How Earth's atmosphere evolved to an oxic state: a status report. Earth Planet. Sci. Lett. 237, 1–20 (2005).
Draganić, I. G. Radiolysis of water: a look at its origin and occurrence in the nature. Radiat. Phys. Chem. 72, 181–186 (2005).
Bekker, A. et al. Dating the rise of atmospheric oxygen. Nature 427, 117–120 (2004).
Lyons, T. W., Reinhard, C. T. & Planavsky, N. J. The rise of oxygen in Earth's early ocean and atmosphere. Nature 506, 307–315 (2014).
Hamilton, T. L., Bryant, D. A. & Macalady, J. L. The role of biology in planetary evolution: cyanobacterial primary production in low-oxygen Proterozoic oceans. Environ. Microbiol. 18, 325–340 (2016).
MacGregor, A. M. The problem of the Precambrian atmosphere. S. Afr. J. Sci. 24, 155–172 (1927).
Tomitani, A., Knoll, A. H., Cavanaugh, C. M. & Ohno, T. The evolutionary diversification of cyanobacteria: molecular–phylogenetic and paleontological perspectives. Proc. Natl Acad. Sci. USA 103, 5442–5447 (2006).
Bosak, T., Liang, B., Sim, M. S. & Petroff, A. P. Morphological record of oxygenic photosynthesis in conical stromatolites. Proc. Natl Acad. Sci. USA 106, 10939–10943 (2009).
Farquhar, J., Zerkle, A. L. & Bekker, A. Geological constraints on the origin of oxygenic photosynthesis. Photosynth. Res. 107, 11–36 (2011).
Kasting, J. F. What caused the rise of atmospheric O2? Chem. Geol. 362, 13–25 (2013).
Goldblatt, C., Lenton, T. M. & Watson, A. J. Bistability of atmospheric oxygen and the Great Oxidation. Nature 443, 683–686 (2006).
Papineau, D., Walker, J. J., Mojzsis, S. J. & Pace, N. R. Composition and structure of microbial communities from stromatolites of Hamelin Pool in Shark Bay, Western Australia. Appl. Environ. Microbiol. 71, 4822–4832 (2005).
Watanabe, Y., Martini, J. E. & Ohmoto, H. Geochemical evidence for terrestrial ecosystems 2.6 billion years ago. Nature 408, 574–578 (2000).
Stüeken, E. E., Catling, D. C. & Buick, R. Contributions to late Archaean sulphur cycling by life on land. Nat. Geosci. 5, 722–725 (2012).
Kasting, J. F. & Catling, D. Evolution of a habitable planet. Annu. Rev. Astron. Astrophys. 41, 429–463 (2003).
Claire, M., Catling, D. C. & Zahnle, K. J. Biogeochemical modelling of the rise in atmospheric oxygen. Geobiology 4, 239–269 (2006).
Sverjensky, D. A. & Lee, N. The Great Oxidation Event and mineral diversification. Elements 6, 31–36 (2010).
Hazen, R. M. et al. Mineral evolution. Am. Mineral. 93, 1693–1720 (2008).
Harrison, J. P. et al. Aerobically respiring prokaryotic strains exhibit a broader temperature-pH-salinity space for cell division than anaerobically respiring and fermentative strains. J. R. Soc. Interface 12, 20150658 (2015).
Melezhik, V. A. et al. in Reading the Archive of Earth's Oxygenation (eds Melezhik, V. A. et al.) 1059–1109 (Springer, 2013).
Zahnle, K., Claire, M. & Catling, D. The loss of mass-independent fractionation in sulfur due to a Palaeoproterozoic collapse of atmospheric methane. Geobiology 4, 271–283 (2006).
Daines, S. J. & Lenton, T. M. The effect of widespread early aerobic marine ecosystems on methane cycling and the Great Oxidation. Earth Planet. Sci. Lett. 434, 42–51 (2016).
