
Abstract
The efficient exploration of biologically relevant chemical space is essential for the discovery of bioactive compounds. A molecular design principle that possesses both biological relevance and structural diversity may more efficiently lead to compound collections that are enriched in diverse bioactivities. Here the diverse pseudo-natural product (PNP) strategy, which combines the biological relevance of the PNP concept with synthetic diversification strategies from diversity-oriented synthesis, is reported. A diverse PNP collection was synthesized from a common divergent intermediate through developed indole dearomatization methodologies to afford three-dimensional molecular frameworks that could be further diversified via intramolecular coupling and/or carbon monoxide insertion. In total, 154 PNPs were synthesized representing eight different classes. Cheminformatic analyses showed that the PNPs are structurally diverse between classes. Biological investigations revealed the extent of diverse bioactivity enrichment of the collection in which four inhibitors of Hedgehog signalling, DNA synthesis, de novo pyrimidine biosynthesis and tubulin polymerization were identified from four different PNP classes.

Similar content being viewed by others
Main
The strategic navigation of biologically relevant chemical space for the discovery of bioactive small molecules is a core principle of chemical biology and drug discovery programmes1,2. Through evolution, nature has explored biologically relevant chemical space to afford current natural product (NP) structures that represent biologically prevalidated chemical matter. NPs have been and continue to be a rich source of chemical probes and therapeutics3; however, the development of NPs into chemical probes and/or therapeutics can be restricted by low availability and insufficient access to derivatives.
Several molecular design principles based on NP structures, such as function-oriented synthesis4,5,6, biology-oriented synthesis7,8, dynamic retrosynthetic analysis9,10 and others11, have successfully addressed these issues by providing compounds that retain the bioactivities of the NPs of interest but are more synthetically tractable than the guiding NPs. Nevertheless, these methods are inherently limited since the resulting compounds have core scaffolds that are present in the guiding NPs and therefore are likely to have the same bioactivities as the guiding NPs. NPs, as well as design principles that are derivatives of NP scaffolds, are also limited due to evolutionary constraints. Natural evolution is a very slow process that has resulted in NPs occupying only a fraction of theoretical NP-like chemical space12. Therefore, relying solely on current NP scaffolds, which are synthesized via existing biosynthetic pathways, has limitations in molecular discovery.
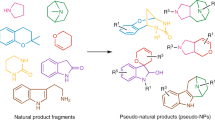
Employing methods that are inspired by nature and go beyond current biosynthetic pathways may facilitate the exploration of biologically relevant NP-like chemical space13,14. The complexity-to-diversity approach is a chemical extension of natural biosynthetic pathways in which suitable NPs or synthetic NP-like compounds are subjected to ring distortion reactions to afford compound collections with diverse scaffolds that are distinct from the starting NP yet retain the biological relevance and complexity of NPs15,16,17. We have recently introduced a design principle for pseudo-natural products (PNPs) through the de novo combination of NP fragments (Fig. 1a)18,19,20,21,22,23,24. The PNP concept identifies fragments characteristic in different NP classes and combines them in arrangements that are novel and not accessible by known biosynthetic pathways. The PNP scaffolds are not known to be produced in nature but may retain the biological relevance of NPs and therefore have diverse bioactivity profiles and targets. Different NP fragments can be combined in different arrangements and connectivity patterns to access various PNP scaffolds that have diverse bioactivity profiles25,26,27. However, access to diverse PNPs can be laborious in which typically specific starting materials must be made for each scaffold.

a, PNP logic: the combination of NP fragments in unprecedented arrangements. b, DOS logic: scaffold diversity using a build–couple–pair strategy from available starting materials. c, Merging the two concepts of PNPs and DOS, generation of dPNPs. d, An overview of dPNP compound class syntheses from common indole-based starting materials.
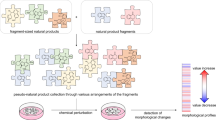
To access diverse scaffolds more rapidly, we envisioned developing a design principle for PNP compound collections based on a common divergent intermediate, whereby one common intermediate could give rise to a variety of PNP scaffolds that may possess various bioactivities. In this context, the diversity-oriented synthesis (DOS) strategy28,29,30,31,32 can cover a wide area of chemical space to generate compound collections with diverse and complex scaffolds, with a build–couple–pair approach as one example31 (Fig. 1b). DOS does lead to diverse compound collections that may have NP-like features that positively contribute to biological performance33, such as high fraction of sp3-hybridized centres and stereogenic content. However, in our perspective, DOS, unlike the PNP design principle, is not directly based on NP structures, and therefore scaffolds resulting from DOS do not necessarily include fragments of biological relevance18. Thus, we envisioned combining the two logics of DOS and PNPs to generate a compound collection that incorporates both scaffold diversity and biological relevance, termed as diverse PNPs (dPNPs, Fig. 1c).
In this Article, we report a strategy to access dPNP scaffolds from a common divergent intermediate. Chemical methodologies for indole dearomatization34,35,36,37,38 were developed to streamline scaffold construction including intramolecular indole dearomatizations via a σ-acylpalladium intermediate through in situ-generated carbon monoxide (CO). Dearomatization is a powerful approach to access underexplored three-dimensional topologies39 from flat aromatic compounds. Furthermore, dearomatization can introduce new stereogenic centre(s) and increase the fraction of sp3 carbons, both of which are favourable properties for target-binding selectivity and clinical progression of drug candidates33,40. The dearomatization of indoles and coupling and/or pairing of intermediates resulted in a total of 154 PNPs constituting eight different classes with diverse combinations and orientations of NP fragments (Fig. 1d). The PNP collection was assessed by cheminformatic methods and was found to be chemically diverse between compound classes. Phenotypic screening and morphological profiling revealed diverse bioactivity profiles within the collection. In particular, unprecedented chemotypes for Hedgehog (Hh) signalling inhibition, DNA synthesis inhibition, de novo pyrimidine biosynthesis inhibition and tubulin modulation were identified.
Results
PNP collection synthesis via indole dearomatizations
The initial design was the construction of a scaffold containing indolenine and indanone fragments (Fig. 2a). Indolenine and indanone fragments are commonly found in various bioactive NPs; however, known biosynthetic pathways do not produce NP scaffolds that have both indolenine and indanone fragments (Supplementary Fig. 1). We envisioned that the combination of these fragments would be possible by dearomatizing planar indole starting materials with a C3-tethered electrophile via a palladium-catalysed intramolecular carbonylation/indole dearomatization cascade. The dearomatative nature of the reaction and resulting spirocyclic fusion pattern should result in compounds with higher degrees of chirality and three-dimensionality41,42.

a, Indolenine and indanone fragment combination to construct spiroindolylindanone PNPs. b, Method development and optimization of a CO insertion reaction to construct spiroindolylindanone PNPs using N-formyl saccharin (2a) as a non-toxic bench-stable solid and inexpensive CO surrogate. c, Substrate scope for the spiroindolylindanone synthesis. aThe PNPs in this manuscript are identified by the letter of the compound class followed by the compound number within the compound class, that is, A1 is class A, compound 1. bIsolated yields are shown.
