
Abstract
Adaptive evolution and phenotypic plasticity will fuel resilience in the geologically unprecedented warming and acidification of the earth’s oceans, however, we have much to learn about the interactions and costs of these mechanisms of resilience. Here, using 20 generations of experimental evolution followed by three generations of reciprocal transplants, we investigated the relationship between adaptation and plasticity in the marine copepod, Acartia tonsa, in future global change conditions (high temperature and high CO2). We found parallel adaptation to global change conditions in genes related to stress response, gene expression regulation, actin regulation, developmental processes, and energy production. However, reciprocal transplantation showed that adaptation resulted in a loss of transcriptional plasticity, reduced fecundity, and reduced population growth when global change-adapted animals were returned to ambient conditions or reared in low food conditions. However, after three successive transplant generations, global change-adapted animals were able to match the ambient-adaptive transcriptional profile. Concurrent changes in allele frequencies and erosion of nucleotide diversity suggest that this recovery occurred via adaptation back to ancestral conditions. These results demonstrate that while plasticity facilitated initial survival in global change conditions, it eroded after 20 generations as populations adapted, limiting resilience to new stressors and previously benign environments.
Similar content being viewed by others


Introduction
Global conditions are changing at a geologically unprecedented rate, and the frequency of extreme events is increasing due to human activities resulting in chronic and novel environmental changes1. However, species can use genetic adaptation, physiological plasticity, or both to persist2,3,4. Genetic adaptation, heritable genetic change that improves the mean fitness of a population in an environment5, enables resilience by shifting the mean phenotype of the population to tolerate different conditions. Selection can shift population genetic variation over short time-scales, as few as one to four generations, in response to an extreme change in the environment6,7,8,9. Conversely, plasticity allows a single genotype to generate different phenotypes in response to the environment10. Considering rapid changes in environmental conditions, adaptive plasticity enables organisms to maintain fitness across environments11 through underlying phenotypic changes, such as shifts in metabolism or gene regulation12. However, the interaction and relative contributions of adaptation and plasticity to population persistence under rapid environmental change remains a critical area of investigation13,14,15,16.
Theoretical models predict that plasticity may increase when a population is exposed to a novel environment, but should decline across longer timeframes17. This reduction is expected when the previous trigger for plasticity is no longer present, the environment is static, or the plasticity is costly enough to be selected against18. Indeed, a reduction of plasticity appears to be common in natural and experimental empirical systems19,20,21,22,23. This experimental work has also revealed faster losses of plasticity during rapid adaptation than theory would predict22,24. For example, Waddington showed that a high temperature-induced plastic phenotype, crossveins in Drosophila wings, can become fixed, or assimilated, within 20 generations of artificial selection, reducing plasticity and the environmental responsiveness of a trait25. Under climate change conditions, populations will experience strong selection, and it is essential to understand how selection in these stressful conditions may alter or reduce the plasticity of natural populations.
Experimental evolution is a particularly powerful approach to understand resilience as organisms can be exposed to specific selective pressures over multiple generations26. From this, one can identify both the adaptive potential and the mechanisms of adaptation of a population, directly providing insight into how a population may cope with changing climates27. Experimental evolution in fruit flies28,29, copepods21, marine algae30, nematodes22, mice31, and others32 have identified phenotypic or genomic responses to selection and have demonstrated that strong selection can rapidly shift or drive a loss of plasticity. The majority of these studies have leveraged model systems and found that when selection regimes are static, plasticity tends to be lost (but see refs. 29,33). While the static conditions in the lab are a limitation of these studies as they do not necessarily represent the variable conditions found in the wild, experimental losses of plasticity under static conditions raise the concern that the strong selection driving rapid adaptation to global change may also purge phenotypic plasticity from populations. Indeed, a loss of plasticity appears to be present in natural systems rapidly adapting, for example, as seen in marine to freshwater transitions in copepods19 and numerous fish species23,34,35, populations of plants and frogs as they adapt to reduced habitat heterogeneity36,37,38, fishes in different predation environments39, and adaptive radiations of cichlids with different prey items40.
The evolution of phenotypic plasticity is dictated by both the degree and predictability of environmental variability relative to generation time; variability must be experienced within the lifespan of an individual to favor plasticity41. Under global change, it is expected that conditions such as prolonged heat waves1, decreasing salinity42, and warmer winter conditions43 will become more common. For short-lived species, these extreme conditions may be longer than their generation time and therefore selection may drive rapid adaptation that reduces plasticity, resulting in populations that are less resilient when environments fluctuate in the future, which is expected under global change1. However, there have been limited experimental evolution studies investigating the link between adaptive evolution and plasticity in non-model metazoans, particularly in multiple stressor scenarios. Further, work focused on understanding the phenotypic plasticity of experimentally evolved populations across multiple generations of reciprocal transplant in common gardens is lacking. Such experiments could test the effects of long-term adaptation and its impacts on plasticity. Incorporating this multigenerational reciprocal common garden perspective after adaptation is essential to better understand if and how organisms can shift phenotypes following rapid environmental change.
The use of outbred natural populations in evolution experiments is a powerful approach for revealing the potential contributions of standing genetic variation and plasticity, particularly in context of future global change conditions44,45. Species that live in dynamic environments, have distributions spanning a wide range of environmental conditions, and have high dispersal capacity are predicted to have the greatest capacity to respond to rapid environmental change due to their high levels of standing genetic variation and physiological plasticity46. Many marine species in particular live in environments that vary across time and space and have life histories that promote dispersal47,48,49. This is true for copepods, the most abundant marine metazoans50, which have plasticity and genetic variation for adaptation to global change conditions21,51,52,53,54. However, few studies have measured the genetic response during adaptive evolution to a multiple stressor environment across multiple generations in a marine metazoan and none, to our knowledge, across any taxa, have measured allelic and gene expression responses across multiple generations of reciprocal transplant following adaptation. Further, the relationship between plasticity and genetic adaptation is only beginning to be understood in marine systems33,55,56. Given their ecological importance and high levels of genetic variation and plasticity, copepods are ideal models to disentangle the interplay of genetic and plastic responses of natural populations of metazoans to global change conditions24,52.
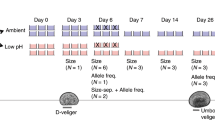
We experimentally evolved then reciprocally transplanted the globally distributed, foundational copepod, Acartia tonsa, to ambient and global change conditions to determine the effects of long-term adaptation on plasticity. This coupling of many generations of experimental evolution to a multi-stressor environment with multiple generations of reciprocal transplant is a first to our knowledge. As a dominant prey item for forage fish, A. tonsa serves as a critical link in the marine food web, supporting economically important fisheries57 and as a major phytoplankton grazer contributes to the storage of atmospheric CO2 and thus mediates marine biogeochemical cycles58. Replicate cultures (4 replicates per condition, ~4000 individuals per replicate) of A. tonsa were subjected to 20 generations of selection in ambient (AM: 400 ppm pCO2 and 18 °C) and combined high CO2 × temperature (ocean warming and acidification, OWA: 2000 ppm pCO2 and 22 °C) conditions followed by three generations of reciprocal transplantation of both lines to opposite conditions (Fig. 1). These conditions were chosen as they represent present-day and a worst-case, yet realistic scenario;59,60 in the coming century, global mean ocean surface temperatures will increase by 2–4 °C61 and oceanic CO2 concentrations will potentially reach 2000 ppm by the year 230062. We denote evolved lines with their abbreviation and the environmental condition during transplant as a subscript (i.e., AMOWA = ambient line transplanted to warming and acidification conditions). RNA was collected from pools of 20 adults from each of four replicate cultures at the first, second, and third generation after transplant for both experimentally evolved and transplanted lines (Fig. 1). We estimated allele frequencies at 322,595 variant sites with at least 50× coverage across all samples (mean coverage 174×) and quantified transcript abundance for an average 24,112 genes.

Blue lines are ambient (AM) pH and temperature that represent current conditions (pCO2: 400 ppm; temperature: 18 °C). Red lines are simulated future warming and acidification (OWA) conditions (pCO2: 2000 ppm; 22 °C). Adult Acartia tonsa were collected from the wild and reared in the lab for three generations. Six hundred laboratory-acclimated adults seeded each of four replicates at AM and OWA conditions where they were reared for 20 non-overlapping generations. At generation 20, each replicate was split in two and transplanted into the same conditions as the previous 20 generations (AMAM, OWAOWA) and to the opposite condition (AMOWA, OWAAM). These transplanted lines were reared for three additional generations and sampled for life-history traits at the first generation and genomics at the end of each generation.
We have previously shown that the OWA lines used here rapidly adapted to OWA conditions by recovering egg production and hatching success63. By reciprocal transplantation of control and OWA-adapted lines, we assess how selection in warming and acidification influenced ancestral plasticity to tolerate environmental changes and reveal the fate of adaptive phenotypic plasticity under rapid environmental change. If directional selection decreases plasticity in a population, it may reduce resilience if the environment shifts again in the future. Similarly, strong selection for resilience to one stressor may have antagonistic effects on the resilience to other stressors. In the oceans, nutrient limitation may increase with rising temperatures64, raising the question of how adaptation to temperature may influence resilience to nutrient limitation. We address three major questions in this study. (1) What are the allelic and gene expression mechanisms underlying adaptation to OWA conditions? (2) To what extent is genetic adaptation vs. transcriptional plasticity required to respond to environmental change? Specifically, does adaptation result in a loss of plasticity in a marine metazoan? (3) Does adaptation result in trade-offs among life-history traits revealed through reciprocal transplant and in low-food stress conditions? We predicted that this coastal copepod would have genetic and plastic mechanisms to respond to warming and acidification conditions and that long-term adaptation would reduce physiological plasticity. Our results support these predictions, revealing the molecular underpinnings of the adaptive mechanisms, and demonstrate a loss of transcriptional plasticity with a capacity for recovery at the expense of adaptive genetic variation.