Catling, D. C., Glein, C. R., Zahnle, K. J. & McKay, C. P. Why O2 is required by complex life on habitable planets and the concept of planetary “oxygenation time”. Astrobiology 5, 415–438 (2005).
Haqq-Misra, J., Kasting, J. F. & Lee, S. Availability of O2 and H2O2 on pre-photosynthetic Earth. Astrobiology 11, 293–302 (2011).
Olson, S. L., Kump, L. R. & Kasting, J. F. Quantifying the areal extent and dissolved oxygen concentrations of Archean oxygen oases. Chem. Geol. 362, 35–43 (2013).
Lenton, T. M. & Daines, S. J. Biogeochemical transformations in the history of the ocean. Annu. Rev. Mar. Sci. 9, 31–58 (2017).
Naqui, A., Chance, B. & Cadenas, E. Reactive oxygen intermediates in biochemistry. Annu. Rev. Biochem. 55, 137–166 (1986).
Imlay, J. A. The molecular mechanisms and physiological consequences of oxidative stress: lessons from a model bacterium. Nat. Rev. Microbiol. 11, 443–454 (2013).
Towe, K. M. Oxygen-collagen priority and the early metazoan fossil record. Proc. Natl Acad. Sci. USA 65, 781–788 (1970).
Williams, T. A., Foster, P. G., Cox, C. J. & Embley, T. M. An archaeal origin of eukaryotes supports only two primary domains of life. Nature 504, 231–236 (2013).
Martin, W. F., Garg, S. & Zimorski, V. Endosymbiotic theories for eukaryote origin. Phil. Trans. R. Soc. Lond. B 370, 20140330 (2015).
Keeling, P. J. The number, speed, and impact of plastid endosymbioses in eukaryotic evolution. Annu. Rev. Plant Biol. 64, 583–607 (2013).
Butterfield, N. J. Bangiomorpha pubescens n. gen., n. sp.: implications for the evolution of sex, multicellularity, and the Mesoproterozoic/Neoproterozoic radiation of eukaryotes. Paleobiology 26, 386–404 (2000).
Knoll, A. H. Paleobiological perspectives on early eukaryotic evolution. Cold Spring Harb. Perspect. Biol. 6, a016121 (2014).
Müller, M. et al. Biochemistry and evolution of anaerobic energy metabolism in eukaryotes. Microbiol. Mol. Biol. Rev. 76, 444–495 (2012).
Martin, W. & Müller, M. The hydrogen hypothesis for the first eukaryote. Nature 392, 37–41 (1998).
van der Giezen, M. & Lenton, T. M. The rise of oxygen and complex life. J. Eukaryot. Microbiol. 59, 111–113 (2012).
Lane, N. & Martin, W. The energetics of genome complexity. Nature 467, 929–934 (2010).
Douglas, A. E. Symbiosis as a general principle in eukaryotic evolution. Cold Spring Harb. Perspect. Biol. 6, a016113 (2014).
Yutin, N., Wolf, M. Y., Wolf, Y. I. & Koonin, E. V. The origins of phagocytosis and eukaryogenesis. Biol. Direct 4, 9 (2009).
Narbonne, G. M. The Ediacara biota: Neoproterozoic origin of animals and their ecosystems. Annu. Rev. Earth Planet. Sci. 33, 421–442 (2005).
Payne, J. L. et al. Two-phase increase in the maximum size of life over 3.5 billion years reflects biological innovation and environmental opportunity. Proc. Natl Acad. Sci. USA 106, 24–27 (2009).
Stanley, S. M. An ecological theory for the sudden origin of multicellular life in the late Precambrian. Proc. Natl Acad. Sci. USA 70, 1486–1489 (1973).
Butterfield, N. J. Macroevolution and macroecology through deep time. Palaeontology 50, 41–55 (2007).
Bengtson, S. Origins and early evolution of predation. Paleontol. Soc. Papers 8, 289–318 (2002).
