
Abstract
Due to their potential impact on ecosystems and biogeochemistry, microbial interactions, such as those between phytoplankton and bacteria, have been studied intensively using specific model organisms. Yet, to what extent interactions differ between closely related organisms, or how these interactions change over time, or culture conditions, remains unclear. Here, we characterize the interactions between five strains each of two globally abundant marine microorganisms, Prochlorococcus (phototroph) and Alteromonas (heterotroph), from the first encounter between individual strains and over more than a year of repeated cycles of exponential growth and long-term nitrogen starvation. Prochlorococcus-Alteromonas interactions had little effect on traditional growth parameters such as Prochlorococcus growth rate, maximal fluorescence, or lag phase, affecting primarily the dynamics of culture decline, which we interpret as representing cell mortality and lysis. The shape of the Prochlorococcus decline curve and the carrying capacity of the co-cultures were determined by the phototroph and not the heterotroph strains involved. Comparing various mathematical models of culture mortality suggests that Prochlorococcus death rate increases over time in mono-cultures but decreases in co-cultures, with cells potentially becoming more resistant to stress. Our results demonstrate intra-species differences in ecologically relevant co-culture outcomes. These include the recycling efficiency of N and whether the interactions are mutually synergistic or competitive. They also highlight the information-rich growth and death curves as a useful readout of the interaction phenotype.
Similar content being viewed by others



Introduction
Interactions among microorganisms occur in every known ecosystem (recently reviewed by [1, 2]). Detailed studies of the interactions between selected model organisms (often in laboratory co-cultures) have begun to reveal the diversity of molecular mechanisms whereby organisms interact with each other [2,3,4]. However, it is currently unknown to what extent the studied interactions differ between organism pairs, growth stages, or environmental conditions. For example, while broad-scale phylogenetic patterns are often observed in microbial interactions, closely related bacteria may differ in the way they interact with other organisms, likely as a result of the significant genetic diversity observed in many microbial clades (e.g. [5, 6]). Additionally, the same pair of interacting organisms can synergize or compete depending on the composition of the culture media and the growth stage of (co)-culture (e.g. [7,8,9]). Finally, both the coarse-grained ecological classification of microbial interactions (e.g. positive/negative) and the high-resolution mechanistic view obtained using advanced physiology and ‘omics approaches are difficult to translate into quantitative, predictive models of organismal growth and decline [1, 10, 11].
Here we explore to what extent intra-clade diversity affects the outcome of microbial interactions, using growth curves as an information-rich view of microbial growth and mortality. Growth curves can be divided into discrete phases (lag, exponential, stationary, decline, and long-term stationary phases), and can be used to extract quantitative parameters such as growth rates and lag times [12, 13]. An extra layer of more subtle information may exist in the shapes of the growth curves, providing hints of important shifts in the physiology of the growing organisms, as classically demonstrated by Jacques Monod for diauxic growth in Escherichia coli [14]. While many studies of bacterial interactions focus on the exponential growth stage or on culture yield at a specific time-point (e.g. [15,16,17]), fewer studies look at the shape and dynamics of the decline phases, which can provide important hints regarding the effect of interactions on the process of microbial mortality (e.g. [18,19,20]).
Our model organisms are two globally abundant clades of marine bacteria: a cyanobacterial primary producer (Prochlorococcus) and a heterotrophic γ-proteobacterium (Alteromonas). Interactions between marine phototrophs (phytoplankton, including cyanobacteria) and heterotrophic bacteria have been studied intensively, as phytoplankton are responsible for about one-half of the photosynthesis on Earth (e.g. [21,22,23,24,25]). Thus, phytoplankton-bacteria interactions may strongly affect community structure and function on scales from microns to thousands of kilometers [26, 27]. Our model primary producer, Prochlorococcus, is found throughout the euphotic zone, the sunlit upper portion, of the oligotrophic (nutrient-poor) ocean. There are multiple Prochlorococcus clades, broadly partitioned into high-light (HL) and low-light (LL) adapted ecotypes, which differ in their photosynthetic parameters and occupy different niches in the ocean (e.g. surface verses deep water, reviewed by [28]). Strains differ also in traits such as the capacity to utilize different forms of inorganic nutrients and organic matter, as well as in their interactions with heterotrophic bacteria and phage. Alteromonas is a clade of free-living marine bacteria, which are also partitioned into surface and deep groups (A. macleodii and A. mediterranea, respectively) [29]. Alteromonas strains also exhibit diverse capabilities to utilize carbohydrates, to acquire iron, and in motility [30]. Interactions between individual strains of Prochlorococcus and Alteromonas have been characterized in some detail [12, 27, 31,32,33,34,35]. While the phenotype and gene expression patterns during interactions vary between strains, this variability has not been explored systematically ([12, 32, 36] and Supplementary Text S1). Strain- and condition-dependent phytoplankton-heterotroph interactions are observed also in other systems, including Synechococcus, a close relative of Prochlorococcus [18, 37, 38], as well as eukaryotic microalgae (e.g. coccolithophores and diatoms, [7,8,9, 39]).
We characterized the interactions between five strains each of Prochlorococcus and Alteromonas, from the first encounter between previously axenic strains (i.e., grown in mono-culture) and across ~1.2 years of co-culture (25 phototroph-heterotroph combinations). The culturing period spanned multiple cycles of exponential growth, culture decline and long-term nitrogen starvation [33]. Nitrogen limitation occurs across wide swaths of the global ocean, and affects a substantial proportion of Prochlorococcus diversity [40, 41]. We focused our analysis on Prochlorococcus growth and decline. Using this dataset of 429 growth curves, as well as associated cell counts, we ask: (i) How do the interactions between Prochlorococcus and Alteromonas vary across the diversity of each organisms? (ii) Do the interactions change over time (i.e. do the organisms adapt to “living together”)? (iii) When, during the life-cycle of a Prochlorococcus batch culture, do microbial interactions have the largest impact on growth, death, and overall culture carrying capacity, and can this impact be quantified?