Results and discussion
Divergence between selected lines
Genetic and transcriptional variation diverged consistently across replicate cultures after 20 generations in warming and acidification vs. ambient conditions (Fig. 2, Supplementary Figs. 1 and 2), supporting our previous results that show rapid adaptation to warming and acidification conditions through selection on hatching success and egg production63. A total of 17,720 loci (5.5% of 322,595 loci with >50x coverage) showed consistent allele frequency divergence (OWAOWA vs. AMAM; Cochran–Mantel–Haenszel test false discovery rate-corrected significance threshold: 5.17e-8). Given the likely polygenic nature of adaptation to warming and acidification conditions and that polygenic traits tend to be genetically redundant65,66, the identified adaptive genetic variation would be prone to false negatives due to the requirement that adaptation be parallel in all replicates. Conversely, linkage disequilibrium contributes some number of false positives when neutral loci are located near the selected loci67. Of potential candidate genes driving adaptation (Supplementary Fig. 2), one gene, NADH-ubiquinone oxidoreductase 49 kDa subunit (NDUFS2), was an extreme outlier for divergence in allele frequencies, 77% change, which was 72% greater than the global average (5% change in allele frequency; Supplementary Fig. 3). NDUFS2 is a nuclear-encoded, core subunit of Complex 1 of the mitochondrial electron transport chain (ETC) that has been shown to be inhibited by heat stress68,69. In addition, mitochondrial function has been linked to variation in thermal tolerance among populations of the intertidal copepod Tigriopus californicus70. The three substitutions identified here were all non-synonymous, in linkage disequilibrium, and in a single alpha helix of the ETC subunit, including an Isoleucine, non-reactive hydrophobic amino acid, to Asparagine, polar, often active amino acid (Supplementary Fig. 2). This alternative NDUFS2 haplotype targeted by selection could improve energy production under heat stress for global change-adapted copepods, a hypothesis worth future functional investigation71.

Principal component analysis of (a) genome-wide variation in allele frequencies (322,595 SNPs) and (b) gene expression (24,927 genes) at the F1 generation. c, d Gene ontology enrichment results from one-sided Mann–Whitney U-test using P-values from Cochran–Mantel–Haenszel tests (allele frequencies, c) and DESeq2 Wald tests (gene expression, d). Gene categories are collapsed for visualization purposes with the number of categories indicated in parentheses. See Supplementary Data 3, 4 and Supplementary Figs. 9 and 10 for full results. Source data are provided as a Source Data file.
A total of 1876 genes (7% of the 24,927 genes surveyed) were differentially expressed between control and experimentally evolved replicate lines in their home environment, differences that could be driven by plastic and evolved mechanisms (OWAOWA vs. AMAM; P < 0.05; Supplementary data 1). There was substantial overlap in the functional classes of genes that had evolved allele frequency and gene expression differences. Shared categories included response to and detection of stress and stimuli, developmental processes, actin organization and regulation, and proteolytic processes (Fig. 2c, d and Supplementary Data 2, 3). These functions may underlie increased survival in warmer and more acidic conditions. For example, proteolysis is integral to the stress response72 while actin modifications have been identified as targets of pH adaptation and resilience in other marine organisms9,73 and are linked to cytoskeleton maintenance that is responsive to low pH stress in copepods74. Despite shared functions, there was little overlap in the specific genes that evolved allele frequency and gene expression differences (16%; Fisher’s exact test; P > 0.05), suggesting that adaptive divergence in protein function and gene expression targeted different, but functionally related genes75. Similarly, there was no correlation between the degree of expression and allelic responses (Supplementary Fig. 4; R2 = 0.002), suggesting no impact of potential allele-biased expression on estimates of allele frequencies from pools of sequenced RNA. We verified the lack of impact of allele-biased expression on allele frequency estimates by comparing to allele frequency estimates from genomic DNA from the non-transplanted lines at F2563(Supplemental Information, Supplementary Fig. 5). While there was no spurious technical link between gene expression and genetic divergence, we found strong allele frequency divergence in regulators of expression, predominantly in genes relating to RNA processing (Fig. 2c) and also ribosomal S6 kinase, a gene involved in the regulation of numerous transcription factors and translation (Supplementary Fig. 2)76. Taken together, these results suggest that regulators of gene expression were under selection and may have been responsible for the adaptive changes in gene expression under global change conditions, in accord with previous work in copepods53.
Impacts of adaptation on plasticity
Selection in a stressful environment could increase or decrease plasticity17. We quantified gene expression plasticity to test the prediction that rapid adaptation to a stressful environment would result in a change in physiological plasticity in OWA animals. We identified plastic changes in gene expression for OWA and AM lines by comparing expression in the home environment after 20 generations to gene expression responses at the end of one generation in transplant conditions (AMAM vs. AMOWA and OWAAM vs. OWAOWA; Fig. 3a). At the first generation of transplant, AM lines showed plastic expression responses for 4719 genes (Padj < 0.05; 20% of genes), a 12.7-fold greater response than for OWA lines (372 genes; 1.6% of genes). However, at the end of the second and third generation in transplant, this difference diminished due to the relative increase of OWA gene expression changes (F2: 4.5%; F3: 4.1%) and decrease of AM changes (F2: 1.7%; F3: 6.3%).

a Gene expression plasticity for AM and OWA transplanted lines as defined by differentially expressed genes within a line following transplant (AMAM vs. AMOWA; OWAOWA vs OWAAM). b Comparison of plastic changes between AM and OWA lines. Color and shape indicate differentially expressed genes by one or both lines. If gene expression plasticity in response to the environment is equal between the two lines, the slope of the relationship would be 1 (dashed black line). The observed slope is shown as the solid black line. c Plastic vs. genetic changes in gene expression. Bar plots show the relative number of plastic vs. genetic changes in expression between AMAM and OWAOWA where the dashed line indicates equal numbers. AM to OWA indicates AM lines moving to OWA conditions (Forward adaptation), OWA to AM is the opposite (Reverse adaptation). Inset plots show the counts of the genes that went into the barplot where PO = plasticity only and GC = genetic change. All proportions of PO to GC significant at P < 0.001, two-sided G-test of independence. Source data are provided as a Source Data file.
To understand the relative expression patterns of the plastic genes, we compared gene expression plasticity in OWA vs. AM lines in response to each environment (measured as log2 fold change: AMOWA vs. AMAM and OWAOWA vs. OWAAM; Fig. 3b). When comparing expression patterns in response to environment, the expectation for equal transcriptional plasticity and equal response to environment for OWA and AM lines would be a slope of one among genes differentially expressed. If there was a reduction in plasticity, perhaps due to genetic assimilation in the evolved line (OWA), the relationship would show a slope less than one19,29, indicating a muted transcriptional response by the OWA line. After one generation in transplant conditions, there was a slight negative relationship, slope of −0.25 (Fig. 3b, top panel). A negative relationship between expression responses could be interpreted as non-adaptive gene expression20 or as a consistent response to the act of transplant, rather than environment, between the lines, i.e., a shared stress response to transplant and an opposite response to environmental condition. In the latter case, equal gene expression plasticity in response to transplant between the lines would yield a slope of −1. In the second generation of transplant, the negative relationship persisted (slope = −0.29), but by F3 there was a positive relationship where each transplanted line converged on the gene expression profile of its new environment (slope = 0.37). However, the slope was less than 1, indicating that AM lines had a greater change in expression by F3 than OWA lines and had higher plasticity to change gene expression profiles in response to their new environment. While expression changes at F1 were likely due to plasticity, differential gene expression at F2 and F3 could be due to plasticity or evolution post-transplant. For example, transgenerational plasticity may cause the gene expression at F1 and F2 to be influenced by the environment of the previous generations54,77,78, but environmental effects decay after 3 generations of common garden79. Therefore, we would expect F1 and F2 to reflect plastic responses influenced by the previous environment, which could explain the inverse relationship at these generations. By F3, the environmental effects of the original environment are no longer present and the plastic response remains, which was reduced in the OWA lines relative to AM (Fig. 3b). Additionally, genetic adaptation across the generations following transplant could drive a shift in gene expression patterns53. Together, these results show a pattern of successive changes across each generation and we disentangle the relative impacts of plasticity and genetic adaptation below. The loss of plasticity with adaptation to the higher temperature of the OWA environment matches predictions from natural populations of A. tonsa. Populations sampled along a latitudinal gradient show a trade-off between thermal tolerance and degree of developmental plasticity80. The same pattern is observed in A. tonsa populations across seasons81.