Initial attempts started with 1a (R1 = Me, R2–6 = H) as a model substrate (Fig. 2b) and CO gas as a carbonyl source with the goal of obtaining dearomatized compound A1 (PNPs are identified by the letter of the compound class followed by the compound number within the compound class (A1 is class A, compound 1)) (refs. 43,44,45,46,47). Treatment of 1a with palladium acetate as a catalyst and 1,1′-bis(diphenylphosphino)ferrocene as a ligand in the presence of triethylamine in benzene at 1 atm. CO gas at 80 °C delivered the desired product A1 in 23% yield. Optimization of different reaction parameters such as solvent, ligand and base in the presence of CO gas did not lead to an improved yield (Supplementary Tables 1–3). We hypothesized that superstoichiometric amounts of CO gas in the medium may be hindering the reaction and focused on CO surrogates that can generate CO gas in a controlled manner in situ48,49,50. Gratifyingly, the use of N-formyl saccharin (2a) in the presence of palladium acetate, Xantphos (4,5-bis(diphenylphosphino)-9,9-dimethylxanthene) and sodium carbonate in N,N-dimethyl formamide (DMF) provided A1 in an excellent yield of 86%. N-formyl saccharin (2a) was developed by Manabe et al. and is an inexpensive, safe, efficient and environmentally friendly CO surrogate49,50. Employing other CO surrogates in place of 2a, such as 1,1′-carbonyldiimidazole, 2,4,6-trichlorophenyl formate, molybdenum hexacarbonyl and dicobaltoctacarbonyl, provided little or no desired product (Supplementary Table 4).
The scope of the developed dearomatization reaction was explored and led to the construction of PNP class A termed spiroindolylindanones (Fig. 2c). Substrates having either electron-withdrawing or electron-rich groups afforded the desired products in moderate to high yields (A2, A4, A6, A7, A22 and A25–A27 and A3, A5, A20 and A21, respectively). Multi-substituted and heterocycle-based aryl bromides also reacted to afford the dearomatized products in good yields (A8, A10 and A29–A31 and A11, A12 and A32–A35, respectively). C5-substituted indoles reacted well regardless of the substituents and substitution pattern of the tethered aryl bromide to furnish spiroindolylindanones A14–A35 in good to excellent yields. C2-phenyl-substituted indoles reacted to afforded the dearomatized products A36–A41 in relatively lower yields than their methyl-substituted derivatives, possibly due to steric and/or electronic reasons. Overall, the developed methodology is an unprecedented example of a carbonylation/intramolecular indole dearomatization via a σ-acylpalladium intermediate cascade. The reaction is operationally simple and robust, which allowed for the rapid synthesis of class A.
Derivatization of compounds belonging to class A was possible through various reaction pathways and sequences (Fig. 3a). Reduction of the indolenine moiety was performed with Hantzsch ester and a catalytic amount of pyridinium p-toluenesulfonate (PPTS) to generate the spiro-indoline–indanone class B with good to excellent diastereoselectivities (6:1 ≥ 20:1 diastereomeric ratio (d.r.))51. Single-crystal analysis of the major diastereomer of compound B10 revealed a cis-relationship between the methyl and carbonyl groups.

a, Access to classes B–H from class A. The relative configuration of B10 was determined by X-ray crystallography (Supplementary Information), and the structures of other products (including class C) were drawn by analogy. Diastereomeric ratios of class B were determined by 1H nuclear magnetic resonance. For individual diastereomeric ratios, see Supplementary Information. b, Synthesis of classes F, G and H from 1 and 5, respectively, utilizing the NH of indole derivatives. Isolated yields are shown. Parent NP examples and the corresponding NP fragments from which the PNPs are derived are shown on the right side of the figures. RT, room temperature; CFL, compact fluorescent lamp; fan, 3 W cooling fan.
Functional group installation at the free amine group of class B generated a new class of compounds (class C). Treatment of 2-methyl indolenines with α-halo-acetyl chloride derivatives furnished exocyclic-olefinic α-halo-amides (class D). The isoquinolinone fragment, a common motif in a variety of biologically important alkaloids, was fused to the spiroindolylindanone scaffold (class A) by employing methyl 2-bromobenzoate in the presence of a palladium/Xantphos catalyst and K3PO4 as the base51. The final indoline–indanone–isoquinolinone scaffold defines a PNP class with both spiro- and edge-fusion combinations (class E).
To establish further scaffold diversity, we envisioned rearranging 1 and subjecting it to different variations of the developed Pd-catalysed reaction conditions. Inspired by previous reports on photocatalytic oxidative semipinacolic rearrangement52,53, we developed a seminal reaction by employing 2 mol% Eosin Y as a photocatalyst and SiO2 as solid additive to induce the rearrangement of 1 to indolin-3-one derivatives 4. By employing the developed Pd-catalysed conditions, indolin-3-one derivatives (4) underwent direct C–N cross couplings in the absence of the CO surrogate 2a to yield 3-oxindole–indoline-fused PNPs (class F, Fig. 3b). In the presence of 2a, a carbonylation/cyclization cascade resulted in a fusion-edge combination of 3-oxindole and dihydroisoquinolone fragments (class G, Fig. 3b). Finally, compounds 5, stemming from oxa-Pictet-Spengler reactions of tryptophol and benzaldehyde derivatives (Supplementary Fig. 2), were treated with N-formyl saccharin for CO insertion. In this case, dearomatization of the indole and formation of the isoquinolinone fragment yielded indoline–isoquinolinone–tetrahydropyran PNPs (class H, Fig. 3b).
In total, 154 PNPs representing eight subclasses with different NP fragment combinations and fusion patterns were synthesized from divergent intermediates 1 or compounds 5. Searches in the Dictionary of Natural Products revealed that neither the NP fragment combinations nor the scaffolds of the subclasses are observed in known NPs (Supplementary Fig. 1). By employing the combination of diverse PNP design with developing and implementing synthetic chemical methodologies, we were able to readily access a diverse range of PNPs that are not possible through existing biosynthetic pathways.
Cheminformatic analysis of the PNP classes
For the characterization of structural and physiochemical properties of the PNP classes, characteristic properties were computed using the open-source software RDKit54. The PNPs were compared with reference sets including NPs in the ChEMBL29 database (representing bioactive NPs, 45,679 compounds)55, DrugBank compounds (representing marketed and investigational drugs, 4,866 compounds)56 and the Enamine Advanced Screening Collection (representing a synthetic drug-like screening library, 527,411 compounds)57. To analyse the molecular shapes of the PNP collection and reference sets, principal moments of inertia (PMI) were calculated58. The PMI calculation revealed that the PNP collection has shape diversity and overlaps with concentrated areas of bioactive compounds (Fig. 4a). Atom connectivities were evaluated by a NP-likeness score59 calculation and were found to notably overlap with DrugBank compounds (Fig. 4b). A lack of considerable overlap with ChEMBL NPs may be due to the PNPs representing fragment combinations and arrangements that are not found in nature. A quantitative estimation of drug-likeness60 (QED) analysis suggested that the PNP collection has favourable drug-like properties, which are similar to the properties calculated for the Enamine Advanced Screening Collection (Fig. 4c). A principal component analysis61 of 17 molecular descriptors shows that the PNP collection occupies chemical space that is shared by high-density regions of all three reference sets (Supplementary Figs. 4–6). Interestingly, 77% of the PNPs have lead-like sizes, that is, 14 ≤ heavy atoms ≤ 26 (refs. 62,63) and, in combination with drug-like properties suggested by QED analysis, indicates that the PNP collection may be lead like. Future compound classes could be designed by combining NP fragments with appropriate molecular properties to afford other lead-like PNP collections.