Porter, S. M. & Knoll, A. H. Testate amoebae in the Neoproterozoic Era: evidence from vase-shaped microfossils in the Chuar Group, Grand Canyon. Paleobiology 26, 360–385 (2000).
Porter, S. The rise of predators. Geology 39, 607–608 (2011).
Cohen, P. A. & Knoll, A. H. Scale microfossils from the mid-Neoproterozoic Fifteenmile Group, Yukon Territory. J. Paleontol. 86, 775–800 (2012).
Knoll, A. H. & Lahr, D. J. G. in Multicellularity: Origins and Evolution (eds Niklas, K. J. & Neumann, S. D. ) 3–16 (MIT Press, 2016).
Rokas, A. The origins of multicellularity and the early history of the genetic toolkit for animal development. Annu. Rev. Genet. 42, 235–251 (2008).
Erwin, D. H. et al. The Cambrian conundrum: early divergence and later ecological success in the early history of animals. Science 334, 1091–1097 (2011).
Parfrey, L. W., Lahr, D., Knoll, A. H. & Katz, L. A. Estimating the timing of early eukaryotic diversification with multigene molecular clocks. Proc. Natl Acad. Sci. USA 108, 13624–13629 (2011).
Buschmann, H., Keller, M., Porret, N., Dietz, H. & Edwards, P. J. The effect of slug grazing on vegetation development and plant species diversity in an experimental grassland. Funct. Ecol. 19, 291–298 (2005).
Hutchinson, G. E. The paradox of the plankton. Am. Nat. 95, 137–145 (1961).
Ripple, W. J. & Beschta, R. L. Trophic cascades in Yellowstone: the first 15 years after wolf reintroduction. Biol. Conserv. 145, 205–213 (2012).
Butterfield, N. J. Animals and the invention of the Phanerozoic Earth system. Trends Ecol. Evol. 26, 81–87 (2011).
Vermeij, G. J. The origin of skeletons. Palaios 4, 585–589 (1989).
Baumgartner, P. O. Mesozoic radiolarites–accumulation as a function of sea surface fertility on Tethyan margins and in ocean basins. Sedimentology 60, 292–318 (2013).
Kiessling, W. Towards an unbiased estimate of fluctuations in reef abundance and volume during the Phanerozoic. Biogeosciences 3, 15–27 (2006).
Kidwell, S. M. & Brenchley, P. J. Patterns in bioclastic accumulation through the Phanerozoic: Changes in input or in destruction? Geology 22, 1139–1143 (1994).
Stanley, S. M. Influence of seawater chemistry on biomineralization throughout Phanerozoic time: Paleontological and experimental evidence. Palaeogeogr. Palaeoclimatol. Palaeoecol. 232, 214–236 (2006).
Maliva, R. G., Knoll, A. H. & Siever, R. Secular change in chert distribution: a reflection of evolving biological participation in the silica cycle. Palaios 4, 519–532 (1989).
Rost, B. & Riebesell, U. in Coccolithophores: From Molecular Processes to Global Impact (eds Thierstein, H. R. & Young, J. R. ) 99–125 (Springer, 2004).
Fowler, S. W. & Knauer, G. A. Role of large particles in the transport of elements and organic compounds through the oceanic water column. Prog. Oceanog. 16, 147–194 (1986).
Lavery, T. J. et al. Iron defecation by sperm whales stimulates carbon export in the Southern Ocean. Proc. R. Soc. Lond. B 277, 3527–3531 (2010).
Hutchinson, G. E. The biogeochemistry of vertebrate excretion. Bull. Am. Mus. Nat. Hist. 96, 1–554 (1950).
Carbone, C. & Narbonne, G. M. When life got smart: the evolution of behavioral complexity through the Ediacaran and early Cambrian of NW Canada. J. Paleontol. 88, 309–330 (2014).
Darwin, C. R. The Formation of Vegetable Mould, Through the Action of Worms, with Observations on Their Habits (John Murray, 1881).