Results and discussion
All Alteromonas strains support long-term survival of Prochlorococcus under N starvation
Previous research showed that Prochlorococcus, and to some extent Synechococcus depend on co-occurring heterotrophic bacteria to survive various types of stress, including nitrogen starvation [33, 34, 42, 43]. At the first encounter between previously axenic Prochlorococcus and Alteromonas (E1), all co-cultures and axenic controls grew exponentially (Fig. 1B, C). However, all axenic cultures showed a rapid and mostly monotonic decrease in fluorescence starting shortly after the cultures stopped growing, reaching levels below the limit of detection after ~20–30 days. None of the axenic Prochlorococcus cultures were able to re-grow when transferred into fresh media after 60 days (Fig. 1C). In contrast, the decline of co-cultures rapidly slowed, and in some cases was interrupted by an extended “plateau” or second growth stage (Fig. 1B). Across multiple experiments, 92% of the co-cultures contained living Prochlorococcus cells for at least 140 days, meaning that they could be revived by transfer into fresh media. Thus, the ability of Alteromonas to support long-term N starvation in Prochlorococcus was conserved in all analyzed strains.
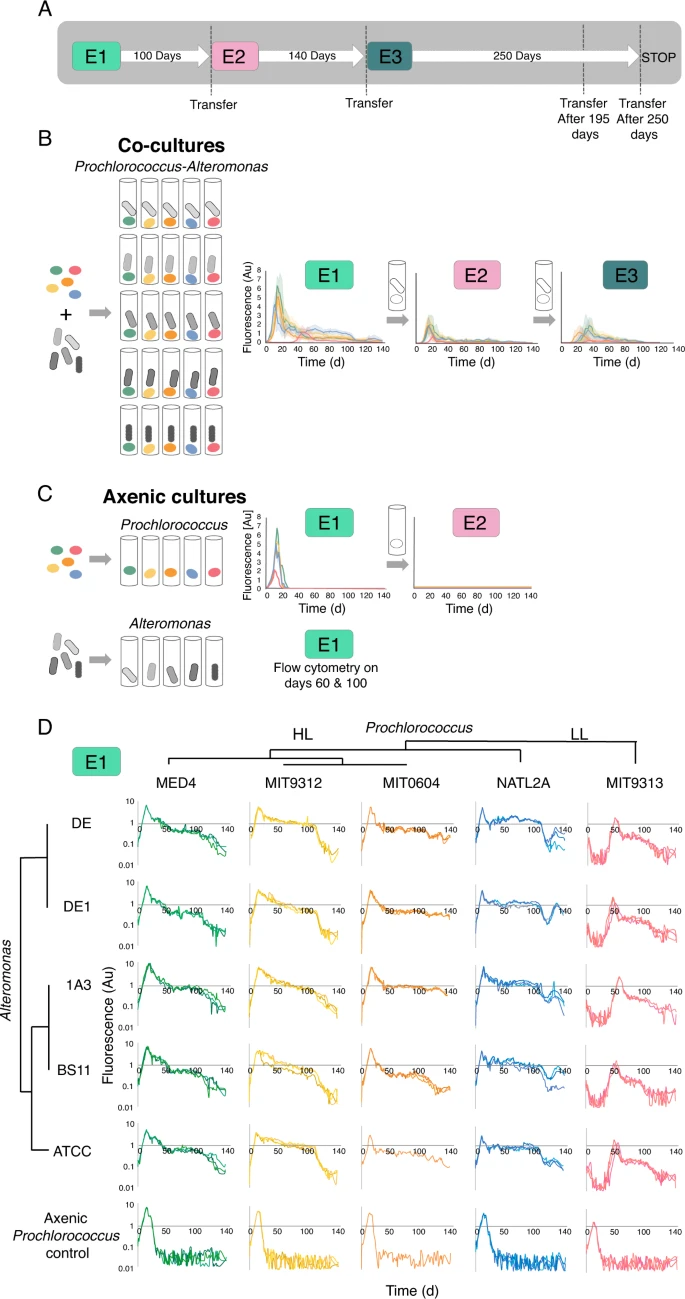
A Schematic illustration of the experimental design. One ml from Experiment E1 was transferred into 20 ml fresh media after 100 days, starting experiment E2. Experiment E2 was similarly transferred into fresh media after 140 days, starting experiment E3. Additional experiments replicating these transfers are described in Supplementary Fig. S1. B Overview of the growth curves of the 25 Prochlorococcus-Alteromonas co-cultures over three transfers spanning ~1.2 years (E1, E2 and E3). Results show mean + standard error from biological triplicates, colored by Prochlorococcus strain as in panel D. C Axenic Prochlorococcus grew exponentially in E1 but failed to grow when transferred into fresh media after 60, 100, or 140 days. Axenic Alteromonas cultures were counted after 60 and 100 days, as their growth cannot be monitored sensitively and non-invasively using fluorescence (optical density is low at these cell numbers). D High reproducibility and strain-specific dynamics of the initial contact between Prochlorococcus and Alteromonas strains (E1). Three biological replicates for each mono-culture and co-culture are shown. Note that the Y axis is linear in panels B, C and logarithmic in panel D. Au: arbitrary units.
It has previously been shown that Prochlorococcus MIT9313 is initially inhibited by co-culture with Alteromonas HOT1A3, while Prochlorococcus MED4 is not [12, 32]. This “delayed growth” phenotype was observed here too, was specific to MIT9313 (not observed in other Prochlorococcus strains) and occurred with all Alteromonas strains tested (Fig. 1D). MIT9313 belongs to the low-light adapted clade IV (LLIV), which are relatively distant from other Prochlorococcus strains and differ from them in multiple physiological aspects including the structure of their cell wall [44], the use of different (and nitrogen-containing) compatible solutes [45], and the production of multiple peptide secondary metabolites (lanthipeptides, [46, 47]). LLIV cells also have larger genomes, and are predicted to take up a higher diversity of organic compounds such as sugars and amino acids [48]. It is intriguing that specifically this strain, which has higher predicted metabolic and regulatory flexibilities [49], is the only one initially inhibited in co-culture with Alteromonas.