a, A PMI plot for the shape of the PNPs (black circles). The corners of the triangle within the plot indicate a rod-like shape (top left), disk-like shape (bottom middle) and sphere-like shape (top right). The contour lines represent a Gaussian kernel density estimation with ten steps. For a PMI plot with individual PNP subclasses, see Supplementary Fig. 3. b, NP-likeness scores of the PNPs (black curve) compared with the DrugBank compound collection (orange curve), ChEMBL NPs (green curve) and Enamine building blocks (blue curve). c, QED of the PNPs (black curve) compared with the DrugBank compound collection (orange curve), ChEMBL NPs (green curve) and Enamine building blocks (blue curve). d, Box plot of intra- and interclass Tanimoto similarity calculations of Morgan fingerprints (ECFC4) following Tukey’s definitions with outliers83. Centre line, median; box limits, upper and lower quartiles; whiskers, 1.5× interquartile range; and points, outliers. The dashed line indicates the 95th percentile median (0.23) of random reference compound subsets. For full cross-similarity values, see Supplementary Figs. 9–10. e, Box plot of intra- and interclass Tanimoto similarity calculations of Morgan fingerprints (ECFP6) following Tukey’s definitions with outliers83. Centre line, median; box limits, upper and lower quartiles; whiskers, 1.5× interquartile range; and points, outliers. The dashed line indicates the 95th percentile median (0.17) of random reference compound subsets. For full cross-similarity values, see Supplementary Figs. 11 and 12. The number of compounds in reference sets is 527,411 (50,000 random compounds were selected for PMI analysis) for Enamine, 4,866 for DrugBank and 45,679 for ChEMBL NPs.
Intra- and interclass Tanimoto similarities of the Morgan fingerprints of two different designs (ECFC4 and ECFP6) were calculated to determine structural diversity (Fig. 4d,e, and for full comparisons, see Supplementary Figs. 9–12). Within compound classes, the median similarities were high (ECFC4 of 0.49–0.77 and ECFP6 of 0.32–0.67), whereas between compound classes the median similarities are distinctly lower (ECFC4 of 0.26–0.44 and ECFP6 of 0.10–0.29). For comparison, a threshold of randomness was determined by calculating the 95th percentile median similarities from 100 subsets with 100 random members from the Enamine Advanced Screening Collection (ECFC4 of 0.23 and ECFP6 of 0.17; for more details, see Supplementary Information). Several interclass PNP similarities are near but do not fall below the randomness thresholds. This may be expected as similar NP fragments are conserved throughout the compound collection, albeit in different arrangements. It can therefore be concluded that the majority of interclass structures of the PNP collection are diverse.
Identification of an inhibitor of Hh signalling
The bioactivity of the PNP collection was investigated by employing cell-based assays monitoring different cellular pathways and signalling cascades including autophagy, kynurenine production and Hh-dependent osteoblast differentiation. Gratifyingly, spiroindolylindanone class A was enriched with inhibitors of Hh-dependent osteoblast differentiation. No compounds from the PNP collection were identified as hits, that is, >50% inhibition at 10 µM, in assays monitoring autophagy or kynurenine production.
The Hh signalling pathway plays an important role during vertebrate embryonic and post-embryonic development including tissue homeostasis and regeneration64. Hh signalling is activated through the binding of Hh ligands to the membrane receptor Patched1, which in turn relieves Patched1-mediated inhibition of the membrane protein Smoothened (SMO). This activation ultimately leads to the transcription of Hh target genes, such as Gli1 and Ptch1, by the transcription factors glioma-associated oncogene homologues 2 and 3 (GLI2 and GLI3) (ref. 65). Unregulated Hh signalling has been connected to cancers such as basal cell carcinoma and medulloblastoma. Therefore, novel therapeutics for inhibiting the Hh pathway are in high demand66.
To identify novel inhibitors of Hh signalling, we employed an osteoblast differentiation assay using C3H/10T1/2 mesenchymal progenitor cells upon stimulation with purmophamine67. Stimulation of the Hh pathway induces osteoblast differentiation, which can be detected through monitoring of alkaline phosphatase expression and activity. The screen revealed compound A23 is an inhibitor of Hh-dependent cell differentiation with a half-maximal inhibitory concentration of 0.93 ± 0.13 μM and no effect on cell viability (Fig. 5a).

a, Selected compounds with structural variation to the most active compound (A23) in an osteoblast differentiation assay. Cell viability was assessed using a CellTiter-Glo Luminescent Cell Viability Assay and treating C3H10T1/2 cells with compound (30 µM) in the absence of purmorphamine for 96 h. The viability of cells treated with DMSO was set to 100%. IC50, half-maximal inhibitory concentration. b, C3H10T1/2 cells were treated for 96 h with 1.5 µM purmorphamine, DMSO as a control or compound A23. The activity of alkaline phosphatase was measured to determine Hh pathway activity. Values for cells treated with purmorphamine and DMSO were set to 100%. The data are the mean values ± s.d. of three biological replicates (n = 3). c, Alpl gene expression; C3H10T1/2 cells were incubated for 96 h with 1.5 µM purmorphamine and DMSO, 1 µM vismodegib (vismo) or 1, 5 or 10 µM of A23 before RT–qPCR. Data are mean values ± s.d. of three biological replicates (n = 3). The P values relative to cells treated with DMSO and purmorphamine are <0.0001 for DMSO-treated, <0.0001 for vismo (1 µM)-treated, 0.0014 for A23 (1 µM)-treated, 0.0005 for A23 (5 µM)-treated and 0.0006 for A23 (10 µM)-treated cells. d,e, Expression of the Hh target gene Gli1 (d) and Ptch1 (e). C3H10T1/2 cells were incubated with 1.5 µM of purmorphamine and DMSO, 1 µM vismo or A23 (1 µM, 5 µM or 10 µM) for 96 h before RT–qPCR. Data are mean values ± s.d. of three biological replicates (n = 3). For Hh target gene Gli1, the P values relative to cells treated with DMSO and purmorphamine are 0.0001 for DMSO-treated, 0.0003 for vismo (1 µM)-treated, 0.0477 for A23 (1 µM)-treated, 0.0052 for A23 (5 µM)-treated and 0.0203 for A23 (10 µM)-treated cells. For Hh target gene Ptch1, the P values relative to cells treated with DMSO and purmorphamine are 0.0010 for DMSO-treated, 0.0024 for vismo (1 µM)-treated, 0.4559 for A23 (1 µM)-treated, 0.0111 for A23 (5 µM)-treated and 0.0142 for A23 (10 µM)-treated cells. f, SMO binding assay. HEK293T cells were transfected with a SMO-expressing plasmid. After 48 h, the cells were fixed and incubated with BODIPY–cyclopamine (green, 5 nM) and treated with either DMSO, vismo or A23 for 4 h. The nuclei were visualized by staining the cells with 4,6-diamidino-2-phenylindole (DAPI) (blue). The images are representative of three biological replicates (n = 3). Scale bar, 30 µm. For c–e, statistical analyses were performed relative to DMSO/purmorphamine by employing unpaired two-tailed t-tests with Welch’s correction (*P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001; NS, not significant).
Variation of the substitution pattern on A23 to establish a structure–activity correlation did not uncover more potent compounds (Fig. 5a). Removal of chlorine and fluorine substituents led to a loss in potency (A1) and affected cell viability. Reintroduction of a chlorine (A16) to the indolenine moiety at R6 led to a more potent activity relative to A1 without affecting cell viability, whereas a methyl (A14) or a fluorine (A15) at the same position led to similar and weaker activities relative to A1, respectively. Addition of a fluorine to the indanone moiety at R4 (A6) led to a loss in potency relative to A1 as did the introduction of a methyl (A3) or dimethoxy substituents (A9). Moving the fluorine substituent to R5 (A7) led to better activity. When compounds with a chlorine at R6 and similar substitution patterns on the indanone moiety as above were evaluated, only the activity of A23 increased, whereas A20, A30 and A27 became less active. Compounds A6, A16, A23 and two other derivatives (A8 and A21) were resynthesized and re-evaluation in an osteoblast differentiation assay resulted in similar activities to the original batches (Supplementary Fig. 13). Compound A23 was the most active inhibitor (Fig. 5b) and was also found to suppress alkaline phosphatase gene expression (Alpl) in a dose-dependent manner (Fig. 5c), which is in line with inhibition of osteoblast differentiation.