Gene expression patterns after long-term experimental evolution and reciprocal transplantation also allow us to test the hypothesis that transcriptional plasticity can be retained and serve as a mechanism to facilitate re-adaptation or transgenerational acclimation to ancestral conditions in the context of adaptation to global change conditions and extreme events. Using the approach described in Ho et al.82, first we identified the genes differentially expressed between AMAM and OWAOWA, changes likely to be adaptive in each environment. These genes were further characterized as requiring “plasticity only” or requiring “genetic change” for both forward adaptation, AM to OWA conditions, and reverse adaptation, OWA to AM conditions, by comparing the relative expression levels in home vs. transplant environments. Specifically, for the forward direction, genes were considered plasticity only (PO) if they were differentially expressed between AMAM vs. AMOWA and not differentially expressed between AMOWA vs. OWAOWA. Genes were considered to require genetic change (GC) if they were differentially expressed between AMOWA vs OWAOWA. PO and GC genes were mutually exclusive but did not include all of the genes differentially expressed between AMAM and OWAOWA (F1: 87% of total; F2: 66%; F3: 80%). Last, we similarly categorized the differentially expressed genes as PO or GC for the reverse adaptation from warming and acidification to ancestral ambient conditions: PO: differentially expressed between OWAOWA vs. OWAAM and not differentially expressed between OWAAM and AMAM; GC: differentially expressed between OWAAM vs. AMAM.
After 20 generations of experimental evolution and one generation of reciprocal transplantation, forward adaptation, ambient to OWA conditions, had more plasticity-only genes (Fig. 3c; 537 genes) than genetic-change-needed genes (Fig. 3c; 339 genes). In contrast, reverse adaptation, i.e., returning to ambient conditions after 20 generations of OWA adaptation, showed the opposite pattern with substantially more genetic-change needed (Fig. 3c; 613 genes) than plasticity genes (Fig. 3c; 122 genes). These results suggest that forward adaptation to OWA conditions was likely initially possible using plasticity (in this case, the plasticity was maintained in the ambient control lines), and that reverse adaptation, after 20 generations in constant conditions, had little plasticity remaining and required genetic change. These results are in contrast to what Ho et al.82 found in high-altitude adapted chickens, natural and introduced guppy populations, and experimental evolution in E. coli where genetic-change genes were more common when moving to the new environment and plasticity-only genes were more common when going back to the original environment. After two and three generations of transplant, however, our results match Ho et al.’s patterns: reverse adaptation, OWA to AM, had more plasticity-only genes than genetic-change-needed genes, while forward adaptation, AM to OWA, had more genetic-change-needed than plasticity only genes (Fig. 3c). It is important to note that F2 and F3 animals had been in the transplant environment for 2 and 3 generations, respectively; gene expression responses could be due to transgenerational plasticity or rapid evolution post-transplant. Hence, we show that phenotypic plasticity does not always ease adaptation back to ancestral environments. Instead, it can facilitate adaptation to a stressful environment and be rapidly lost, in accord with theoretical work17, and recover via transgenerational or genetic mechanisms, explored next.
Relationship between gene expression and allele frequencies
To explore the relationship between gene expression and allele frequency changes across transplant generations, we used discriminant analysis of principal components (DAPC). Discriminant function space was generated from AMAM and OWAOWA, representing the adaptive expression differences after 20 generations in each environment (Fig. 4, blue and red shaded zones; Supplementary Data 1). We assume that consistent divergence in expression between lines in their home environments is indicative of an adaptive expression profile in that environment (see methods20). Transplanted lines were fit to discriminant function space to determine if and how expression changed to match the adaptive expression profile where the adaptive plasticity for each line is summarized by the length of the arrow in Fig. 4a. Both transplanted lines significantly moved in discriminant function space towards the non-transplanted counterpart across all three generations (Markov chain Monte Carlo (MCMC) generalized linear mixed model; PMCMC F1AM < 4e-04; F2AM = 0.0007; F3AM < 4e-04; F1OWA = 0.001; F2OWA = 0.004; F3OWA < 4e-04). However, matching the results above, we found lower overall plasticity in OWA transplanted copepods relative to AM transplanted copepods at F1; expression changed to a greater extent for AM as compared to OWA lines at F1 (PMCMC = 0.03; Fig. 4a). However, by F2 and F3, AM and OWA lines demonstrated similar shifts where lines converge on the adaptive expression profile by the F3 generation (PMCMC > 0.05; Fig. 4a). Alternative methods based on DeSeq2 differential expression recapitulate these sequential adaptive expression shifts (Supplementary Fig. 6). This increase in adaptive gene expression across generations provides additional evidence that copepods, even after 20 generations of experimental evolution in ambient or warming and acidification conditions and despite differences in initial levels of plasticity, were able to match adaptive gene expression profiles increasingly with each successive generation.

Discriminant analysis of principal components across three transplant generations for a gene expression and b allele frequencies. The x-axis shows the discriminant function space that maximizes differences between lines in their home environments and the background shading represents the “home” discriminant function space for each line. Shape and color of points indicate selection line and treatment condition, respectively. Small points are individual replicates while the mean change for each group is represented by the large transparent points; arrows connect home to transplant means. Significantly different shifts between lines are represented by black arrows and asterisks. Source data are provided as a Source Data file.
Using a similar DAPC approach with allele frequencies, we tracked shifts in allele frequencies towards adaptive alleles following transplant for each successive generation (Fig. 4b). Across all three generations, allele frequency changes within OWA were highly significant following transplant (PMCMC F1 = 0.001; F2 = 0.001; F3 = 0.0007) while AM shifts were somewhat less significant (F1 PMCMC = 0.006; F2 = 0.003; F3 = 0.03). At generations F1 and F2, we found allele frequency changes were similar between AM and OWA lines (PMCMC > 0.05; Fig. 4b). However, by F3 transplanted OWA animals showed significantly greater adaptive evolution than AM (PMCMC = 0.02). These shifts in allele frequencies were again mirrored by an independent analysis based on Cochran–Mantel–Haenszel tests (Supplementary Fig. 6). Together with the gene expression analysis, these results indicate that AMOWA animals were able to match the warming and acidification adaptive transcriptional profiles through physiological and/or transgenerational plasticity and minor changes in allele frequencies. Conversely, the OWA animals had dramatically reduced transcriptional plasticity (Figs. 3a–c and 4a, top panels) and seem to have matched the adaptive transcriptional profile through evolution, i.e., via selective changes in allele frequencies across the three generations of transplant (Figs. 3a–c and 4b, bottom panels). Future work should explore the relationship between genetic and epigenetic changes and resulting gene expression differences.
Costs of adaptation
Rapid adaptation can carry costs at the genetic and phenotypic levels. For example, a selective bottleneck can drive a loss of genetic diversity during long-term selection or short-term transplant. To test for this possibility, we quantified nucleotide diversity (π) for each treatment following 20 generations of selection and after transplant. We found no significant loss of genetic diversity after long-term adaptation to warming and acidification conditions (OWAOWA, Fig. 5a, PTukey > 0.05). Similarly, ambient lines, both transplanted and not, maintained high levels of genetic diversity (PTukey > 0.05). However, when transplanted back to ambient conditions, OWAAM showed a significant drop in diversity by F3 (PTukey < 0.05; Fig. 5a), indicating a global loss of genetic diversity. This suggests that the successive shifts in allele frequency after transplant of OWA to AM (OWAAM; Fig. 4b) resulted in a loss of genetic diversity (Fig. 5a). Taken together, these results indicate that matching gene expression to the environment after transplant (OWA to AM) was driven to some degree by adaptation across multiple generations rather than plasticity alone. To test if the loss of genetic diversity after transplant from OWA to AM was concentrated in specific regions of the genome, we compared changes in nucleotide diversity in regions identified as adaptively diverged between warming and acidification and ambient lines (i.e., regions containing 17,720 consistently divergent loci) vs. all other loci (“non-adaptive”). For OWAAM lines, the loss of nucleotide diversity was concentrated in genomic regions containing adaptive loci (Fig. 5b; Wilcoxon test P = 0.007), while there was no loss of diversity in AMOWA or OWAOWA lines as control contrasts (Fig. 5b; P = 0.16; P = 0.07). In addition, the loss of nucleotide diversity for OWAAM lines was concentrated in genes with functions related to sequestration of actin monomers, cytokinesis, and response to stress (Fig. 5c and Supplementary Data 4), similar to those underlying adaptive genetic divergence between AMAM and OWAOWA lines (Fig. 2 and Supplementary Data 2). AMOWA and OWAOWA showed no functional enrichment for regions losing genetic diversity. The lack of nucleotide diversity loss and the absence of functional enrichment indicates that selection was a weak force for AMOWA lines, at least over short periods, which is consistent with the sustained ability for a plastic response in AM lines.

a Median nucleotide diversity (π) for all treatments and generations following transplant (n = 4 biologically independent replicates per treatment and generation). Boxes represent the upper and lower 25% quantiles and median while whiskers are min and max. Letters above each point show significance from Tukey post hoc comparisons following an ANOVA where P < 0.05; b Mean change in π from after transplant for adaptive and non-adaptive SNPs with 95% confidence intervals (n = 9177 π estimates per replicate). Changes relative to AMAM F1. Letters indicate significance from two-sided Wilcoxon tests with a Bonferroni correction and 0.05 significance threshold. c Gene ontology enrichment for the loss of genetic diversity of OWAAM at transplant F3; AMOWA showed no enrichment. Source data are provided as a Source Data file.