Canfield, D. E. & Farquhar, J. Animal evolution, bioturbation, and the sulfate concentration of the oceans. Proc. Natl Acad. Sci. USA 106, 8123–8127 (2009).
Boyle, R. A. et al. Stabilization of the coupled oxygen and phosphorus cycles by the evolution of bioturbation. Nat. Geosci. 7, 671–676 (2014).
Logan, G. A., Hayes, J. M., Hieshima, G. B. & Summons, R. E. Terminal Proterozoic reorganization of biogeochemical cycles. Nature 376, 53–56 (1995).
Christian, H. J. et al. Global frequency and distribution of lightning as observed from space by the Optical Transient Detector. J. Geophys. Res. 108, 4005 (2003).
Wierzchowski, J., Heathcott, M. & Flannigan, M. D. Lightning and lightning fire, central cordillera, Canada. Int. J. Wildland Fire 11, 41–51 (2002).
Yair, Y. New results on planetary lightning. Adv. Space Res. 50, 293–310 (2012).
Belcher, C. M. & McElwain, J. C. Limits for combustion in low O2 redefine paleoatmospheric predictions for the Mesozoic. Science 321, 1197–1200 (2008).
Belcher, C. M., Yearsley, J. M., Hadden, R. M., McElwain, J. C. & Rein, G. Baseline intrinsic flammability of Earth's ecosystems estimated from paleoatmospheric oxygen over the past 350 million years. Proc. Natl Acad. Sci. USA 107, 22448–22453 (2010).
Glasspool, I. J., Edwards, D. & Axe, L. Charcoal in the Silurian as evidence for the earliest wildfire. Geology 32, 381–383 (2004).
Keeley, J. E., Pausas, J. G., Rundel, P. W., Bond, W. J. & Bradstock, R. A. Fire as an evolutionary pressure shaping plant traits. Trends Plant Sci. 16, 406–411 (2011).
Bond, W. J., Woodward, F. I. & Midgley, G. F. The global distribution of ecosystems in a world without fire. New Phytol. 165, 525–538 (2005).
Bond, W. J. & Scott, A. C. Fire and the spread of flowering plants in the Cretaceous. New Phytol. 188, 1137–1150 (2010).
Moreau, C. S., Bell, C. D., Vila, R., Archibald, S. B. & Pierce, N. E. Phylogeny of the ants: diversification in the age of angiosperms. Science 312, 101–104 (2006).
Cardinal, S. & Danforth, B. N. Bees diversified in the age of eudicots. Proc. R. Soc. Lond. B 280, 20122686 (2012).
Wilson, G. P. et al. Adaptive radiation of multituberculate mammals before the extinction of dinosaurs. Nature 483, 457–460 (2013).
Lenton, T. M. in Fire Phenomena and the Earth System: An Interdisciplinary Guide to Fire Science (ed. Belcher, C. M. ) 289–308 (John Wiley and Sons, 2013).
Wrangham, R. W., Jones, J. H., Laden, G., Pilbeam, D. & Conklin-Brittain, N. The raw and the stolen: cooking and the ecology of human origins. Curr. Anthropol. 40, 567–594 (1999).
Gowlett, J. A. J. & Wrangham, R. W. Earliest fire in Africa: towards the convergence of archaeological evidence and the cooking hypothesis. Azania 48, 5–30 (2013).
Carmody, R. N., Weintraub, G. S. & Wrangham, R. W. Energetic consequences of thermal and nonthermal food processing. Proc. Natl Acad. Sci. USA 108, 19199–19203 (2011).
Groopman, E. E., Carmody, R. N. & Wrangham, R. W. Cooking increases net energy gain from a lipid-rich food. Am. J. Phys. Anthropol. 156, 11–18 (2015).
Lotka, A. J. Elements of Physical Biology (Williams and Wilkins, 1925).
Smil, V. Enriching the Earth (MIT Press, 2001).