Evaluation of Hh genes by quantitative reverse transcription PCR (RT–qPCR)68 showed that A23 reduced expression of the Hh target genes Ptch1 and Gli1 (reduction of 46% and 47% at 10 µM, respectively, Fig. 5d,e). As SMO is frequently targeted by small molecules, the binding of A23 to SMO was explored in a competition assay using fluorinated boron-dipyrromethene (BODIPY)–cyclopamine69. Cyclopamine is a NP that binds to the heptahelical bundle in SMO70 that is targeted by the most SMO modulators. Compound A23 did not compete with BODIPY–cyclopamine and, therefore, most likely does not bind to SMO (Fig. 5f). All together, these results indicate that A23 is inhibiting Hh signalling independent of direct SMO modulation. It should be noted that current therapies affecting Hh signalling target SMO, which can cause mutations leading to drug resistance71. Therefore, novel inhibitors of the Hh pathway that do not target SMO are of particular interest.
PNP biological evaluation via the CPA
Beyond monitoring specific biological processes or signalling pathways, the PNP library was evaluated by morphological profiling via the cell painting assay (CPA)72,73. This unbiased morphological profiling method monitors phenotypic changes in cells upon compound treatment by employing six fluorescent dyes that selectively stain various compartments. High-content imaging via multi-channel fluorescence microscopy and automated image analysis extracts and quantifies the changes of cellular morphologies into 579 cellular features to generate a characteristic profile.
An induction value, representing the percentage of significantly changed features relative to dimethylsulfoxide (DMSO) controls, is a measure for compound activity in the CPA74. Investigation of the PNP collection at concentrations ranging from 1 to 50 µM revealed that 78% of the compounds are characterized by an induction value ≥5% and are considered bioactive (Supplementary Fig. 14). Furthermore, all compound classes contained CPA-active compounds. For comparison, 42% of compounds with annotated bioactivities (2,230 of 5,307) and 45% of our in-house, non-annotated compounds (6,581 of 14,784) have induction values ≥5% at concentrations up to 50 µM.
The CPA can enable target or mode-of-action (MoA) identification by directly comparing full CPA profiles of uncharacterized compounds with reference compounds with annotated bioactivities75. We have recently introduced the concept of a CPA subprofile analysis to facilitate rapid target or MoA prediction76. In short, subprofiles are defined by reducing the number of profile features to only those that are shared by annotated compounds in a particular bioactivity cluster. High similarities (>85%) to bioactivity cluster profiles can facilitate the annotation of uncharacterized compounds without prior knowledge of the top biosimilar reference compounds. So far, 12 bioactivity cluster subprofiles have been defined that include biological targets related to AKT/PI3K/MTOR, Aurora kinases, bromodomain and extra-terminal (BET) domain, de novo pyrimidine biosynthesis, DNA synthesis, histone deacetylase (HDAC), HSP90, Na+/K+ ATPases and tubulin, or processes such as lysosomotropism/cholesterol homeostasis regulation, protein synthesis and uncoupling of the mitochondrial proton gradient.
All CPA PNPs profiles with induction values >15% and relative cell counts >50% were subjected to a subprofile analysis. Overall, several PNPs from different compound classes induced subprofiles that were similar to reference subprofiles including inhibition of DNA synthesis, inhibition of de novo pyrimidine biosynthesis and modulation of tubulin (Fig. 6a and Supplementary Figs. 15 and 16).

a, A heat map showing the percentage of PNPs in each class that have >85% similarity to a bioactivity cluster profile and induction values >15%. The PNPs were measured at various concentrations (≤50 µM), generating several profiles for each PNP. For each PNP, the profile with the highest similarity to a bioactivity cluster was selected for this analysis. L/CH, lysosomotropism/cholesterol homeostasis. For heat maps of bioactivity cluster similarities of individual profiles of hit compounds, see Supplementary Figs. 15 and 16. b, The chemical structure of C3, C3-resynthesized, C18 and E13. c, The influence of C3, C3-resynthesized and C18 on DNA content and the cell cycle. U2OS cells were treated with DMSO or compound (30 µM) for 22 h followed by the addition of 10 µM EdU and incubated for an additional 2 h. DNA-incorporated EdU was labelled with Alexa Fluor 488 via click reaction and DNA was stained with propidium iodide. Single-cell analysis via flow cytometry measuring EdU incorporation and total DNA content was used to determine the percentage of cells in either G1 (2N), S (2N–4N) or G2 (4N) phase. Data are mean values ± s.d. (n = 10,000 cells examined over three biologically independent samples). Histograms of the FACS analysis can be found in Supplementary Fig. 19. d, Uridine rescue assay. HCT116 cells were treated with either DMSO (control), E13 or E13 in the presence of uridine (100 µM). Cell confluence was used as a measure of cell proliferation and was monitored over a 96 h period using an IncuCyte ZOOM/S3. Data are mean values ± s.d. of four independent replicates (n = 4).
Compound C3 (Fig. 6b) was identified by the CPA subprofile analysis as a potential inhibitor of DNA synthesis and was the only compound in class C that induced a profile with >85% similarity to the DNA synthesis cluster profile with all other compounds in the class, having <60% similarity (Supplementary Fig. 17). A brief structure–phenotype relationship74 study of C3 was conducted with respect to the DNA synthesis cluster. The indolenine and N-unsubstituted indoline synthetic precursors to C3 (A1 and B1, respectively) both have weak activity in the CPA and low similarity to the DNA synthesis profile (Supplementary Fig. 18). Introduction of an acrylamide moiety to B1 affords C3, which has a high similarity to the DNA synthesis profile (92% similarity). Other N-functionalized derivatives of B1 including α-halo amide compounds (C1 and C2), an ethyl carbamate compound (C4), an N-allylated compound (C5) and an N-tosylated compound all have very low similarities to the DNA synthesis profile. These results suggest that the acrylamide moiety of C3 is crucial for producing a highly similar phenotype to the DNA synthesis profile.
The influence of C3 on DNA synthesis and the cell cycle was investigated by fluorescence-activated cell sorting (FACS) flow cytometry employing 5-ethynyl-2’-deoxyuridine (EdU) and propidium iodide. EdU is an alkynyl thymidine analog that can be incorporated into newly synthesized DNA, and the attachment of a fluorophore via click chemistry and subsequent microscopy can be used to quantify DNA synthesis77. Additional staining with propidium iodide, which quantifies overall DNA content, and single-cell analysis via FACS flow cytometry can determine the influence of compounds on the cell cycle. At 30 µM, compound C3 led to a remarkable decrease in DNA-incorporated EdU (Supplementary Figs. 19 and 20) and a depletion of cells in the S phase relative to DMSO controls (Fig. 6c). These results indicate that C3 prevents DNA synthesis and confirms the CPA-generated MoA hypothesis. Compound C3 was resynthesized (C3-resynthesized) and reverified to be an inhibitor of DNA synthesis (Fig. 6c and Supplementary Figs. 19 and 20). Since C3 has an acrylamide moiety that may act as a Michael acceptor, a saturated ethyl amide derivative (C18, Fig. 6b) was synthesized and evaluated. Interestingly, C18 provided similar results to a DMSO control via FACS flow cytometry (Fig. 6c and Supplementary Figs. 19 and 20) and confirms that the acrylamide moiety of C3 is necessary for its activity as a DNA synthesis inhibitor.