To identify life-history traits that could be under selection and reveal potential costs of adaptation, we measured egg production rate across three days of adulthood, survivorship from nauplii to reproductive adult, and combined these metrics with development rate, time to adulthood, and sex ratio to estimate the population net reproductive rate, lambda83, in the first generation after transplant (F1). We found that fecundity was equally high for both lines in their home environments but declined for OWA lines after transplant to AM conditions (48% reduction in egg production relative to OWAOWA; P < 0.02; Fig. 6a). Conversely, transplanted AMOWA were able to maintain similarly high egg production as the non-transplanted lines (Fig. 6a). In contrast to egg production, survivorship from nauplii to reproductive maturity was 60% lower for warming and acidification compared to ambient lineages (P < 0.001; Fig. 6b). Similar to egg production rates, lambda reflected stable population sizes for AM lines in both conditions and OWAOWA, but declined with OWAAM transplant (P = 0.01, Fig. 6c); these estimates were congruent with observed culture densities. The estimated stable growth rate of OWAOWA lines despite decreased survival relative to AM lines (Fig. 6b) was due to compensation in other life-history traits including higher egg production, faster development, and earlier reproductive timing in OWA conditions63, as has been observed in many other taxa including Drosophila84. These results suggest that egg production rate, faster development and earlier time to adulthood may have been life-history traits diverging between ambient and warming and acidification conditions, yielding stable population sizes to compensate for relatively poor survival, as shown with these same lines in our related work63. That poor survival was not observed for AM animals in OWA conditions suggests OWA lines lost plasticity and that there was not a cost to maintaining this plasticity in the constant AM conditions.

For all panels, shape indicates selection line, color indicates environmental condition. Data are presented as mean values and error bars represent 95% confidence intervals. a Egg production rate under ad-libitum and food-limited conditions (n = 6–12 independent mate pairs per food condition per treatment). b Survivorship under ad-libitum and food-limited conditions (n = 3–4 independent experiments per food condition per treatment). c Lambda (net reproductive rate) calculated from combined fecundity, development rate, sex ratio, and survivorship data where a values >1 indicate positive growth (n = 24–28 independent estimates per treatment per food condition). Capital letters in a and c represent statistically separate groups (P < 0.05) from two-way anovas with a post hoc two-sided t-test to correct for multiple testing. Groups with multiple letters are included in multiple statistical groups. Source data are provided as a Source Data file.
The lower survivorship and lambda of the OWA lines relative to AMOWA transplants suggests that evolution in OWA conditions occurred with a trade-off among life-history traits, e.g., survival and age at reproduction85,86 or a trade-off at the physiological level. In experimental evolution studies, lower performance of stress-adapted lines in ancestral environments is common87,88,89, particularly for complex organisms and when stressful conditions are constant90,91,92. The reduced ability of OWA-adapted lines to maintain high fitness across environments could be due to antagonistic pleiotropy, trade-offs at the molecular, cellular or life-history trait levels, via selection for a locus associated with multiple traits, which are adaptive in one environment and mal-adaptive when the environment changes93.
To test the hypothesis that adaptation to a stressful environment could reduce fitness when conditions change to a novel stressor, we reared all evolved and transplanted lines in low-food conditions, a stressor that is predicted to increase in frequency as climate changes64, and measured the same life-history traits at F1. In support of this hypothesis, we found that OWA animals performed poorly under low-food availability regardless of environment (Fig. 6). Under low food, relative to AM lines, OWA lines experienced a 67% reduction in egg production rates (P < 0.001; Fig. 6a), 70% reduction in survival (P < 0.001; Fig. 6b), and a 40% reduction in fitness (P < 0.001; Fig. 6c). These results suggest that selection in warming and acidification conditions may result in populations, which are less resilient to further environmental changes, including expected changes such as a return to ambient conditions or low-food availability94. This reduced flexibility may be driven by increased energetic costs of life in warming and acidification conditions leading to increased stress under low-food conditions95,96 or antagonistic pleiotropy and the costs of adaptation for a complex organism to adapt to a complex stressor92,97, both high pCO2 and high temperature. In the case of increased metabolic costs in OWA conditions, we would not expect genetic divergence between the lines and would see limited differences in plastic transcriptional responses to transplant; the loss of robustness in OWA animals would be due to lower habitat quality and not due to directional selection. Instead, we observe a signal of decreased plasticity driven by selection where there was consistent genomic divergence between OWA and AM lines coupled with a high degree of transcriptional plasticity in AM animals that is not present in OWA animals. This result supports previous findings that long-term evolution in a constant, stressful selective environment can result in specialization, i.e., a loss of plasticity, for example due to antagonistic pleiotropy87,91 or due to high costs of maintaining a generally plastic phenotype18. Alternatively, this rapid loss of plasticity could be driven by neutral mechanisms following the relaxation of selection for plastic phenotypes in a static environment, such as drift or the accumulation of mutations, though the relatively short time frame makes this less likely than selective processes98. Under either mechanism, in the context of global change, such specialization could rapidly occur for populations of a short-lived species experiencing an extreme event.
Combining long-term experimental evolution of a marine metazoan with three generations of reciprocal transplant, we revealed important and complex interactions between adaptation and plasticity in response to global change conditions. Transplanted AM lines experienced much weaker shifts in allele frequency and no targeted loss of adaptive genetic diversity, indicating that, in the short-term (three generations) plasticity largely buffered populations from environmental change allowing AM lines to tolerate warming and acidification conditions. Conversely, the data suggest that over the longer-term (20 generations) transplanted OWA animals required larger shifts in allele frequency and experienced targeted loss of adaptive genetic diversity to survive in what would be predicted to be relaxed selection in a previously benign environment. This indicates that plasticity to acclimate was lost as adaptation to warming and acidification conditions proceeded, indicating costs to maintain plasticity in warming and acidification conditions18. However, these costs appear to be environmentally dependent as plasticity was largely maintained, and therefore less costly, in ambient conditions. While our work here indicates that genetic divergence and selection have shaped gene expression plasticity of OWA animals, future work, such as eQTL studies, would further strengthen this connection.
These results advance our understanding of the relationship between plasticity and adaptive evolution by providing evidence that plasticity did not impede adaptation but, over time, adaptation eroded plasticity. Importantly, by including multiple generations of transplantation following 20 generations of selection, we show that, even with a loss of plasticity, animals can re-evolve to some degree back to their ancestral conditions. However, this re-adaptation carries the cost of losing adaptive genetic variation.
This interplay between plasticity and adaptation has important implications for our understanding of mechanisms of species persistence to global change conditions. We demonstrate that copepods have the capacity to adapt over 20 generations, approximately one year, using the standing genetic variation that exists in natural populations. However, reciprocal transplant and food challenges revealed significant costs of this adaptation; populations lost physiological plasticity, the ability to tolerate food limitation, and ultimately adaptive genetic variation for warming and acidification conditions. Given the increasing frequency of extreme environmental conditions, rapid adaptation of A. tonsa to a temporary extreme may result in a population that is maladapted when conditions return to the previous state. Thus, while plasticity may enable persistence initially, adaptation can drive a loss of plasticity that can lead to reduced resiliency as environmental conditions fluctuate. These results demonstrate the utility of experimental evolution in understanding complex adaptation in natural systems and to reveal the mechanisms, time-scales of responses, consequences, and reversibility of this process. As we begin to incorporate adaptive potential and plasticity into species persistence models99,100,101,102, these results caution that, even for species predicted to be resilient to rapid global change, there may be unforeseen costs to adaptive evolution.
Methods
Experimental set-up
Copepods (n = 1000) were collected in June of 2016 from Esker Point Beach in Groton, CT, USA (decimal degrees: 41.320725, −72.001643) and raised for at least three generations prior to the start of selection to limit maternal effects78. Parental individuals were acclimated to one of two experimental conditions: (1) AM—Ambient temperature (18 ˚C), Ambient CO2 (pH ~8.2; pCO2 ~400 μatm) or (2) OWA—High temperature (22 ˚C), high CO2 (pH ~7.5; pCO2 ~2000 μatm; see Supplementary Table 1 for measured values). Cultures were fed every 48–72 h at food-replete concentrations of carbon (>800 μg C/L) split equally between three species of prey phytoplankton. Prey phytoplankton included Tetraselmis spp., Rhodomonas spp., and Thalassiosira weissflogii, which is a common diet combination used for rearing copepods103. About 400 females and 200 males were used to seed the treatments, yielding an estimated 15,000 eggs and newly hatched nauplii for the initial F0 generation. Four replicate cultures per treatment were maintained in individual 3 L culture containers where each temperature was housed in one of two identical incubators: one incubator for OWA conditions, one for AM conditions; see Dam et al.63 for expanded details. To assess functional life-history traits, smaller volume experiments were housed in the same temperature-controlled incubators in custom plexiglass enclosures with the atmosphere continuously flooded with CO2 infused air at the appropriate concentration, which allows for passive diffusion of CO2 into the experiments. Copepods were raised in the respective two experimental treatments for 20 generations before the reciprocal transplant. To create non-overlapping generations, at each generation, adults were removed after laying eggs to allow for tracking of each generation. Adults were left to lay eggs for 1 week after we observed the first evidence of a new generation (i.e., new nauplii). This allowed nearly all surviving adults to procreate while minimizing the chance that remaining adults would cannibalize or mate with their offspring.