Erisman, J. W., Sutton, M. A., Galloway, J., Klimont, Z. & Winiwarter, W. How a century of ammonia synthesis changed the world. Nat. Geosci. 1, 636–639 (2008).
Pimm, S. L. et al. The biodiversity of species and their rates of extinction, distribution, and protection. Science 344, 1246752 (2014).
de Duve, C. Singularities (Cambridge Univ. Press, 2005).
Knoll, A. H. & Bambach, R. K. Directionality in the history of life: diffusion from the left wall or repeated scaling of the right? Paleobiology 26, 1–14 (2000).
Maynard Smith, J. & Szathmáry, E. The Major Transitions in Evolution (WH Freeman, 1995).
Szathmáry, E. Toward major evolutionary transitions theory 2.0. Proc. Natl Acad. Sci. USA 112, 10104–10111 (2015).
Calcott, B. & Sterelny, K. (eds) The Major Transitions in Evolution Revisited (MIT Press, 2011).
Morris, J. J., Kirkegaard, R., Szul, M. J., Johnson, Z. I. & Zinser, E. R. Facilitation of robust growth of Prochlorococcus colonies and dilute liquid cultures by ‘helper’ heterotrophic bacteria. Appl. Environ. Microbiol. 74, 4530–4534 (2008).
Abed, R. M. M. Interaction between cyanobacteria and aerobic heterotrophic bacteria in the degradation of hydrocarbons. Int. Biodeter. Biodegr. 64, 58–64 (2010).
Shen, H., Niu, Y., Xie, P., Tao, M. & Yang, X. Morphological and physiological changes in Microcystis aeruginosa as a result of interactions with heterotrophic bacteria. Freshwater Biol. 56, 1065–1080 (2011).
Benton, M. J. & Twitchett, R. J. How to kill (almost) all life: the end-Permian extinction event. Trends Ecol. Evol. 18, 358–365 (2003).
Song, H. et al. Anoxia/high temperature double whammy during the Permian–Triassic marine crisis and its aftermath. Sci. Rep. 4, 4132 (2014).
Kasprak, A. H. et al. Episodic photic zone euxinia in the northeastern Panthalassic Ocean during the end-Triassic extinction. Geology 43, 307–310 (2015).
Kring, D. A. The Chicxulub impact event and its environmental consequences at the Cretaceous–Tertiary boundary. Palaeogeogr. Palaeoclimatol. Palaeoecol. 255, 4–21 (2007).
Vermeij, G. J. Inequality and the directionality of history. Am. Nat. 153, 243–253 (1999).
Smil, V. Energy in Nature and Society: General Energetics of Complex Systems (MIT Press, 2008).
Stevenson, D. J. Life-sustaining planets in interstellar space? Nature 400, 32 (1999).
Strigari, L. E., Barnabè, M., Marshall, P. J. & Blandford, R. D. Nomads of the Galaxy. Mon. Not. R. Astron. Soc. 423, 1856–1865 (2012).
Nealson, K. H., Inagaki, F. & Takai, K. Hydrogen-driven subsurface lithoautotrophic microbial ecosystems (SLiMEs): do they exist and why should we care? Trends Microbiol. 13, 405–410 (2005).
Watson, A. J. Implications of an anthropic model of evolution for emergence of complex life and intelligence. Astrobiology 8, 175–185 (2008).
Miller, B. G. & Wolfenden, R. Catalytic proficiency: the unusual case of OMP decarboxylase. Annu. Rev. Biochem. 71, 847–885 (2002).
Kim, J., Dong, H., Seabaugh, J., Newell, S. W. & Eberl, D. D. Role of microbes in the smectite-to-illite reaction. Science 303, 830–832 (2004).
Farquhar, G. D., Ehleringer, J. R. & Hubick, K. T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 40, 503–537 (1989).
Schidlowski, M. Carbon isotopes as biogeochemical recorders of life over 3.8 Ga of Earth history: evolution of a concept. Precambrian Res. 106, 117–134 (2001).