The CPA subprofile analysis (Fig. 6a) also suggested that E13 (Fig. 6b) may affect a target related to de novo pyrimidine biosynthesis (85% biosimilarity to the de novo pyrimidine biosynthesis profile and 28% induction at 50 µM) (ref. 78). Pyrimidine biosynthesis is one facet of the biosynthesis of nucleic acids and nucleotide cofactors that are required for DNA and RNA synthesis as well as several other essential cellular processes79. The dysregulation of nucleotide metabolism is prevalent in various types of cancers as well as viral infections, and small molecules that target nucleotide biosynthetic pathways have been successful therapeutics to combat these diseases79.
To examine whether E13 affects de novo pyrimidine biosynthesis, a uridine rescue assay was employed (Fig. 6d). If pyrimidine biosynthesis is inhibited, pyrimidine nucleotides will be depleted, leading to the suppression of cellular growth due to the lack of DNA synthesis and transcription. The supplementation of excess uridine can restore normal cell growth by bypassing the need for de novo pyrimidine biosynthesis. When treated with E13 at 30 µM, the growth of HCT116 cells was impaired over a 96 h time frame (Fig. 6d). Co-treatment of cells with E13 and supplemental uridine (100 µM) completely restored cell growth relative to the DMSO control, confirming the CPA-guided hypothesis that E13 is inhibiting de novo pyrimidine biosynthesis. Compound E13 was resynthesized and its bioactivity as an inhibitor of de novo pyrimidine biosynthesis was reconfirmed (Supplementary Fig. 21a).
Besides E13, no other compounds in class E had similarities to the de novo pyrimidine biosynthesis bioactivity profile >85% and induction values >20%, suggesting that they either weakly inhibit or do not inhibit de novo pyrimidine biosynthesis (Supplementary Fig. 22). Compounds E18 and E4, which had moderate similarities to the de novo pyrimidine biosynthesis bioactivity profile (84% and 79% similarity at 50 µM, respectively) and weak induction values (10% and 15% at 50 µM, respectively), were resynthesized and subjected to the uridine rescue assay. At 50 µM concentrations, E18 had weak activity, whereas E4 inhibited cell proliferation but could not be rescued by supplemental uridine and therefore is probably not affecting de novo pyrimidine biosynthesis (Supplementary Fig. 21b,c).
From the CPA subprofile analysis, class F was found to be enriched in profiles that had >85% similarity to the tubulin-targeting cluster (25% of all compounds in class F). Closer examination of these profiles resulted in the identification of six possible tubulin-targeting compounds that characteristically reduce cell count (Fig. 7a,b). A structure–phenotype relationship study74 of class F was conducted by comparing similarity with the tubulin cluster profile and induction relative to concentration (Supplementary Fig. 23). Unsubstituted compound F1 gave a moderate induction value (30%) with a high similarity to the tubulin cluster profile at 30 µM (93% similarity). Installation of a fluoride to the eastern hemisphere either gave similar (F2 and F6) or more potent activity (F4), whereas the introduction of a methyl (F3) or a fused-phenyl (F8) group resulted in lower induction values and low similarities to the tubulin cluster profile relative to F1. Methoxy-substituted compound F5 gave a high tubulin cluster profile similarity at a relatively low concentration of 3 µM. A similar dioxole derivative (F7) retained high tubulin cluster similarity but only at a concentration of 30 µM or higher. Incorporation of a methyl (F15 versus F6) or chloride (F14 versus F4) to the western hemisphere of the scaffold or an α-phenyl ketone (F21 versus F4) led to relatively lower inductions and low tubulin cluster profile similarities. A closely related 2H-isoquinolinone derivative (G4) was also less active than its indoline counterpart (F5).

a, Structures of the six class F compounds that have >85% similarity to the tubulin cluster profile. b, Tubulin cluster profile relative to class F compounds that have >85% similarity to the tubulin cluster profile. The selected profiles are those that have >85% similarity to the tubulin cluster profile and >20% induction at the lowest concentrations per compound. The reference profile is the first profile (100% biosimilarity) for which all subsequent profiles are compared. The tubulin profile has 424 features and is divided into three segments: cell, cytoplasm and nuclei. Biosim, biosimilarity to the tubulin cluster profile; ind, induction (percentage of significantly changed features relative to DMSO controls); and conc, concentration. The induction value reported is relative to the full CPA profiles with 579 features. c, Influence on the microtubule network. U2OS cells were treated for 24 h with DMSO (control) or F5 before staining for tubulin (green) and DNA (blue). Scale bar, 50 µm. d, Quantification of mitotic cells via immunocytochemistry. U2OS cells were treated with DMSO (negative control), F5, nocodazole (noc, positive control) or colchicine (col, positive control) for 24 h before staining of cells for phospho-histone H3 and DNA. Cells in mitosis were quantified as the percentage of phospho-histone H3-positive cells. Data are mean values ± s.d. of three independent replicates (n = 3). Statistical analyses were performed relative to the DMSO control by employing unpaired two-tailed t-tests (****P < 0.0001). The P values relative to cells treated with DMSO are <0.0001 for F5 (10 µM)-treated, <0.0001 for F5 (30 µM)-treated, <0.0001 for noc-treated and <0.0001 for col-treated cells. e, In vitro tubulin polymerization assay. The polymerization was initiated upon addition of guanosine triphosphate (GTP) to porcine tubulin and quantified by means of turbidity measurement at 340 nm and 37 °C. DMSO was used as a negative control and noc was used as a positive control for tubulin destabilization. Data are representative of three independent experiments (n = 3).
Compound F5 was selected for further investigation since it has a good induction value at a relatively low concentration (3 µM) while retaining a high similarity to the tubulin subcluster, indicating that it may be the most potent compound. To further corroborate the tubulin-targeting prediction, the full profile of F5 at 3 µM was compared with the full profiles of reference compounds. Gratifyingly, 11 of the first 12 most similar reference compounds are in the tubulin-targeting bioactivity cluster (Supplementary Fig. 24).
To investigate the predicted bioactivity, U2OS cells were treated with F5 for 24 h followed by immunostaining for tubulin and DNA. Upon treatment of F5, there was an accumulation of round cells with condensed microtubules and multipolar spindles (Fig. 7c), which are consistent with a tubulin-targeting phenotype80. Notably, upon treatment of F5 (10 µM), the abundance of miotic cells increased by ninefold relative to the DMSO control as quantified by means of phospho-histone H3-positive cells (Fig. 7d). A concentration-dependent response was observed with a nearly two-fold increase in mitotic cells upon treatment with 10 μM to 30 μM of F5. Additionally, compound F5 suppressed in vitro tubulin polymerization at 20 μM and parallels the activity of the known tubulin polymerization inhibitor nocodazole (Fig. 7e). These results confirm that the PNP F5 is a microtubule modulator and validates the target prediction of the CPA subprofile analysis. Compound F5 was resynthesized and functionalized to afford four additional derivatives (Supplementary Fig. 25a). The bioactivity of F5 was reconfirmed in a tubulin polymerization assay, and the derivatives were either inactive or moderately active relative to the parent compound (Supplementary Fig. 25b). Furthermore, F5 was more potent than F4, and F4 was more potent than F15 (Supplementary Fig. 25b), as was suggested by the structure–phenotype analysis (Supplementary Fig. 23).