Differences in co-culture phenotype are related to Prochlorococcus and not Alteromonas strains and occur primarily during the decline stage
While co-culture with all Alteromonas strains had a major effect on Prochlorococcus viability after long-term starvation, there was no significant effect of co-culture on traditional metrics of growth such as maximal growth rate, maximal fluorescence, and lag phase (with the exception of the previously described inhibition of MIT9313; Fig. 2A–C). However, a visual inspection of the growth curves suggested subtle yet consistent differences in the shape of the growth curve, and especially the decline phase, between the different Prochlorococcus strains in the co-cultures (Fig. 1D). To test this, we used the growth curves as input for a principal component analysis (PCA), revealing that the growth curves from each Prochlorococcus strain clustered together, regardless of which Alteromonas strain they were co-cultured with (Fig. 2D). The growth curves of all high-light adapted strains (MED4, MIT9312, and MIT0604) were relatively similar, the low-light I strain NATL2A was somewhat distinct, and the low-light IV strain MIT9313 was a clear outlier (Fig. 2D), consistent with this strain being the only one initially inhibited in all co-cultures. Random forest classification supported the observation that the growth curve shapes were affected more by the Prochlorococcus rather than Alteromonas strains, and also confirmed the visual observation that most of the features differentiating between Prochlorococcus strains occurred during culture decline (random forest is a supervised machine learning algorithm explained in more detail in Supplementary Text S2; see also Supplementary Fig. S2). Thus, co-culture with Alteromonas affects the decline stage of Prochlorococcus in co-culture in a way that differs between Prochlorococcus but not Alteromonas strains.
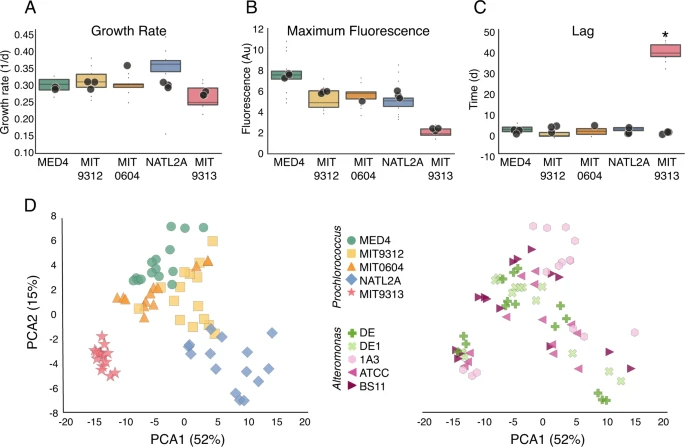
A Growth rate, B Maximum fluorescence, and C duration of lag phase during experiment E1. Box-plots represent mean and 75th percentile of co-cultures, circles represent measurements of individual cultures of the axenic controls. The only significant difference between axenic and co-cultures is in the length of the lag phase for MIT9313 (Bonferroni corrected ANOVA, p < 0.001). D PCA ordination of the growth curves colored by Prochlorococcus (left) and by Alteromonas (right) strains. The growth curves cluster by Prochlorococcus strain (Adonis test, F(4,68) = 42.3, R2 = 0.71, p = 0.001) and only marginally by Alteromonas strain (Adonis test, F(4,68) = 2.29, R2 = 0.11, p = 0.017). Au arbitrary units.
We next asked whether the phenotypes of interaction, which were observed when high cell densities of Prochlorococcus and Alteromonas interacted for the first time (E1), were maintained after the cells had lived together in co-culture for extended periods. We therefore continued to transfer the co-cultures into fresh media over multiple additional transfers, performed 40–200 days after the initial inoculations. In total, fluorescence measurements are available for a cumulative period of 380 days, which the cells spent in co-culture (Fig. 1A; Supplementary Fig. S1). The ability of Prochlorococcus to survive long-term N starvation, the clustering of the growth curves by Prochlorococcus but not Alteromonas strains, and the results of the random forest classification, were all reproduced in subsequent transfers (Fig. 1B; Supplementary Figs. S2, S3; Supplementary Text S3). These observations are thus robust to the cumulative time the organisms have been interacting and the cell densities of both organisms when transferred to new media (see below).
Differences in the carrying capacity suggest different modes of interaction
While Alteromonas clearly support Prochlorococcus, by enabling it to survive long-term N starvation, is the reciprocal interaction also synergistic? Do Prochlorococcus enhance the growth of Alteromonas, and does the interaction affect the overall carrying capacity of the system, defined here as the ability to efficiently utilize the limiting resource (nitrogen)? To answer these questions, we used the flow cytometry cell counts of Prochlorococcus and Alteromonas on days 60, 100, and 140 of experiment E1 to infer the nitrogen (N) biomass of each population grown alone or in co-culture (Fig. 3; Supplementary Table S1; see Methods and Supplementary Text S4 for the calculations and caveats).

A Growth curves of experiment E1, arrows showing the days where cell numbers were counted by flow cytometry (see also Supplementary Table S1). Axenic curves shown are from all Prochlorococcus strains. Thick lines represent mean fluorescence. B Total calculated N biomass of the cultures in E1 on days 60 and 100 in μmol N/L. Dashed line indicates total available N in co-cultures. C Log2 Fold Change (log2FC) of Alteromonas N biomass relative to axenic Alteromonas controls on days 60 and 100. Asterisks indicate statistically significant FC (Bonferroni corrected ANOVA p < 0.05). ALT Alteromonas, PRO Prochlorococcus, Au arbitrary units.
The overall carrying capacity of the co-cultures was higher than the axenic Alteromonas cultures, and much higher than the axenic Prochlorococcus (Fig. 3B). On day 60, the mean carrying capacity of the co-cultures was 2–3 times higher than that of the Axenic Alteromonas (69 ± 35 compared with 32 ± 22 μmol N/L), suggesting that the heterotroph benefited from carbon fixed by the phototrophic Prochlorococcus partner. Indeed, most of this cellular N was found in the Alteromonas cells (76 ± 13%, see Supplementary Text S4 for a sensitivity analysis). The ability of axenic Alteromonas to survive in the absence of organic matter from Prochlorococcus is not surprising, as an Alteromonas strain distantly related to the ones studied here, AltSIO, can utilize a large fraction of the labile organic material found in natural seawater used to make the growth media [50]. In contrast, in axenic Prochlorococcus cultures only a small fraction of N in the system was found in cell biomass (~0.01 μmol N/L). This likely reflects the inability of all Prochlorococcus strains to recycle organic nitrogen lost due to exudation or cell lysis (Fig. 3B).