Allen, J. F. The function of genomes in bioenergetic organelles. Phil. Trans. R. Soc. Lond. B 358, 19–38 (2003).
Fritz-Laylin, L. K. et al. The genome of Naegleria gruberi illuminates early eukaryotic versatility. Cell 140, 631–642 (2010).
Akashi, H. & Gojobori, T. Metabolic efficiency and amino acid composition in the proteomes of Escherichia coli and Bacillus subtilis. Proc. Natl Acad. Sci. USA 99, 3695–3700 (2002).
Seligmann, H. Cost-minimization of amino acid usage. J. Mol. Evol. 56, 151–161 (2003).
Swire, J. Selection on synthesis cost affects interprotein amino acid usage in all three domains of life. J. Mol. Evol. 64, 558–571 (2007).
Craig, C. L. & Weber, R. S. Selection costs of amino acid substitutions in ColE1 and ColIa gene clusters harbored by Escherichia coli. Mol. Biol. Evol. 15, 774–776 (1998).
Mills, D. B. & Canfield, D. E. Oxygen and animal evolution: did a rise of atmospheric oxygen trigger the origin of animals? BioEssays 36, 1145–1155 (2014).
Mentel, M., Röttger, M., Leys, S., Tielens, A. G. M. & Martin, W. F. Of early animals, anaerobic mitochondria, and a modern sponge. BioEssays 36, 924–932 (2014).
Sperling, E. A. et al. Oxygen, ecology, and the Cambrian radiation of animals. Proc. Natl Acad. Sci. USA 110, 13446–13451 (2013).
Nursall, J. R. Oxygen as a prerequisite to the origin of the Metazoa. Nature 183, 1170–1172 (1959).
Knoll, A. H. & Carroll, S. B. Early animal evolution: emerging views from comparative biology and geology. Science 284, 2129–2137 (1999).
Chen, X. et al. Rise to modern levels of ocean oxygenation coincided with the Cambrian radiation of animals. Nat. Commun. 6, 7142 (2015).
Reinhard, C. T., Planavsky, N. J., Olson, S. L., Lyons, T. W. & Erwin, D. H. Earth's oxygen cycle and the evolution of animal life. Proc. Natl Acad. Sci. USA 113, 8933–8938 (2016).
Lenton, T. M., Boyle, R. A., Poulton, S. W., Shields-Zhou, G. A. & Butterfield, N. J. Co-evolution of eukaryotes and ocean oxygenation in the Neoproterozoic era. Nat. Geosci. 7, 257–265 (2014).
Acknowledgements
Many thanks to G. Carr, T. Carvalho, D. C. Catling, D. Haydon, T. Goldberg, P. Jarne, A. H. Knoll, E. Kroll, N. Judson, N. Lane, T. Lenormand, G. Lichfield, B. C. T. Mason, O. Morton, J. Rolff, J. Swire, and especially A. Courtiol for helpful discussions and for comments on an earlier draft of the manuscript. Many thanks to W. F. Martin and T. M. Lenton for insightful reviews that improved the manuscript. Figure 1 was drawn by graphic designer F. Zsolnai, many thanks.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The author declares no competing financial interests.
Rights and permissions
About this article

Cite this article
Judson, O. The energy expansions of evolution. Nat Ecol Evol 1, 0138 (2017). https://doi.org/10.1038/s41559-017-0138
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41559-017-0138
This article is cited by
-
The oxygen bottleneck for technospheres
Nature Astronomy (2023)
-
Linking extreme seasonality and gene expression in Arctic marine protists
Scientific Reports (2023)
-
Exploring the structural aspects and therapeutic perspectives of cyanobacterial phycobiliproteins
3 Biotech (2022)
-
Harmonics, evolutionary generators, DANCE, and HEAR—functional dimensions
Environmental Science and Pollution Research (2021)
-
Equal fitness paradigm explained by a trade-off between generation time and energy production rate
Nature Ecology & Evolution (2018)