Reciprocal transplant
At the 21st generation, we performed a reciprocal transplant between copepods from AM and OWA. The lineage x environment interaction yielded four unique treatments. Each of the four replicates from each treatment was split to yield four additional replicates for each of two new transplant treatments: AMOWA and OWAAM (as well as sham transfers: AMAM and OWAOWA). Transplant treatments were housed in incubators that corresponded to the new environments along with the original treatments: AMOWA and OWAOWA were housed in the OWA incubator while OWAAM and AMAM cultures were housed in the AM incubator. Copepods were raised for three subsequent generations and were fed every 48–72 h as described above. This design led to 48 total samples for genomics: 2 treatments × 2 transplant/non-transplant × 4 replicates × 3 generations.
Genomics
For each sample, RNA from pools of 20 individuals was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) and purified with Qiagen RNeasy spin columns (Qiagen, Germantown, MD, USA). RNAseq libraries were prepared by Novogene (Sacramento, CA, USA) and sequenced with 150 bp paired end reads on an Illumina NovaSeq 600, generating 1.29 billion reads. Raw reads were trimmed for quality and adapter contamination using Trimmomatic V0.36104 where leading and trailing quality was set at 2, sliding window length was 4 with a quality of 2, and minimum length was 31.
To quantify genetic variation, SuperTranscripts were generated from the reference transcriptome105 using the Trinity_gene_splice_modeler.py command in the Trinity pipeline106. SuperTranscripts act as a reference by collapsing isoforms into a single transcript, which enables variant detection without a reference genome107. Trimmed reads were aligned to the SuperTranscripts using bwa mem108 and duplicate reads were marked with Samblaster v 0.1.22109.
Variants were called using VarScan v2.3110 (two alternate reads to call a variant, P-value of 0.1, minimum coverage of 30x, minimum variant allele frequency of 0.01). Following this, variants were stringently filtered to include no missing data, only sites where at least four samples were called as variable and each with a minor allele frequency of at least 0.025 (minimum of one heterozygous individual), any sites with depth greater than 3x the median (median = 132x), and sites with a minimum coverage of 50x per sample (minimum 2.5 reads/diploid individual; average 174× coverage, average 8.7 reads/diploid individual after filtering; Supplementary Fig. 7). This reduced called variants from an initial 5,547,802 to 322,595.
Gene expression analysis
Gene expression was quantified at the gene level as recommended111 using Salmon v0.10.2112 and transcript abundances were converted to gene-level counts using the tximport package in R113. The reference transcriptome was indexed using quasi mapping with a k-mer length of 31 and transcripts were quantified while correcting for sequence specific bias and GC bias. We were interested in assessing expression patterns between treatments with each generation and how these patterns changed through time and, therefore, analyzed each generation separately. Genes with low expression (fewer than 10 counts in >90% of the samples) were removed from the dataset. This left 23,324, 24,882, and 24,132 genes for F1, F2, and F3, respectively. Counts were normalized and log transformed using the rlog command in DESeq2114.
DESeq2 was used to identify differentially expressed genes. We assume that each line has adapted to their respective environment after 20 generations, as we have demonstrated for these lines previously63. We also assume that the gene expression profiles for each line in their home environment is indicative of an adaptive expression profile for that environment20. Given that we know the populations adapted to their home environments63 and that other studies have found an association between genes that show changes in expression towards the adaptive state and fitness related phenotypes115,116, this assumption is reasonable to make. However, it should be noted that we cannot be positive, which transcriptional changes are directly linked to fitness with the current data and divergence in protein function likely also explains some fitness differences between lines. Differentially expressed genes were identified using the model ~Line + Treatment + Line:Treatment and any contrasts were considered significant when adjusted P-values were <0.05. We identified the plastic genes for each line as the genes changing expression between environments within a line (OWAOWA vs. OWAAM; AMAM vs. AMOWA). We compare the plasticity between AM and OWA lines by plotting the log2 fold change between OWAOWA/OWAAM and AMOWA/AMAM, similar to approaches taken in other plasticity research19,22. To assess the mechanisms driving gene expression divergence between lines in their home environments after 20 generations, we partitioned gene expression changes into Plastic Only and Genetic Change genes82. For genes that were significantly different in expression between OWAOWA and AMAM (F1: 1129 genes; F2: 527 genes; F3: 465 genes), we assessed the mechanisms enabling the transition from AM to OWA (forward evolution) and OWA to AM (reverse). For AM to OWA, when a gene was differentially expressed between the transplanted and home line (AMOWA vs. OWAOWA), the gene expression shift required genetic change. Alternatively, if the gene was significantly plastic (significant AMAM vs. AMOWA) with no genetic change, this was a plasticity only gene. These two categories were mutually exclusive but did not include all genes differentially expressed between AMAM and OWAOWA due to the cases where genes were not differentially expressed between AMAM vs. AMOWA or AMOWA vs. OWAOWA, even though gene expression was different between AMAM vs OWAOWA. We conduct the opposite analysis for OWA to AM. With this analysis we can partition the changes in expression after 20 generations of selection into those that were achieved through plastic vs. evolved mechanisms.
Discriminant analysis of principal components
Discriminant analysis of principal components (DAPC) was used to quantify the degree to which gene expression and allele frequencies converged on the adaptive state within each environment. We know, as above, that lines have adapted to their home environment and made the same assumptions regarding adaptive transcriptional profiles. Using the filtered, normalized, and transformed expression data from DeSeq2, shifts in gene expression in transplanted lines across generations were quantified using DAPC in the adegenet package in R117. Discriminant functions for each generation were first generated using the non-transplanted lines to identify genes consistently differentially expressed. Four principal components that represented 81, 78, and 80% of the variation at F1, F2, and F3 were retained and two clusters that represented the two non-transplanted lines were used. Transplanted lines were fit to this discriminant function space and used MCMCglmm models in R118 to model the effect of line origin and transplant on movement in discriminant function space. In all, 2500 posterior estimates were generated and the difference in the transplant effect for each line was quantified by calculating the absolute difference between these estimates. This can be viewed as the difference between the lengths of the lines representing average shifts in Fig. 4. These differences were used to generate a 95% credible interval and the proportion of positive or negative values were considered a P-value for the difference in magnitude of the effect of transplant on each line in discriminant function space.
DAPC was also used to identify divergence in allele frequency between the AM and OWA lines. The same approach as for gene expression (above) was taken where we generated the DAPC with lines in their home environment and then fit transplanted lines to discriminant function space. Here, we retained three principal components that represented 56, 59, and 63% of the total variation at each generation and again used two clusters.
Adaptive allele frequency and gene expression divergence
To identify loci consistently shifting in response to selection, Cochran–Mantel–Haenszel (CMH) tests were used to identify specific SNPs that were consistently diverged between the AM and OWA lines and represent the most likely targets of adaptation. This approach looks for consistent changes in allele frequencies across all replicates and is a common and powerful technique in experimental evolution26,119. We calculated CMH P-values for AMAM vs. OWAOWA lines (n = 4, each) for each of the three sampled generations. Significance thresholds (P < 5.17e-08) were defined with Bonferroni corrections using the total number of SNPs (322,595) multiplied by three, representing the total number of tests conducted. The SNPs that were identified in all three generations were considered adaptive (Supplementary Fig. 8). This approach assumes that adaptive differences between AMAM and OWAOWA lines after 20 generations are much larger than any additional response to selection during the three generations of transplant. As such, the signals of adaptation in the home environment at F1 should be consistent in F2 and F3 and can be used to identify loci under selection.
DESeq2 was used to identify the specific genes that were adaptively differentially expressed between non-transplanted AM and OWA lines at each generation. Using the same gene set and model from DeSeq2, above, we identified differentially expressed genes between AM and OWA in their home environment with an adjusted significance threshold of P-values < 0.05.
Quantifying genetic diversity
We estimate genetic diversity (π) for each replicate using Popoolation120 with 100 bp non-overlapping sliding windows. To quantify if π was differentially lost in any treatment, median π values were compared using an Anova with a Tukey post hoc test. We next tested if regions containing adaptively divergent loci between non-transplanted OWA and AM lines lost π at a different rate than regions containing only neutral variants. The change in π was calculated for each replicate and the change in π for OWAAM and AMOWA were compared using Wilcoxon Rank Sum tests with bonferroni corrections. Gene ontology enrichment was performed with GO Mann–Whitney U121, which requires no significance threshold, but takes the change in π for each group and asks if any functional category falls towards the tails of this distribution (in our case, one-sided to identify disproportionately low values). We use this same approach for gene expression and allele frequencies divergence between lines in their home environments. For allele frequencies, the minimum P-value was chosen for each gene.
Finally, we tested for overlap between the allelic and gene expression results to determine if the genes in each were significantly correlated. We correlated the relationship between the –log10 of the CMH P-values for the allele frequencies and the differential gene expression P-values between OWAOWA and AMAM. This analysis showed the two sets were distinct and the variation explained by each was less than 1% at each generation, indicating that there was minimal bias in estimating allele frequencies due to differentially expressed genes.
Life-history traits
Day-specific survivorship was measured every 48–72 h, with food media replaced on monitoring days. Food media was provided at the appropriate food concentration (>800 μg C/L for food-replete, 250 μg C/L for food-limited, and 0 μg C/L for starved) divided in equal carbon proportions between the three afore-mentioned prey species in 0.2 μm filtered seawater collected from Long Island Sound. Food media was acclimated to the appropriate temperature and CO2 concentration prior to replacement. Survivorship was assessed among twelve 250-mL beakers per treatment (3 food concentrations × 4 replicates = 12 beakers per treatment) containing 25 individual N1 nauplii in the same plexiglass enclosures as described above and monitored until sexual maturity (adulthood). Life-history results are only presented for food replete and food-limited conditions because no starved individuals survived. Log rank analysis of survivorship was assessed using the survival122,123 and survminer124 packages in R.