Mutual synergism was not observed across all strain combinations. While some Prochlorococcus strains (MED4, MIT0604, and NATL2A) supported significantly higher Alteromonas N biomass compared to the axenic control (log2FC 1.3 ± 0.6), co-cultures with MIT9312 and MIT9313 resulted in similar or lower Alteromonas biomass (log2FC −0.2 ± 0.9) (Fig. 3C). Therefore, on day 60, some of the interactions were mutually synergistic whereas in other cases Prochlorococcus do not support Alteromonas and may even compete with it. The two “non-mutually-synergistic” Prochlorococcus strains belong to different ecotypes but were isolated from the same drop of water from the Gulf Stream [51]. In all co-cultures the Prochlorococcus population benefited from the presence of Alteromonas (log2FC 10 ± 4 in N biomass compared to axenic controls).
In contrast to day 60, after 100 days essentially all of the interactions were mutually synergistic, with Alteromonas supporting the growth of all Prochlorococcus strains (log2FC 10 ± 1) and Prochlorococcus increasing the Alteromonas biomass in all strains with the exception of BS11 (log2FC 3 ± 1.5) (Fig. 3C). This suggests that the mode of interaction (synergist verses competition) may change temporally during the extended period of N starvation.
On day 140 the carrying capacity of the co-cultures declined further (only 1% of the N was in biomass). This suggests that the system is not in steady state, with a slow yet constant reduction in carrying capacity. We speculate that this is driven by the loss of bioavailable N from the system (i.e., most of the nitrogen is in a recalcitrant form that cannot be utilized by either partner). This is further supported by the observation that while the co-cultures after 140 days were still alive and could be transferred to new media, in a subsequent experiment, only 16/30 cultures could be transferred after 195 days, and only 3/75 cultures could be transferred after 250 days (Supplementary Fig. S5). MIT0604 (HLII) was the strain most likely to survive transfer after these extended periods and was also the most abundant Prochlorococcus strain after 140 days (1.46 ± 1 μmol N/L MIT0604 biomass verses 0.26 ± 0.56 μmol N/L for all other strains). While we do not currently have an explanation for the higher survival of this strain, it is noteworthy that it is the only strain to utilize nitrate [52].
Modeling the effect of co-culture on Prochlorococcus mortality
Given that the clearest effect of co-culture was on the decline phase of the co-cultures, we asked whether we could quantify and model the effect of Alteromonas on Prochlorococcus mortality. While the growth of bacteria has been extensively studied and modelled, the decline of bacterial cultures is much less studied, and mortality is rarely represented in ecological or biogeochemical models of microbial dynamics [53]. Bacterial mortality has, however, often been modelled in the context of food safety and genome evolution, using either mechanistic or descriptive approaches [53,54,55,56,57]. We chose to focus on four of these previously described models which are relatively simple and have a clear biological interpretation (Table 1). The exponential model is the simplest and most commonly used one, where a constant portion of the population dies over time [58]. The bi-exponential model is slightly more complex, representing two separate subpopulations in the community, each with its own death rate [55]. The Weibull model is probabilistic, modeling a heterogeneous population with a diverse stress tolerance [53, 59], finally, the harmonic model employs a quadratic rate of decline which is often associated with predator-prey interactions or cellular encounter rates [58]. When fitting each of these models to the decline phase of the growth curves, the Weibull model stands out as it has a low error for both axenic and co-cultures (Table 1) as well as in consequent transfers (Supplementary Table S2), the bi-exponential model is a better fit for the co-cultures but does not represent well the axenic ones. Based on the Weibull model, and assuming that culture fluorescence is related to the number of non-lysed cells in the media (Fig. S7), axenic Prochlorococcus cells die more than ten-fold faster than cells in co-culture (2-decimal reduction time, td2, is 12.58 ± 3.85 days for axenic cultures and 316 ± 337 days for co-cultures). Similar results were obtained with the bi-exponential model (Supplementary Text S5).
In the Weibull model, the “shape parameter” (n) represents the change over time (as the cultures decline) in the susceptibility of the bacterial community to stress. A shape parameter above one represents an increasing probability that cells will die as time increases (e.g. due to the accumulation of damage), whereas a shape parameter below one suggests that, as the culture declines, the cells become more resistant to damage. Axenic cultures have high mean shape value of 2.1 ± 0.9, suggesting an accumulation of cell damage leading to increasing death rate (Fig. 4A). In contrast, the mean shape value of co-cultures is significantly lower and below 1 (0.4 ± 0.2, student t-test, p < 0.001), suggesting that during N starvation in co-culture the Prochlorococcus cells are acclimating over time to the nutrient stress conditions.

A Weibull shape (n) in Axenic Prochlorococcus (PRO) and in Co-cultures. In all axenic samples n > 1, in most co-cultures n < 1 (students t-test, p < 0.001). B Scatter plot showing the reverse correlation between the total N biomass of the co-cultures on day 60 and Weibull shape (n). Pearson r = −0.33, p = 5e-3. Circles represent co-cultures with mutual synergistic interactions (i.e. Prochlorococcus strains MED4, MIT0604, and NATL2A), X represent potential competitive interactions (strains MIT9312, MIT9313). C Scatter plot showing the correlation between the total N biomass of the co-cultures on day 60 and Weibull root mean square error (RMSE). Pearson r = 0.64, p < 0.001. For the correlations with total N biomass on day 100 see Supplementary Fig. S6. D–F Weibull model fit for selected decline curves (FL). D Mutually synergistic co-culture of Prochlorococcus NATL2A and Alteromonas DE1. E Potentially competitive co-culture of Prochlorococcus MIT9312 and Alteromonas DE1. F Axenic Prochlorococcus NATL2A. FL fluorescence (arbitrary units). RMSE root mean square error.