Conclusions
In molecular discovery programmes, several design strategies have been applied with the goal of efficiently exploring biologically relevant chemical space. DOS focuses on the rapid synthesis of compound collections that have high chemical diversity with the intention of probing a wide range of biological space; however, the compounds generated may not necessarily be biologically relevant. The PNP principle provides structurally focused collections by combining NP fragments in arrangements not found in existing NP structures to afford NP-like scaffolds that are endowed with the inherent biological relevance of NPs.
We have proposed the diverse PNP concept that combines the chemical diversity of DOS collections with the inherent biological relevance of the PNP concept to afford compound collections that may be enriched with both structural and biological diversity. The diverse PNP concept has been demonstrated by employing a divergent intermediate strategy along with the development and implementation of synthetic methodologies for the rapid access to eight chemically diverse PNP classes. Of note, dearomatization methodologies were employed to convert aromatic ring systems into more three-dimensional moieties with new stereogenic centre(s) and increased fraction of sp3 carbons (Supplementary Fig. 26). The biological diversity of the collection was exemplified by phenotypic screening and morphological profiling in which four compounds from different classes were identified to have unique bioactivities as either an inhibitor of Hh signalling (A23), tubulin polymerization (F5), DNA synthesis (C3) or de novo pyrimidine biosynthesis (E13).
The diverse PNP concept may help guide the future design of NP-like compound collections that embody both chemical diversity and biological relevance. Incorporation of both of these features may lead to high-quality compound collections that more effectively probe diverse biologically relevant chemical space and, in conjunction with suitable screening technologies, lead to the identification of unexpected or novel bioactivities as demonstrated here. Recently, virtual screenings of ultra-large make on-demand compound collections have been used to expand the exploration of biologically relevant chemical space81,82. While this approach can rapidly evaluate billions of compounds and lead to the discovery of new chemotypes, it is limited to known targets and binding sites, whereas assessing compounds, such as the collection described here, in an unbiased manner may lead to the identification of novel targets, binding pockets or molecular glues. Further biological screening campaigns of these and other PNPs may provide context for the biological relevance of PNPs relative to NPs and collections of different designs as well as the importance of chemical diversity within PNP design.
Methods
Procedure for CO insertion/indole dearomatization cascade
An oven-dried screw-capped reaction tube with a magnetic stir bar was charged with 3-(2-bromobenzyl)-2-methyl-1H-indole (0.5 mmol), N-formyl saccharin (2a, 1.5 equiv., 0.75 mmol), Pd(OAc)2 (3 mol%, 0.015 mmol, 3.4 mg), Xantphos (4.5 mol%, 13 mg) and Na2CO3 (3 equiv., 159 mg). A screw cap fitted with a rubber septum was attached to the reaction tube then degassed and refilled with argon; the process was repeated two additional times. Dry DMF (5 ml) was added to the reaction tube, and the reaction tube was placed in metal block and vigorously stirred at room temperature for 10 min followed by heating to 85 °C. The reaction mixture was cooled to room temperature after 36 h, diluted with 10 ml of ethyl acetate and 25 ml of water. The organic layer was separated and concentrated under vacuum. The crude mixture was purified by column chromatography using silica gel (100–200 mesh size) and petroleum ether/ethyl acetate as the eluent.
Cheminformatic analyses
All details related to the cheminformatic analyses can be found in the repository at ref. 84.
Biological assays
All details pertaining to the biological assays can be found in Supplementary Information.
Primers used for RT–qPCR:
Primer | Forward (5′–3′) | Reverse (5′–3′) |
|---|---|---|
Ptch1 | CTCTGGAGCAGATTTCCAAGG | TGCCGCAGTTCTTTTGAATG |
Gli1 | CACCGTGGGAGTAAACAGGCCTTCC | CCAGAGCGTTACACACCTGCCCTTC |
Alpl | ATCTTTGGTCTGGCTCCCATG | TTTCCCGTTCACCGTCCAC |
Gapdh | CAGTGCCAGCCTCGTC | CAATCTCCACTTTGCCACTG |
Ap3dl | CAGAGGGCTCATCGGTACAC | GCCGGAAGTCCAACTTCTCA |
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The data that support this study are available within the article and Supplementary Information. The reference datasets used on the manuscript (stemming from Enamine Advanced Screening Collection, https://enamine.net/hit-finding/compound-collections/screening-collection/advanced-collection downloaded on 7 December 2020; Drugbank approved and investigational drugs, v.5.1.8; ChEMBL v30) are not included, but the steps to generate them are described in the code repository. All structures covered in this manuscript and their calculated properties are included in the GitHub repository at ref. 84. Crystallographic data for the structure reported in the article have been deposited at the Cambridge Crystallographic Data Centre, under deposition number CCDC 2221540 (B10). Copies of the data can be obtained free of charge via https://www.ccdc.cam.ac.uk/structures. Further datasets generated and analysed during the current study are available from the corresponding authors upon request.
Code availability
The code for the standardization of chemical structures, the extraction of NPs from ChEMBL (one of the reference datasets), the calculation of chemical descriptors for the principal component analysis and for the calculation of PMI is available as a GitHub repository at ref. 84.
References
Dobson, C. M. Chemical space and biology. Nature 432, 824–828 (2004).
Grygorenko, O. O., Volochnyuk, D. M., Ryabukhin, S. V. & Judd, D. B. The symbiotic relationship between drug discovery and organic chemistry. Chem. Eur. J. 26, 1196–1237 (2020).
Newman, D. J. & Cragg, G. M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 83, 770–803 (2020).
Wender, P. A., Verma, V. A., Paxton, T. J. & Pillow, T. H. Function-oriented synthesis, step economy, and drug design. Acc. Chem. Res. 41, 40–49 (2008).
Micalizio, G. C. & Hale, S. B. Reaction design, discovery, and development as a foundation to function-oriented synthesis. Acc. Chem. Res. 48, 663–673 (2015).
Crane, E. A. & Gademann, K. Capturing biological activity in natural product fragments by chemical synthesis. Angew. Chem. Int. Ed. 55, 3882–3902 (2016).
Wetzel, S., Bon, R. S., Kumar, K. & Waldmann, H. Biology-oriented synthesis. Angew. Chem. Int. Ed. 50, 10800–10826 (2011).
Van Hattum, H. & Waldmann, H. Biology-oriented synthesis: harnessing the power of evolution. J. Am. Chem. Soc. 136, 11853–11859 (2014).
Huffman, B. J. & Shenvi, R. A. Natural products in the ‘marketplace’: interfacing synthesis and biology. J. Am. Chem. Soc. 141, 3332–3346 (2019).
Woo, S. & Shenvi, R. A. Natural product synthesis through the lens of informatics. Acc. Chem. Res. 54, 1157–1167 (2021).
McLeod, M. C. et al. Probing chemical space with alkaloid-inspired libraries. Nat. Chem. 6, 133–140 (2014).
Pye, C. R., Bertin, M. J., Lokey, R. S., Gerwick, W. H. & Linington, R. G. Retrospective analysis of natural products provides insights for future discovery trends. Proc. Natl Acad. Sci. USA 114, 5601–5606 (2017).
Grigalunas, M., Burhop, A., Christoforow, A. & Waldmann, H. Pseudo-natural products and natural product-inspired methods in chemical biology and drug discovery. Curr. Opin. Chem. Biol. 56, 111–118 (2020).