Egg production rate (EPR) and hatching success (HS) were assessed with 36 individual mate pairs of newly matured adults per treatment (3 pairs per food concentration × 3 food concentrations × 4 replicates = 36 pairs per treatment). Adults were incubated in 25 mL petri dishes (FisherScientific, Waltham, MA, USA) over three days in the same temperature-controlled incubators and plexiglass enclosures described above. After the initial three-day incubation, adults were assessed for survival and removed to avoid cannibalism of eggs. Eggs were allowed to hatch over a subsequent 3-day incubation. Food media was prepared as described for survivorship and replaced daily during egg laying to ensure accurate food concentrations were near saturation and not reduced due to daily grazing125. Lids of petri dishes were left off-center to allow for full contact with the atmosphere and diffusion of CO2. Plates with dead males were still evaluated for EPR, but not HS. Plates with dead females were not evaluated because we cannot estimate egg production or hatching when no females were alive; mortality was estimated separately and we do not want to confound these two measures. The maximum number of plates excluded was six in the OWAAM treatment at food-limited conditions. No other treatment experienced more than three plates excluded from the assay. After the hatching period, plates were preserved with non-acid Lugol’s solution and eggs counted and nauplii counted. Per capita EPR was calculated as (Eu + Eh)/t where Eu represents unhatched eggs, Eh represents hatched eggs (nauplii), and t represents egg laying time. Hatching success was calculated as Eh/(Eu + Eh).
The population net reproductive rate, λ, was calculated as the dominant eigenvalue of an assembled projected age-structured Leslie Matrix constructed from survivorship and fecundity data83. Briefly, day-specific probabilities of survivorship are calculated from day-specific survivorship as \({P}_{x}=\frac{{{l}}_{x}}{{{l}}_{x-1}}\) where lx represents the number of individuals on day x and lx–1 represents the number of individuals on day x–1. Probabilities of survivorship on day 1 are assumed to be 100%. Per capita EPR and HS are calculated as described above, with fecundity rates equaling the product of EPR and HS. As only females produce offspring, total fecundity rates must be scaled to the sex ratio (ratio of females:males) observed in survivorship experiments. To account for differences in individual development time for each treatment, fecundity rates are assigned to all days after the first matured adult is observed. We assume that survivorship in each beaker is equally as likely to experience the fecundity values observed in EPR experiments. Therefore, each mate-pair fecundity rate was paired with each survivorship beaker to construct a matrix. This yields a maximum of 48 matrices per treatment per food concentration (4 beakers \(\times\) 12 mate pairs). Errors of lambda were calculated as the 95% confidence interval using the formula: \(\underline{x}\pm Z(\frac{SD}{\sqrt{n}})\) where \(\underline{x}\) represents the mean, \(Z\) represents the 95% z-score (1.960), SD represents the standard deviation of the mean, and n represents the number of observations.
We constructed linear mixed models with line, environment, and food concentration as fixed effects with all interactions, and culture replicate as a random effect. The results for net reproductive rate fall within a normal distribution with an inflation of zero values. Thus, we constructed zero-inflated generalized linear mixed models to account for the additional zeroes. Two post hoc tests were conducted. First, to quantify differences in plasticity between the lines, we evaluated genotype \(\times\) environment interactions on life-history traits separately for each food concentration. Second, post hoc t-test comparisons were used to conduct pairwise comparisons of the interactions for each life-history trait.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
The raw sequence data generated in this study have been deposited at NCBI under BioProject PRJNA555881. Life-history data are available as a supplemental file and allele frequency and gene expression table is available on Figshare (https://doi.org/10.6084/m9.figshare.10301690). Source data for all figures are provided with this manuscript.
Code availability
Code to reproduce all analyses are archived on Zenodo: https://doi.org/10.5281/zenodo.5840148126.
References
Rahmstorf, S. & Coumou, D. Increase of extreme events in a warming world. Proc. Natl Acad. Sci. USA 108, 17905–17909 (2011).
Somero, G. N. The physiology of climate change: how potentials for acclimatization and genetic adaptation will determine ‘winners’ and ‘losers’. J. Exp. Biol. 213, 912–920 (2010).
Hoffmann, A. A. & Sgrò, C. M. Climate change and evolutionary adaptation. Nature 470, 479–485 (2011).
Chevin, L.-M., Lande, R. & Mace, G. M. Adaptation, plasticity, and extinction in a changing environment: towards a predictive theory. PLoS Biol. 8, e1000357 (2010).
Kawecki, T. J. & Ebert, D. Conceptual issues in local adaptation. Ecol. Lett. 7, 1225–1241 (2004).
Campbell-Staton, S. C. et al. Winter storms drive rapid phenotypic, regulatory, and genomic shifts in the green anole lizard. Science 357, 495–498 (2017).
Barrett, R. D. H. et al. Linking a mutation to survival in wild mice. Science 363, 499–504 (2019).
Therkildsen, N. O. et al. Contrasting genomic shifts underlie parallel phenotypic evolution in response to fishing. Science 365, 487–490 (2019).
Brennan, R. S., Garrett, A. D., Huber, K. E., Hargarten, H. & Pespeni, M. H. Rare genetic variation and balanced polymorphisms are important for survival in global change conditions. Proc. R. Soc. B: Biol. Sci. 286, 20190943 (2019).
Stearns, S. C. The evolutionary significance of phenotypic plasticity. Bioscience 39, 436–445 (1989).
Thompson, J. D. Phenotypic plasticity as a component of evolutionary change. Trends Ecol. Evol. 6, 246–249 (1991).
Kelly, M. Adaptation to climate change through genetic accommodation and assimilation of plastic phenotypes. Philos. Trans. R. Soc. Lond. B Biol. Sci. 374, 20180176 (2019).
Chevin, L. M., Collins, S. & Lefèvre, F. Phenotypic plasticity and evolutionary demographic responses to climate change: taking theory out to the field. Funct. Ecol. https://doi.org/10.1111/j.1365-2435.2012.02043.x (2013).
Hendry, A. P. Key questions on the role of phenotypic plasticity in eco-evolutionary dynamics. J. Hered. 107, 25–41 (2016).
Calosi, P., De Wit, P., Thor, P. & Dupont, S. Will life find a way? Evolution of marine species under global change. Evol. Appl. 9, 1035–1042 (2016).
Fox, R. J., Donelson, J. M., Schunter, C., Ravasi, T. & Gaitán-Espitia, J. D. Beyond buying time: the role of plasticity in phenotypic adaptation to rapid environmental change. Philos. Trans. R. Soc. Lond. B Biol. Sci. 374, 20180174 (2019).
Lande, R. Adaptation to an extraordinary environment by evolution of phenotypic plasticity and genetic assimilation. J. Evol. Biol. 22, 1435–1446 (2009).
Murren, C. J. et al. Constraints on the evolution of phenotypic plasticity: limits and costs of phenotype and plasticity. Heredity 115, 293–301 (2015).
Posavi, M., Gulisija, D., Munro, J. B., Silva, J. C. & Lee, C. E. Rapid evolution of genome-wide gene expression and plasticity during saline to freshwater invasions by the copepod Eurytemora affinis species complex. Mol. Ecol. 29, 4835–4856 (2020).
Ghalambor, C. K. et al. Non-adaptive plasticity potentiates rapid adaptive evolution of gene expression in nature. Nature 525, 372–375 (2015).
Kelly, M. W., Pankey, M. S., DeBiasse, M. B. & Plachetzki, D. C. Adaptation to heat stress reduces phenotypic and transcriptional plasticity in a marine copepod. Funct. Ecol. 31, 398–406 (2017).
Sikkink, K. L., Reynolds, R. M., Ituarte, C. M., Cresko, W. A. & Phillips, P. C. Rapid evolution of phenotypic plasticity and shifting thresholds of genetic assimilation in the nematode Caenorhabditis remanei. G3 4, 1103–1112 (2014).
Brennan, R. S., Galvez, F. & Whitehead, A. Reciprocal osmotic challenges reveal mechanisms of divergence in phenotypic plasticity in the killifish Fundulus heteroclitus. J. Exp. Biol. 218, 1212–1222 (2015).
Kelly, M. W., Pankey, M. S. & DeBiasse, M. B. Adaptation to heat stress reduces phenotypic and transcriptional plasticity in a marine copepod. Funct. Ecol. https://doi.org/10.1111/1365-2435.12725 (2017).
Waddington, C. H. Genetic assimilation of an acquired character. Evolution 7, 118–126 (1953).
Schlötterer, C., Kofler, R., Versace, E., Tobler, R. & Franssen, S. U. Combining experimental evolution with next-generation sequencing: a powerful tool to study adaptation from standing genetic variation. Heredity 114, 431–440 (2015).
Munday, P. L., Warner, R. R., Monro, K., Pandolfi, J. M. & Marshall, D. J. Predicting evolutionary responses to climate change in the sea. Ecol. Lett. 16, 1488–1500 (2013).
Huang, Y. & Agrawal, A. F. Experimental evolution of gene expression and plasticity in alternative selective regimes. PLoS Genet. 12, e1006336 (2016).