While the molecular and physiological mechanisms of Prochlorococcus adaptation are currently unclear, the Weibull shape parameter decreases as the total N in cellular biomass increases, suggesting that the Prochlorococcus acclimation process is related to the ability to recycle N between the specific Prochlorococcus and Alteromonas strains in co-culture (Fig. 4B). Thus, the rate of acclimation is higher in the co-cultures supporting high N biomass and mutually synergistic interaction (NATL2A, MED4, and MIT0604) compared to MIT9312 and MIT9313, where Alteromonas do not gain from the interaction (0.36 ± 0.14 verses 0.52 ± 0.16, student t-test p < 0.001).
While the Weibull model is useful in quantifying mortality rates and raising the hypothesis that Prochlorococcus cells are acclimating to starvation over time, none of the tested models was able to fully capture the intricate dynamics of culture decline (Fig. 4C, D). In most mutually synergistic co-cultures involving Prochlorococcus strains NATL2A, MED4, and MIT0604, culture decline was not monotonic, and was interrupted by additional growth phases about 40–50 and 100 days after the cultures started declining (Fig. 4D). These latter growth phases were mostly absent in co-cultures with MIT9312 and MIT9313 (RMSE 0.36 ± 0.1 verses 0.2 ± 0.1 in the other strains, student t-test p < 0.001). The correlation between N biomass and the secondary growth phases (i.e., higher deviation from simple Weibull model; Fig. 4C) suggest that these phases may also be related to the ability of the interacting partners to recycle N through mutually beneficial metabolic interactions.
Conclusions and future prospects
Elucidating the mechanisms of microbial interactions requires well-characterized model systems. However, extending the insights from such models across the diversity of organisms and environmental conditions remains challenging. Our results from the highly simplified system of multiple Prochlorococcus and Alteromonas strains provide an important step towards this goal. Using the rich information on interaction phenotypes present in the growth and decline curves, we identify conserved and strain-specific facets of these interactions. Despite the genetic diversity across the Alteromonas strains studied [30], it was primarily the identity of the Prochlorococcus strain that determined the interaction phenotype. This manifests in the growth and decline rates, in the shape of the curve (primarily the decline phase), in the amount of N retained in biomass, and in whether the co-cultures are mutually synergistic or, potentially, competitive.
Under our laboratory conditions, it is likely that the combined response of both interacting partners to nitrogen starvation underlies the dynamics of the long-term co-cultures, although other stressors such as the increase in osmolarity/salinity or the accumulation of waste products cannot be ruled out [18, 34, 60]. This response is dynamic, as illustrated by the reproducible deviations of the fluorescence curves from the monotonic decline predicted by all models tested (“second growth” stages; Fig. 4). Three different (non-mutually-exclusive) processes may underlie these dynamics. Firstly, it is likely that one or both organisms modify their physiology or metabolism over time, for example through the activation of stringent responses, utilization of N or C storage pools, rewiring of metabolism to utilize available N sources, or activation of mechanisms such as extracellular enzymes allowing the cells to access previously unusable substrates (e.g. [61, 62]). Secondly, it is possible that there are “invisible” ecological dynamics underlying the observed fluorescence curves, for example cyclic changes in the abundance of Alteromonas cells. Under such a scenario, rapid Prochlorococcus mortality could produce an increase in Alteromonas abundance, resulting in degradation and remineralization of dead Prochlorococcus biomass and the release of resources that can drive subsequent Prochlorococcus growth. Thirdly, both Prochlorococcus and Alteromonas populations may be evolving, for example through emergence of genetically distinct populations better adapted to nutrient starvation (reminiscent of the GASP phenotype described in E. coli and other bacteria [63]).
Why is it the identity of the primary producer (Prochlorococcus) rather than the heterotrophic “recycler” (Alteromonas) that determines the outcome of the co-culture? A-priori, it was reasonable to assume that the co-culture phenotype would be affected by the differences between the Alteromonas strains in their ability to degrade and utilize polysaccharides and a variety of other organic molecules [30, 64]. We speculate that the increased growth of Alteromonas in the co-cultures compared to the axenic ones is fueled primarily by the availability of major biomass components released by Prochlorococcus as they die, such as proteins, amino acids, and nucleotides. Such common macromolecules do not require highly specialized metabolic processes to degrade and utilize, and hence can be utilized by all of the Alteromonas strains [65]. It is possible that the differences between Alteromonas strains may manifest when more complex macromolecules are available, e.g. from plant material, or when all of the “easy to digest” (labile) organic matter has been utilized and only complex macromolecules remain [66]. These conditions may not have been met in our experiments. It is also possible that co-culture with a more diverse range of heterotrophic bacteria, including additional Alteromonas species, would reveal more pronounced differences in the effect of the heterotroph of the primary producer. Similarly, we currently do not know why some Prochlorococcus strains support a mutually synergistic interaction with Alteromonas relatively early during the long-term N starvation (day 60) whereas other strains do not, and why at a later stage (day 100) almost all interactions are mutually beneficial. We could not identify any metabolic traits [11] clearly differentiating MIT9313 and MIT9312 (the “competitive” strains) from the others, suggesting more subtle differences exist between the Prochlorococcus strains in the organic matter they produce or in their response to N starvation (e.g. [67, 68]).
Our results identify patterns in the interactions between clades of abundant marine phototrophs and heterotrophs, under conditions where nutrients are scarce, and their availability likely depends on recycling between phototrophs and heterotrophs. Whether or not such mechanisms may be physiologically relevant in the oligotrophic ocean, much of which is N-stressed [40], remains to be tested. For example, in the oceans, rapid turnover of Prochlorococcus cells due to grazing and viral lysis likely means that cells are, on average, younger than those in laboratory cultures, which may affect their mortality rates [69]. Furthermore, stressors such as phage infection and grazing are missing in laboratory cultures. It is, however, noteworthy that the high heterotroph/phototroph biomass ratio observed during long-term N starvation here and in other studies [18] is similar to that of much of the open oligotrophic ocean (e.g. [70] and references therein). Additionally, Alteromonas may allow Prochlorococcus to adapt to light starvation [43] and to the presence of ROS (e.g. [71]), as well as other stressors that can be encountered in the open ocean. The supportive role of Alteromonas cannot be taken for granted, as it also depends on culture conditions, for example CO2 concentrations [27].