Alkubaisi, B. O. et al. Complexity-to-diversity and pseudo-natural product strategies as powerful platforms for deciphering next-generation therapeutics. ChemMedChem 18, e202300117 (2023).
Huigens, R. W. et al. A ring-distortion strategy to construct stereochemically complex and structurally diverse compounds from natural products. Nat. Chem. 5, 195–202 (2013).
Morrison, K. C. & Hergenrother, P. J. Natural products as starting points for the synthesis of complex and diverse compounds. Nat. Prod. Rep. 31, 6–14 (2014).
Motika, S. E. & Hergenrother, P. J. Re-engineering natural products to engage new biological targets. Nat. Prod. Rep. 37, 1395–1403 (2020).
Karageorgis, G., Foley, D. J., Laraia, L. & Waldmann, H. Principle and design of pseudo-natural products. Nat. Chem. 12, 227–235 (2020).
Karageorgis, G., Foley, D. J., Laraia, L., Brakmann, S. & Waldmann, H. Pseudo natural products—chemical evolution of natural product structure. Angew. Chem. Int. Ed. 60, 15705–15723 (2021).
Grigalunas, M., Brakmann, S. & Waldmann, H. Chemical evolution of natural product structure. J. Am. Chem. Soc. 144, 3314–3329 (2022).
Akbarzadeh, M. et al. The pseudo-natural product rhonin targets RHOGDI. Angew. Chem. Int. Ed. 61, e202115193 (2022).
Grigalunas, M. et al. Unprecedented combination of polyketide natural product fragments identifies the new Hedgehog signaling pathway inhibitor grismonone. Chem. Eur. J. 28, e202202164 (2022).
Davies, C. et al. Identification of a novel pseudo‐natural product type IV IDO1 inhibitor chemotype. Angew. Chem. Int. Ed. 61, e202209374 (2022).
Yao, R. et al. Identification of 5-HT2 serotonin receptor modulators through the synthesis of a diverse, tropane- and quinuclidine-alkaloid-inspired compound library. J. Med. Chem. 66, 11536–11554 (2023).
Liu, J. et al. Design, synthesis, and biological evaluation of chemically and biologically diverse pyrroquinoline pseudo natural products. Angew. Chem. Int. Ed. 60, 4648–4656 (2021).
Grigalunas, M. et al. Natural product fragment combination to performance-diverse pseudo-natural products. Nat. Commun. 12, 1883 (2021).
Liu, J. et al. Combination of pseudo-natural product design and formal natural product ring distortion yields stereochemically and biologically diverse pseudo-sesquiterpenoid alkaloids. Angew. Chem. Int. Ed. 60, 21384–21395 (2021).
Burke, M. D. & Schreiber, S. L. A planning strategy for diversity-oriented synthesis. Angew. Chem. Int. Ed. 43, 46–58 (2004).
Cordier, C., Morton, D., Murrison, S., Nelson, A. & O’Leary-Steele, C. Natural products as an inspiration in the diversity-oriented synthesis of bioactive compound libraries. Nat. Prod. Rep. 25, 719–737 (2008).
Schreiber, S. L. Molecular diversity by design. Nature 457, 153–154 (2009).
Morton, D., Leach, S., Cordier, C., Warriner, S. & Nelson, A. Synthesis of natural-product-like molecules with over eighty distinct scaffolds. Angew. Chem. Int. Ed. 48, 104–109 (2009).
O’ Connor, C. J., Beckmann, H. S. G. & Spring, D. R. Diversity-oriented synthesis: producing chemical tools for dissecting biology. Chem. Soc. Rev. 41, 4444–4456 (2012).
Clemons, P. A. et al. Small molecules of different origins have distinct distributions of structural complexity that correlate with protein-binding profiles. Proc. Natl Acad. Sci. USA 107, 18787–18792 (2010).
Petrone, D. A., Yen, A., Zeidan, N. & Lautens, M. Dearomative indole bisfunctionalization via a diastereoselective palladium-catalyzed arylcyanation. Org. Lett. 17, 4838–4841 (2015).
Zeidan, N., Beisel, T., Ross, R. & Lautens, M. Palladium-catalyzed arylation/heteroarylation of indoles: access to 2,3-functionalized indolines. Org. Lett. 20, 7332–7335 (2018).
Zheng, C. & You, S. L. Catalytic asymmetric dearomatization (CADA) reaction-enabled total synthesis of indole-based natural products. Nat. Prod. Rep. 36, 1589–1605 (2019).
Abou-Hamdan, H., Kouklovsky, C. & Vincent, G. Dearomatization reactions of indoles to access 3D indoline structures. Synlett 31, 1775–1788 (2020).
Liu, Y.-Z., Song, H., Zheng, C. & You, S.-L. Cascade asymmetric dearomative cyclization reactions via transition-metal-catalysis. Nat. Synth. 1, 203–216 (2022).
Bauer, R. A., Wurst, J. M. & Tan, D. S. Expanding the range of ‘druggable’ targets with natural product-based libraries: an academic perspective. Curr. Opin. Chem. Biol. 14, 308–314 (2010).
Lovering, F., Bikker, J. & Humblet, C. Escape from flatland: Increasing saturation as an approach to improving clinical success. J. Med. Chem. 52, 6752–6756 (2009).
Müller, G., Berkenbosch, T., Benningshof, J. C. J., Stumpfe, D. & Bajorath, J. Charting biologically relevant spirocyclic compound space. Chem. Eur. J. 23, 703–710 (2017).
Talele, T. T. Opportunities for tapping into three-dimensional chemical space through a quaternary carbon. J. Med. Chem. 63, 13291–13315 (2020).
Bera, S., Daniliuc, C. G. & Studer, A. Oxidative N-heterocyclic carbene catalyzed dearomatization of indoles to spirocyclic indolenines with a quaternary carbon stereocenter. Angew. Chem. Int. Ed. 56, 7402–7406 (2017).
Bai, Y., Davis, D. C. & Dai, M. Natural product synthesis via palladium-catalyzed carbonylation. J. Org. Chem. 82, 2319–2328 (2017).
Ma, K., Martin, B. S., Yin, X. & Dai, M. Natural product syntheses: via carbonylative cyclizations. Nat. Prod. Rep. 36, 174–219 (2019).
Breuers, C. B. J., Daniliuc, C. G. & Studer, A. Dearomatizing cyclization of 2-iodoindoles by oxidative NHC catalysis to access spirocyclic indolenines and oxindoles bearing an all carbon quaternary stereocenter. Org. Lett. 24, 4960–4964 (2022).
Yasui, M. et al. Synthesis of spiro[indole-3,3′-pyrrolidine]-2′-(thi)ones. J. Org. Chem. 88, 1093–1106 (2023).
Ren, W. & Yamane, M. Mo(CO)6-mediated carbamoylation of aryl halides. J. Org. Chem. 75, 8410–8415 (2010).
Ueda, T., Konishi, H. & Manabe, K. Palladium-catalyzed reductive carbonylation of aryl halides with N-formylsaccharin as a CO source. Angew. Chem. Int. Ed. 52, 8611–8615 (2013).
Tan, Y. et al. N-formylsaccharin as a CO source: applications and recent developments. ChemistrySelect 6, 2343–2349 (2021).
You, S. L. Recent developments in asymmetric transfer hydrogenation with Hantzsch esters: a biomimetic approach. Chem. Asian. J. 2, 820–827 (2007).
Susanti, D., Ng, L. L. R. & Chan, P. W. H. Silica gel-mediated hydroamination/semipinacol rearrangement of 2-alkylaminophenylprop-1-yn-3-ols: synthesis of 2-oxindoles from alkynes and 1-(2-aminophenyl) ketones. Adv. Synth. Catal. 356, 353–358 (2014).