Mallard, F., Nolte, V. & Schlötterer, C. The evolution of phenotypic plasticity in response to temperature stress. Genome Biol. Evol. 12, 2429–2440 (2020).
Schaum, C. E. & Collins, S. Plasticity predicts evolution in a marine alga. Proc. Biol. Sci. 281, 20141486 (2014).
Kelly, S. A., Czech, P. P., Wight, J. T., Blank, K. M. & Garland, T. Jr Experimental evolution and phenotypic plasticity of hindlimb bones in high-activity house mice. J. Morphol. 267, 360–374 (2006).
Garland, T. Jr & Kelly, S. A. Phenotypic plasticity and experimental evolution. J. Exp. Biol. 209, 2344–2361 (2006).
Gibbin, E. M., Massamba N’Siala, G., Chakravarti, L. J., Jarrold, M. D. & Calosi, P. The evolution of phenotypic plasticity under global change. Sci. Rep. 7, 17253 (2017).
McCairns, R. J. S. & Bernatchez, L. Adaptive divergence between freshwater and marine sticklebacks: insights into the role of phenotypic plasticity from an integrated analysis of candidate gene expression. Evolution 64, 1029–1047 (2010).
Whitehead, A. The evolutionary radiation of diverse osmotolerant physiologies in killifish (Fundulus sp.). Evolution 64, 2070–2085 (2010).
Lind, M. I. & Johansson, F. The degree of adaptive phenotypic plasticity is correlated with the spatial environmental heterogeneity experienced by island populations of Rana temporaria. J. Evol. Biol. 20, 1288–1297 (2007).
Lázaro-Nogal, A. et al. Environmental heterogeneity leads to higher plasticity in dry-edge populations of a semi-arid Chilean shrub: insights into climate change responses. J. Ecol. 103, 338–350 (2015).
Gianoli, E. Plasticity of traits and correlations in two populations of Convolvulus arvensis (Convolvulaceae) differing in environmental heterogeneity. Int. J. Plant Sci. 165, 825–832 (2004).
Fischer, E. K., Song, Y., Hughes, K. A., Zhou, W. & Hoke, K. L. Nonparallel transcriptional divergence during parallel adaptation. Mol. Ecol. 30, 1516–1530 (2021).
Gunter, H. M., Schneider, R. F., Karner, I., Sturmbauer, C. & Meyer, A. Molecular investigation of genetic assimilation during the rapid adaptive radiations of East African cichlid fishes. Mol. Ecol. 26, 6634–6653 (2017).
Bitter, M. C. et al. Fluctuating selection and global change: a synthesis and review on disentangling the roles of climate amplitude, predictability and novelty. Proc. Biol. Sci. 288, 20210727 (2021).
Skliris, N. et al. Salinity changes in the World Ocean since 1950 in relation to changing surface freshwater fluxes. Clim. Dyn. 43, 709–736 (2014).
Collins, M. et al. Long-term climate change: projections, commitments and irreversibility. in Climate Change 2013-The Physical Science Basis: Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. 1029–1136 (Cambridge University Press, 2013).
Sunday, J. M. et al. Evolution in an acidifying ocean. Trends Ecol. Evol. 29, 117–125 (2014).
Reusch, T. B. H. & Boyd, P. W. Experimental evolution meets marine phytoplankton. Evolution 67, 1849–1859 (2013).
Palumbi, S. R., Evans, T. G., Pespeni, M. H. & Somero, G. N. Present and future adaptation of marine species assemblages. Oceanography https://doi.org/10.5670/oceanog.2019.314 (2019).
Helmuth, B. et al. Long-term, high frequency in situ measurements of intertidal mussel bed temperatures using biomimetic sensors. Sci. Data 3, 160087 (2016).
Feely, R. A., Sabine, C. L., Hernandez-Ayon, J. M., Ianson, D. & Hales, B. Evidence for upwelling of corrosive ‘acidified’ water onto the continental shelf. Science 320, 1490–1492 (2008).
Cowen, R. K. & Sponaugle, S. Larval dispersal and marine population connectivity. Ann. Rev. Mar. Sci. 1, 443–466 (2009).
Huys, R. & Boxshall, G. A. Copepod Evolution. (marinespecies.org, 1991).
Langer, J. A. F. et al. Acclimation and adaptation of the coastal calanoid copepod Acartia tonsa to ocean acidification: a long-term laboratory investigation. Mar. Ecol. Prog. Ser. 619, 35–51 (2019).
Dam, H. G. Evolutionary adaptation of marine zooplankton to global change. Ann. Rev. Mar. Sci. 5, 349–370 (2013).
De Wit, P., Dupont, S. & Thor, P. Selection on oxidative phosphorylation and ribosomal structure as a multigenerational response to ocean acidification in the common copepod Pseudocalanus acuspes. Evol. Appl. 9, 1112–1123 (2016).
Thor, P. & Dupont, S. Transgenerational effects alleviate severe fecundity loss during ocean acidification in a ubiquitous planktonic copepod. Glob. Chang. Biol. 21, 2261–2271 (2015).
Donelson, J. M. et al. Understanding interactions between plasticity, adaptation and range shifts in response to marine environmental change. Philos. Trans. R. Soc. Lond. B Biol. Sci. 374, 20180186 (2019).
Gibbin, E. M. et al. Can multi-generational exposure to ocean warming and acidification lead to the adaptation of life history and physiology in a marine metazoan? J. Exp. Biol. 220, 551–563 (2017).
Mauchline, J. The Biology of Calanoid Copepods (Academic Press, 1998).
Steinberg, D. K. & Landry, M. R. Zooplankton and the ocean carbon cycle. Ann. Rev. Mar. Sci. 9, 413–444 (2017).
Gobler, C. J. & Baumann, H. Hypoxia and acidification in ocean ecosystems: coupled dynamics and effects on marine life. Biol. Lett. 12, 20150976 (2016).
Rice, E., Dam, H. G. & Stewart, G. Impact of climate change on estuarine zooplankton: surface water warming in Long Island Sound is associated with changes in copepod size and community structure. Estuaries Coasts 38, 13–23 (2015).
IPCC. Climate Change 2014: Mitigation of Climate Change. Contribution of Working Group III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Vol. 1454 (IPCC, 2014).
Caldeira, K. & Wickett, M. E. Oceanography: anthropogenic carbon and ocean pH. Nature 425, 365 (2003).
Dam, H. G. et al. Rapid, but limited, zooplankton adaptation to simultaneous warming and acidification. Nat. Clim. Chang. 11, 780–786 (2021).
Behrenfeld, M. J. et al. Climate-driven trends in contemporary ocean productivity. Nature 444, 752–755 (2006).
Barghi, N., Hermisson, J. & Schlötterer, C. Polygenic adaptation: a unifying framework to understand positive selection. Nat. Rev. Genet. 21, 769–781 (2020).
Láruson, Á. J., Yeaman, S. & Lotterhos, K. E. The importance of genetic redundancy in evolution. Trends Ecol. Evol. 35, 809–822 (2020).
Tobler, R. et al. Massive habitat-specific genomic response in D. melanogaster populations during experimental evolution in hot and cold environments. Mol. Biol. Evol. 31, 364–375 (2014).
Belhadj Slimen, I. et al. Reactive oxygen species, heat stress and oxidative-induced mitochondrial damage. A review. Int. J. Hyperth. 30, 513–523 (2014).
Downs, C. A. & Heckathorn, S. A. The mitochondrial small heat-shock protein protects NADH:ubiquinone oxidoreductase of the electron transport chain during heat stress in plants. FEBS Lett. 430, 246–250 (1998).
Harada, A. E., Healy, T. M. & Burton, R. S. Variation in thermal tolerance and its relationship to mitochondrial function across populations of Tigriopus californicus. Front. Physiol. 10, 213 (2019).
Chung, D. J. & Schulte, P. M. Mitochondria and the thermal limits of ectotherms. J. Exp. Biol. 223 (2020).
Mathew, A. N. U. & Morimoto, R. I. Role of the heat-shock response in the life and death of proteins. Ann. N. Y. Acad. Sci. 851, 99–111 (1998).
Evans, T. G., Pespeni, M. H., Hofmann, G. E., Palumbi, S. R. & Sanford, E. Transcriptomic responses to seawater acidification among sea urchin populations inhabiting a natural pH mosaic. Mol. Ecol. 26, 2257–2275 (2017).
Bailey, A. et al. Regulation of gene expression is associated with tolerance of the Arctic copepod Calanus glacialis to CO2-acidified sea water. Ecol. Evol. 7, 7145–7160 (2017).
Tenaillon, O. et al. The molecular diversity of adaptive convergence. Science 335, 457–461 (2012).
Anjum, R. & Blenis, J. The RSK family of kinases: emerging roles in cellular signalling. Nat. Rev. Mol. Cell Biol. 9, 747–758 (2008).
Marshall, D. J. Transgenerational plasticity in the sea: context-dependent maternal effects across the life history. Ecology 89, 418–427 (2008).
Vehmaa, A., Brutemark, A. & Engström-Öst, J. Maternal effects may act as an adaptation mechanism for copepods facing pH and temperature changes. PLoS ONE 7, e48538 (2012).
Skinner, M. K. What is an epigenetic transgenerational phenotype? F3 or F2. Reprod. Toxicol. 25, 2–6 (2008).