The co-cultures did not reach a steady state, and did not represent a closed system. Thus, processes not represented in these simplified laboratory co-cultures, are necessary to explain the long-term stability over decades of Prochlorococcus in the oceans [72]. Such processes could include multi-organism interactions, as natural communities are much more complex than the laboratory co-cultures, as well as oceanographic processes such as nutrient injection through deep mixing. More generally, cell mortality is intimately linked with the amount and type of recycled organic matter, yet the rate of mortality in natural communities is highly unconstrained [73]. Hence, better representation of mortality in mathematical models (e.g. the use of appropriate mortality formulations) is likely important for understanding biogeochemical cycles [73]. This may entail using one of the “off the shelf” models presented here, with their limitations (e.g. the Weibull model requires an estimate of the time of decline, whereas quadratic expressions for mortality are already used in such models [74,75,76]), or the development of new models that better link cell physiology, ecology, perhaps genome structure, and mortality.
Materials and methods
Strains and experiment set up
Axenic Prochlorococcus strains MED4 (HLI), MIT9312 (HLII), MIT0604 (HLII), NATL2A (LLI), and MIT9313 (LLIV) were maintained under constant cold while light (27 μmole photons m−2 s−1) at 22 °C [12, 77]. We used Pro99 media (natural seawater-based) that was modified by reducing the concentration of NH4 from 800 μM to 100 μM (Pro99-LowN), resulting in Prochlorococcus entering stationary stage due to the depletion of available inorganic N [78]. Alteromonas strains HOT1A3, Black sea 11, ATCC27126, AltDE1, and AltDE were maintained in ProMM [42]. Prior to the experiment, the axenicity of the Prochlorococcus cultures was tested by inoculating 1 ml culture into 15 ml ProMM [77], and no heterotrophic contaminants were observed by flow cytometry in axenic cultures after 60, 100, and 140 days. At the start of each co-culture experiment, Alteromonas cells from stationary-stage cultures (24–72 hour old) were centrifuged (10 minutes, room temperature, 10,000 g), the growth media decanted, and the cells re-suspended in the experimental media (Pro99-LowN). The Prochlorococcus cultures (growing exponentially) and the re-suspended Alteromonas cells were then counted using BD FACSCanto II Flow Cytometry Analyzer Systems (BD Biosciences). The initial cell concentrations in both co-cultures and axenic controls were 1 × 106 Prochlorococcus cells ml−1 and/or 1 × 107 Alteromonas cells ml−1. Axenic Alteromonas cultures were grown without any added C source besides DOC from the seawater-based media. The experiment was performed using triplicate 20 ml cultures in borosilicate test tubes (2.5 cm diameter, 15 cm length).
Fluorescence and Flow cytometry
Bulk chlorophyll fluorescence (FL) (ex440; em680) was measured almost daily using a Fluorescence Spectrophotometer (Cary Eclipse, Varian). Samples for flow cytometry were taken after 60, 100, and 140 days of experiment E1, fixed with glutaraldehyde (0.125% final concentration), incubated in the dark for 10 min and stored in −80 °C until analysis. Fluorescent beads (2 μm diameter, Polysciences, Warminster, PA, USA) were added as an internal standard. Data was acquired and processed with FlowJo software. Flow cytometry was performed unstained to count Prochlorococcus cells followed by staining with SYBR Green I (Molecular Probes/ ThermoFisher) to count Alteromonas cells.
Growth rate
Growth was computed––by solving the equation:
Where Nt represents the number of cells at time t, N0 is the initial number of cells, µ is the growth rate, and L is the growth lag. The LAN transformed equation was used to compute growth rate:
Linear regression was run on the growth phase, predicting ln(Nt) based on time t with R2 > 0.9. The growth rate µ is the regression coefficient.
Carrying capacity
The carrying capacity of the cultures was defined as the amount of nitrogen retained in cell biomass (rather than as dissolved organic N) at various stages of long-term co-culture. Cell numbers from flow cytometry were converted into nitrogen using 7 fg N cell−1 for the high-light strains MED4, MIT9312, and MIT0605, 10.5 fg N cell−1 for strain NATL2A and 14 fg N cell−1 for strain MIT9313 [79, 80]. For Alteromonas we used a value of 13 fg N cell−1 [50, 81]. We note that the values we used are at the lower end of measured cell values, which reach up to 20 fg N cell−1 for low-light Prochlorococcus and 25 fg N cell−1 for Alteromonas, since using the higher N cell quota leads to biomass that is higher than the total nitrogen available in the system (Supplementary Text S4). This assumption is supported by studies showing that cells contain less nitrogen under long-term N stress compared to exponential growth [33, 82, 83].
The cell numbers were converted to μmol/L by the formula:
Where X is the number of cells per ml, QN is the cell N quota, and MWN is the molecular weight of nitrogen. See Supplementary Text S4 for the detailed calculations and caveats.
Fit to decline models
The following functions were used to fit against the measured fluorescence:
Where FLt is the Fluorescence measured at time t, FLmax is the maximum fluorescence measured, tmax is the time when the fluorescence was highest, and a, a1, a2, n, f are the model parameters estimated by the fitting function.
The decline function was fit against each growth curve via curve_fit() function from scipy package (1.3.0), using the parameters: method=‘dogbox’, loss=‘soft_l1’, f_scale = 0.1. Each model was fit using random initial parameter values and the initial values of 0.5 per parameter, and the fit with the lowest RSME selected. Goodness of fit was measures using root mean square error (RMSE).