Bu, L. et al. Organocatalytic asymmetric cascade aerobic oxidation and semipinacol rearrangement reaction: a visible light-induced approach to access chiral 2,2-disubstituted indolin-3-ones. Chem. Asian J. 13, 2382–2387 (2018).
RDKit: Open-source cheminformatics v2022.03.5 http://www.rdkit.org
Tanwar, S. et al. A new ChEMBL dataset for the similarity-based target fishing engine FastTargetPred: annotation of an exhaustive list of linear tetrapeptides. Data Brief 42, 108159 (2022).
Wishart, D. S. et al. DrugBank 5.0: a major update to the DrugBank database for 2018. Nucleic Acids Res. 46, D1074–D1082 (2018).
Advanced collection. Enamine https://enamine.net/hit-finding/compound-collections/screening-collection/advanced-collection (2020).
Sauer, W. H. B. & Schwarz, M. K. Molecular shape diversity of combinatorial libraries: a prerequisite for broad bioactivity. J. Chem. Inf. Comp. Sci. 43, 987–1003 (2003).
Ertl, P., Roggo, S. & Schuffenhauer, A. Natural product-likeness score and its application for prioritization of compound libraries. J. Chem. Inf. Model. 48, 68–74 (2008).
Bickerton, G. R., Paolini, G. V., Besnard, J., Muresan, S. & Hopkins, A. L. Quantifying the chemical beauty of drugs. Nat. Chem. 4, 90–98 (2012).
Pedregosa, F. et al. Scikit-learn: machine learning in Python. J. Mach. Learn Res. 12, 2825–2830 (2011).
Doveston, R., Marsden, S. & Nelson, A. Towards the realisation of lead-oriented synthesis. Drug Discov. Today 19, 813–819 (2014).
Drugbank appoved and experimental drugs Drugbank https://www.drugbank.ca/releases/latest#structures (2020).
Wu, F., Zhang, Y., Sun, B., McMahon, A. P. & Wang, Y. Hedgehog signaling: from basic biology to cancer therapy. Cell Chem. Biol. 24, 252–280 (2017).
Hui, C. C. & Angers, S. Gli proteins in development and disease. Annu. Rev. Cell Dev. Biol. 27, 513–537 (2011).
Peukert, S. & Miller-Moslin, K. Small-molecule inhibitors of the hedgehog signaling pathway as cancer therapeutics. ChemMedChem 5, 500–512 (2010).
Wu, X., Walker, J., Zhang, J., Ding, S. & Schultz, P. G. Purporphamine induces osteogenesis by activation of the Hedgehog signaling pathway. Chem. Biol. 11, 1229–1238 (2004).
Kremer, L. et al. Discovery of a novel inhibitor of the Hedgehog signaling pathway through cell-based compound discovery and target prediction. Angew. Chem. Int. Ed. 56, 13021–13025 (2017).
Sinha, S. & Chen, J. K. Purmorphamine activates the Hedgehog pathway by targeting Smoothened. Nat. Chem. Biol. 2, 29–30 (2006).
Chen, J. K., Taipale, J., Cooper, M. K. & Beachy, P. A. Inhibition of Hedgehog signaling by direct binding of cyclopamine to Smoothened. Genes Dev. 16, 2743–2748 (2002).
Yauch, R. L. et al. Smoothened mutation confers resistance to a Hedgehog pathway inhibitor in medulloblastoma. Science 326, 572–574 (2009).
Bray, M.-A. et al. Cell Painting, a high-content image-based assay for morphological profiling using multiplexed fluorescent dyes. Nat. Protoc. 11, 1757–1774 (2016).
Caicedo, J. C., Singh, S. & Carpenter, A. E. Applications in image-based profiling of perturbations. Curr. Opin. Biotechnol. 39, 134–142 (2016).
Christoforow, A. et al. Design, synthesis, and phenotypic profiling of pyrano-furo-pyridone pseudo natural products. Angew. Chem. Int. Ed. 58, 14715–14723 (2019).
Ziegler, S., Sievers, S. & Waldmann, H. Morphological profiling of small molecules. Cell Chem. Biol. 28, 300–319 (2021).
Pahl, A. et al. Morphological subprofile analysis for bioactivity annotation of small molecules. Cell Chem. Biol. 30, 839–853 (2023).
Salic, A. & Mitchison, T. J. A chemical method for fast and sensitive detection of DNA synthesis in vivo. Proc. Natl Acad. Sci. USA 105, 2415–2420 (2008).
Schölermann, B. et al. Identification of dihydroorotate dehydrogenase inhibitors using the cell painting assay. ChemBioChem 23, e202200475 (2022).
Wu, H. L. et al. Targeting nucleotide metabolism: a promising approach to enhance cancer immunotherapy. J. Hematol. Oncol. 15, 45 (2022).
Akbarzadeh, M. et al. Morphological profiling by means of the cell painting assay enables identification of tubulin-targeting compounds. Cell Chem. Biol. 29, 1053–1064 (2022).
Lyu, J. et al. Ultra-large library docking for discovering new chemotypes. Nature 566, 224–229 (2019).
Sadybekov, A. A. et al. Synthon-based ligand discovery in virtual libraries of over 11 billion compounds. Nature 601, 452–459 (2022).
Tukey, J. W. Explanatory Data Analysis (Addison-Wesley, 1977).
mpimp-comas/2023_bag_bio_diverse_pnps: revision. Zenodo https://doi.org/10.5281/zenodo.8320827 (2023).
Acknowledgements
We acknowledge the Max-Planck Gesellschaft for financial support. S.B. acknowledges the Alexander von Humboldt Stiftung for a post-doctoral fellowship. This work was co-funded by the European Union (Drug Discovery Hub Dortmund, EFRE-0200481) and Innovative Medicines Initiative (grant agreement number 115489), the resources of which are composed of financial contribution from the European Union’s Seventh Framework Programme (FP7/2007-2013) and European Federation of Pharmaceutical Industries and Associations (EFPIA) companies’ in-kind contribution and the programme ‘Netzwerke 2021’, an initiative of the Ministry of Culture and Science of the State of North Rhine Westphalia. S.B. acknowledges F. Sörries, R. Borgmeier and P. Brech for helping in compound synthesis.
Funding
Open access funding provided by Max Planck Society.
Author information
Authors and Affiliations
Contributions
S.B., M.G. and H.W. conceived and designed the project. S.B. and J.L. performed the chemical experiments and analysed the data. L.W. helped with insightful discussions related to library expansion. A.P. and M.G. performed and/or analysed the cheminfomatic analyses. S.S. and A.P. performed the CPA and/or processed the data. S.B. and M.G. analysed the cell painting data. S.B., J.L., S.P., J.B., S.K., B.S., R.Z., S.S. and S.Z. performed and/or analysed the biological experiments. L.B. and C.S. conducted X-ray diffraction (XRD) and analysed the results. S.B., M.G. and H.W. prepared the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Chemistry thanks Oleksandr Grygorenko and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary Information
Supplementary Tables 1–13; Figs. 1–27; cheminformatic, biological experimental and chemistry experimental details; and compound characterization data and spectra.
Supplementary Data 1
Crystallographic data for compound B10; CCDC reference 2221540.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bag, S., Liu, J., Patil, S. et al. A divergent intermediate strategy yields biologically diverse pseudo-natural products. Nat. Chem. (2024). https://doi.org/10.1038/s41557-024-01458-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41557-024-01458-4