Sasaki, M. C. & Dam, H. G. Integrating patterns of thermal tolerance and phenotypic plasticity with population genetics to improve understanding of vulnerability to warming in a widespread copepod. Glob. Chang. Biol. 25, 4147–4164 (2019).
Sasaki, M. C. & Dam, H. G. Genetic differentiation underlies seasonal variation in thermal tolerance, body size, and plasticity in a short‐lived copepod. Ecol. Evol. 90, 193 (2020).
Ho, W.-C., Li, D., Zhu, Q. & Zhang, J. Phenotypic plasticity as a long-term memory easing readaptations to ancestral environments. Sci. Adv. 6, eaba3388 (2020).
Caswell, H. Matrix population models. Encyclopedia of Environmetrics 3, https://doi.org/10.1002/9781118445112.stat07481 (2006).
Huey, R. B., Wakefield, T., Crill, W. D. & Gilchrist, G. W. Within- and between-generation effects of temperature on early fecundity of Drosophila melanogaster. Heredity 74, 216–223 (1995). Pt 2.
Zwaan, B., Bijlsma, R. & Hoekstra, R. F. Direct selection on life span in Drosophila melanogaster. Evolution 49, 649–659 (1995).
Reznick, D. A., Bryga, H. & Endler, J. A. Experimentally induced life-history evolution in a natural population. Nature 346, 357–359 (1990).
Jerison, E. R., Nguyen Ba, A. N., Desai, M. M. & Kryazhimskiy, S. Chance and necessity in the pleiotropic consequences of adaptation for budding yeast. Nat. Ecol. Evol. 4, 601–611 (2020).
Zhong, S., Khodursky, A., Dykhuizen, D. E. & Dean, A. M. Evolutionary genomics of ecological specialization. Proc. Natl Acad. Sci. USA 101, 11719–11724 (2004).
MacLean, R. C., Bell, G. & Rainey, P. B. The evolution of a pleiotropic fitness tradeoff in Pseudomonas fluorescens. Proc. Natl Acad. Sci. USA 101, 8072–8077 (2004).
Bettencourt, B. R., Feder, M. E. & Cavicchi, S. Experimental evolution of HSP70 expression and thermotolerance in Drosophila melanogaster. Evolution 53, 484–492 (1999).
Schaum, C.-E., Buckling, A., Smirnoff, N., Studholme, D. J. & Yvon-Durocher, G. Environmental fluctuations accelerate molecular evolution of thermal tolerance in a marine diatom. Nat. Commun. 9, 1719 (2018).
Orr, H. A. Adaptation and the cost of complexity. Evolution 54, 13–20 (2000).
Chen, P. & Zhang, J. Antagonistic pleiotropy conceals molecular adaptations in changing environments. Nat. Ecol. Evol. 4, 461–469 (2020).
Cox, P. M., Betts, R. A., Jones, C. D., Spall, S. A. & Totterdell, I. J. Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model. Nature 408, 184–187 (2000).
Mayor, D. J., Sommer, U., Cook, K. B. & Viant, M. R. The metabolic response of marine copepods to environmental warming and ocean acidification in the absence of food. Sci. Rep. 5, 13690 (2015).
Pedersen, S. A. et al. Multigenerational exposure to ocean acidification during food limitation reveals consequences for copepod scope for growth and vital rates. Environ. Sci. Technol. 48, 12275–12284 (2014).
Bono, L. M., Smith, L. B. Jr, Pfennig, D. W. & Burch, C. L. The emergence of performance trade-offs during local adaptation: insights from experimental evolution. Mol. Ecol. 26, 1720–1733 (2017).
Masel, J., King, O. D. & Maughan, H. The loss of adaptive plasticity during long periods of environmental stasis. Am. Nat. 169, 38–46 (2007).
Bay, R. A. et al. Genomic signals of selection predict climate-driven population declines in a migratory bird. Science 359, 83–86 (2018).
Bay, R. A. et al. Predicting responses to contemporary environmental change using evolutionary response architectures. Am. Nat. 189, 463–473 (2017).
Bush, A. et al. Incorporating evolutionary adaptation in species distribution modelling reduces projected vulnerability to climate change. Ecol. Lett. 19, 1468–1478 (2016).
Valladares, F. et al. The effects of phenotypic plasticity and local adaptation on forecasts of species range shifts under climate change. Ecol. Lett. 17, 1351–1364 (2014).
Feinberg, L. R. & Dam, H. G. Effects of diet on dimensions, density and sinking rates of fecal pellets of the copepod Acartia tonsa. Mar. Ecol. Prog. Ser. 175, 87–96 (1998).
Bolger, A. M., Lohse, M. & Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120 (2014).
Jørgensen, T. S. et al. The genome and mRNA transcriptome of the cosmopolitan calanoid copepod Acartia tonsa Dana improve the understanding of copepod genome size evolution. Genome Biol. Evol. https://doi.org/10.1093/gbe/evz067 (2019).
Haas, B. J. et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 8, 1494–1512 (2013).
Davidson, N. M., Hawkins, A. D. K. & Oshlack, A. SuperTranscripts: a data driven reference for analysis and visualisation of transcriptomes. Genome Biol. 18, 148 (2017).
Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. Preprint at https://arxiv.org/abs/1303.3997 (2013).
Faust, G. G. & Hall, I. M. SAMBLASTER: fast duplicate marking and structural variant read extraction. Bioinformatics 30, 2503–2505 (2014).
Koboldt, D. C. et al. VarScan 2: somatic mutation and copy number alteration discovery in cancer by exome sequencing. Genome Res. 22, 568–576 (2012).
Soneson, C., Love, M. I. & Robinson, M. D. Differential analyses for RNA-seq: transcript-level estimates improve gene-level inferences. F1000Res. 4, 1521 (2016).
Patro, R., Duggal, G., Love, M. I., Irizarry, R. A. & Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 14, 417–419 (2017).
R Core Team. R: A Language and Environment for Statistical Computing (R Core Team, 2019).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Kenkel, C. D. & Matz, M. V. Gene expression plasticity as a mechanism of coral adaptation to a variable environment. Nat. Ecol. Evol. 1, 14 (2016).
Campbell-Staton, S. C., Velotta, J. P. & Winchell, K. M. Selection on adaptive and maladaptive gene expression plasticity during thermal adaptation to urban heat islands. Nat. Commun. 12, 6195 (2021).
Jombart, T. & Ahmed, I. adegenet 1.3-1: new tools for the analysis of genome-wide SNP data. Bioinformatics https://doi.org/10.1093/bioinformatics/btr521 (2011).
Hadfield, J. D. MCMC methods for multi-response generalized linear mixed models: The MCMCglmm R Package. J. Stat. Softw. 33, 1–22 (2010).
Orozco-terWengel, P. et al. Adaptation of Drosophila to a novel laboratory environment reveals temporally heterogeneous trajectories of selected alleles. Mol. Ecol. 21, 4931–4941 (2012).
Kofler, R. et al. PoPoolation: a toolbox for population genetic analysis of next generation sequencing data from pooled individuals. PLoS ONE 6, e15925 (2011).
Wright, R. M., Aglyamova, G. V., Meyer, E. & Matz, M. V. Gene expression associated with white syndromes in a reef building coral, Acropora hyacinthus. BMC Genomics 16, 371 (2015).
Therneau, T. M. & Grambsch, P. M. Modeling Survival Data: Extending the Cox Model (Springer, 2013).
Therneau, T. A Package for Survival Analysis in S. version 2.38. (Mayo Foundation, 2015).
Kassambara, A., Kosinski, M., Biecek, P. & Fabian, S. Package ‘survminer’. Drawing Survival Curves using ‘ggplot2’. (R package version 0. 3. 1.) (2017).
Houde, S. E. L. & Roman, M. R. Effects of food quality on the functional ingestion response of the copepod Acartia tonsa. Mar. Ecol. Prog. Ser. 40, 69–77 (1987).
Brennan, R. S. et al. Code repository for ‘Loss of transcriptional plasticity but sustained adaptive capacity after adaptation to global change conditions in a marine copepod’. Zenodo https://doi.org/10.5281/zenodo.5840148 (2022).
Acknowledgements
We thank Lydia Norton and Gihong Park for culture maintenance and experimental help. RSB appreciates the sponsorship from Maile Neel as a Visiting Research Scientist in the Department of Plant Science & Landscape Architecture at the University of Maryland College Park. Funding: This work was funded by the National Science Foundation grants to OCE 1559075 (M.H.P.) and OCE 1559180 (H.G.D., H.B., and M.F.) as well as Connecticut Sea Grant R/LR‐25 (H.G.D., M.F., and H.B.).
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
Conceptualization and experimental design: R.S.B., J.A.d., H.G.D., M.F., H.B., M.H.P.; Experimental execution: J.A.d.; Data collection and analysis: R.S.B. and J.A.d.; Writing—original draft: R.S.B. and M.H.P.; Writing-review and editing: R.S.B., J.A.d., H.G.D., M.F., H.B., M.H.P.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Pierre De Wit and the other anonymous reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Brennan, R.S., deMayo, J.A., Dam, H.G. et al. Loss of transcriptional plasticity but sustained adaptive capacity after adaptation to global change conditions in a marine copepod. Nat Commun 13, 1147 (2022). https://doi.org/10.1038/s41467-022-28742-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-022-28742-6
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