In the Weibull model the time needed to reduce the population by d factors of 10 (td2) was estimated as in [59], using the formula:
Random forest classification
To detect difference in the curve pattern and not timing specific differences, the curves were aligned such that max growth point are at time zero, and time points from 10 days prior to max growth to 80 days after were selected. Since the specific measurement time points were different in different experiments and samples, rolling average was used to get mean fluorescence per day, and interpolation used to fill in missing measurements. The Fluorescence measurements were standardized via standard scalar by subtracting the mean and scaling to unit variance of each feature. Random forest model was run in 10x cross validation [84] to classify the curves by Prochlorococcus and by Alteromonas strains. To find the most significant days in Prochlorococcus classification, the model was built 30 times and the mean importance of all features (i.e., measurement days) calculated. Data preprocessing was done by pandas (0.25.0). Scaling and model fitting using sklearn (0.21.2).
PCA ordination
PCA ordination was run on the growth curves. The fluorescence measurements were standardized via standard scalar by subtracting the mean and scaling to unit variance of each feature. Ordination was computed via principal component analysis (PCA). Data preprocessing was done by pandas (0.25.0). Scaling and PCA was done using sklearn (0.21.2).
Statistics
Statistics were computed using the statsmodels package in python. Multi test correction was done by t_test_pairwise() using Bonferroni correction. Permanova analysis by adonis2 from R vegan package (R 3.61, vegan 2.5-7).
Data availability
All the code and data used in this manuscript is available on https://github.com/wosnat/ccpa.
References
Gralka M, Szabo R, Stocker R, Cordero OX. Trophic interactions and the drivers of microbial community assembly. Curr Biol. 2020;30:R1176–R1188.
Fritts RK, McCully AL, McKinlay JB. Extracellular metabolism sets the table for microbial cross-feeding. Microbiol Mol Biol Rev. 2021;85:1–29.
Weisskopf L, Schulz S, Garbeva P. Microbial volatile organic compounds in intra-kingdom and inter-kingdom interactions. Nat Rev Microbiol. 2021;19:391–404.
Harrington V, Lau L, Seddu K, Suez J. Ecology and medicine converge at the microbiome-host. mSystems. 2021;6:4–8.
Long RA, Azam F. Microscale patchiness of bacterioplankton assemblage richness in seawater. Aquat Micro Ecol. 2001;26:103–13.
Cordero OX, Wildschutte H, Kirkup B, Proehl S, Ngo L, Hussain F, et al. Ecological populations of bacteria act as socially cohesive units of antibiotic production and resistance. Science. 2012;337:1228–31.
Grossart HP, Simon M. Interactions of planktonic algae and bacteria: Effects on algal growth and organic matter dynamics. Aquat Micro Ecol. 2007;47:163–76.
Segev E, Wyche TP, Kim KH, Petersen J, Ellebrandt C, Vlamakis H, et al. Dynamic metabolic exchange governs a marine algal-bacterial interaction. Elife. 2016;5:1–28.
Seyedsayamdost MR, Case RJ, Kolter R, Clardy J. The Jekyll-and-Hyde chemistry of Phaeobacter Gallaeciensis. Nat Chem. 2011;3:331–5.
Antoniewicz MR. A guide to deciphering microbial interactions and metabolic fluxes in microbiome communities. Curr Opin Biotechnol. 2020;64:230–7.
Zoccarato L, Sher D, Miki T, Segrè D, Grossart H-P. A comparative whole-genome approach identifies bacterial traits for marine microbial interactions. Commun Biol. 2022;5:1–13.
Sher D, Thompson JW, Kashtan N, Croal L, Chisholm SW. Response of Prochlorococcus ecotypes to co-culture with diverse marine bacteria. ISME J. 2011;5:1125–32.
Warringer J, Anevski D, Liu B, Blomberg A. Chemogenetic fingerprinting by analysis of cellular growth dynamics. BMC Chem Biol. 2008;8:1–10.
Monod J. The growth of bacterial cultures. Annu Rev M. 1949;3:371–94.
Foster KR, Bell T. Competition, not cooperation, dominates interactions among culturable microbial species. Curr Biol. 2012;22:1845–50.
Pacheco AR, Osborne ML, Segrè D. Non-additive microbial community responses to environmental complexity. Nat Commun. 2021;12:1–11.
Vetsigian K, Jajoo R, Kishony R. Structure and evolution of Streptomyces interaction networks in soil and in silico. PLoS Biol. 2011;9:1–12.
Christie-Oleza JA, Sousoni D, Lloyd M, Armengaud J, Scanlan DJ. Nutrient recycling facilitates long-term stability of marine microbial phototroph–heterotroph interactions. Nat Microbiol. 2017;2:17100.
Ratzke C, Gore J. Modifying and reacting to the environmental pH drives bacterial interactions. PLoS Biol. 2018;16:e2004248.
Grossart HP. Interactions between marine bacteria and axenic diatoms (Cylindrotheca fusiformis, Nitzschia laevis, and Thalassiosira weissflogii) incubated under various conditions in the lab. Aquat Micro Ecol. 1999;19:1–11.
Field CB, Behrenfeld MJ, Randerson JT, Falkowski P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science (80-). 1998;281:237–40.
Cole JJ. Interaction between bacteria and alge in aquatic ecosystems. Ann Rev Ecol Syst. 1982;13:291–314.
Seymour JR, Amin SA, Raina JB, Stocker R. Zooming in on the phycosphere: the ecological interface for phytoplankton-bacteria relationships. Nat Microbiol. 2017;2:17065.
Ramanan R, Kim BH, Cho DH, Oh HM, Kim HS. Algae-bacteria interactions: evolution, ecology and emerging applications. Biotechnol Adv. 2016;34:14–29.
Cirri E, Pohnert G. Algae−bacteria interactions that balance the planktonic microbiome. N. Phytol. 2019;223:100–6.
Morris JJ, Lenski RE, Zinser ER. The black queen hypothesis: evolution of dependencies through adaptive gene loss. mBio. 2012;3:1–7.
Hennon GM, Morris JJ, Haley ST, Zinser ER, Durrant AR, Entwistle E, et al. The impact of elevated CO2 on Prochlorococcus and microbial interactions with ‘helper’ bacterium. Alteromonas ISME J. 2018;12:520–31.
Biller SJ, Berube PM, Lindell D, Chisholm SW. Prochlorococcus: The structure and function of collective diversity. Nat Rev Microbiol. 2015;13:13–27.
López-Pérez M, Rodriguez-Valera F. Pangenome evolution in themarine bacterium Alteromonas. Genome Biol Evol. 2016;8:1556–70.
Koch H, Germscheid N, Freese HM, Noriega-Ortega B, Lücking D, Berger M, et al. Genomic, metabolic and phenotypic variability shapes ecological differentiation and intraspecies interactions of Alteromonas macleodii. Sci Rep. 2020;10:1–14.
Hou S, López-Pérez M, Pfreundt U, Belkin N, Stüber K, Huettel B, et al. Benefit from decline: the primary transcriptome of Alteromonas macleodii str. Te101 during Trichodesmium demise. ISME J. 2018;12:981–96.
Aharonovich D, Sher D. Transcriptional response of Prochlorococcus to co-culture with a marine Alteromonas: Differences between strains and the involvement of putative infochemicals. ISME J. 2016;10:2892–906.
Roth-Rosenberg D, Aharonovich D, Luzzatto-Knaan T, Vogts A, Zoccarato L, Eigemann F, et al. Prochlorococcus cells rely on microbial interactions rather than on chlorotic resting stages to survive long-term nutrient starvation. mBio. 2020;11:1–13.
Morris JJ, Johnson ZI, Szul MJ, Keller M, Zinser ER. Dependence of the cyanobacterium Prochlorococcus on hydrogen peroxide scavenging microbes for growth at the ocean’s surface. PLoS One. 2011;6:e16805.
Diner RE, Schwenck SM, McCrow JP, Zheng H, Allen AE. Genetic manipulation of competition for nitrate between heterotrophic bacteria and diatoms. Front Microbiol. 2016;7:880.
Biller SJ, Coe A, Chisholm SW. Torn apart and reunited: Impact of a heterotroph on the transcriptome of Prochlorococcus. ISME J. 2016;10:2831–43.
Zheng Q, Wang Y, Lu J, Lin W, Chen F, Jiao N. Metagenomic and metaproteomic insights into photoautotrophic and heterotrophic interactions in a Synechococcus culture. mBio. 2020;11:e03261-19.
Calfee BC, Glasgo LD, Zinser ER. Prochlorococcus exudate stimulates heterotrophic bacterial competition with rival phytoplankton for available nitrogen. mBio. 2022;13:e0257121.
Amin SA, Hmelo LR, Van Tol HM, Durham BP, Carlson LT, Heal KR, et al. Interaction and signalling between a cosmopolitan phytoplankton and associated bacteria. Nature. 2015;522:98–101.
Ustick LJ, Larkin AA, Garcia CA, Garcia NS, Brock ML, Lee JA, et al. Metagenomic analysis reveals global-scale patterns of ocean nutrient limitation. Science (80-). 2021;372:287–91.
Saito MA, McIlvin MR, Moran DM, Goepfert TJ, DiTullio GR, Post AF, et al. Multiple nutrient stresses at intersecting Pacific Ocean biomes detected by protein biomarkers. Science (80-). 2014;345:1173–7.
Morris JJ, Kirkegaard R, Szul MJ, Johnson ZI, Zinser ER. Facilitation of robust growth of Prochlorococcus colonies and dilute liquid cultures by ‘helper’ heterotrophic bacteria. Appl Environ Microbiol. 2008;74:4530–4.
Coe A, Ghizzoni J, LeGault K, Biller S, Roggensack SE, Chisholm SW. Survival of Prochlorococcus in extended darkness. Limnol Oceanogr. 2016;61:1375–88.
Ting CS, Hsieh C, Sundararaman S, Mannella C, Marko M. Cryo-electron tomography reveals the comparative three-dimensional architecture of Prochlorococcus, a globally important marine cyanobacterium. J Bacteriol. 2007;189:4485–93.
Klähn S, Steglich C, Hess WR, Hagemann M. Glucosylglycerate: A secondary compatible solute common to marine cyanobacteria from nitrogen-poor environments. Environ Microbiol. 2010;12:83–94.
Cubillos-Ruiz A, Berta-Thompson JW, Becker JW, Van Der Donk WA, Chisholm SW. Evolutionary radiation of lanthipeptides in marine cyanobacteria. Proc Natl Acad Sci USA. 2017;114:E5424–E5433.
Li B, Sher D, Kelly L, Shi Y, Huang K, Knerr PJ, et al. Catalytic promiscuity in the biosynthesis of cyclic peptide secondary metabolites in planktonic marine cyanobacteria. Proc Natl Acad Sci USA. 2010;107:10430–5.
Yelton AP, Acinas SG, Sunagawa S, Bork P, Pedrós-Alió C, Chisholm SW. Global genetic capacity for mixotrophy in marine picocyanobacteria. ISME J. 2016;10:2946–57.
Kettler GC, Martiny AC, Huang K, Zucker J, Coleman ML, Rodrigue S, et al. Patterns and implications of gene gain and loss in the evolution of Prochlorococcus. PLoS Genet. 2007;3:2515–28.
Pedler BE, Aluwihare LI, Azam F. Single bacterial strain capable of significant contribution to carbon cycling in the surface ocean. Proc Natl Acad Sci USA. 2014;111:7202–7.
Moore LR, Rocap G, Chisholm SW. Physiology and molecular phylogeny of coexisting Prochlorococcus ecotypes. Nature. 1998;393:464–7.
Hackl T, Laurenceau R, Ankenbrand MJ, Bliem C, Cariani Z, Thomas E, et al. Novel integrative elements and genomic plasticity in ocean ecosystems. bioRxiv. 2020.
Shoemaker WR, Jones SE, Muscarella ME, Behringer MG, Lehmkuhl BK, Lennon JT. Microbial population dynamics and evolutionary outcomes under extreme energy limitation. Proc Natl Acad Sci USA. 2021;118:1–8.
Brouwer AF, Eisenberg MC, Remais JV, Collender PA, Meza R, Eisenberg JNS. Modeling biphasic environmental decay of pathogens and implications for risk analysis. Environ Sci Technol. 2017;51:2186–96.
Crane SR, Moore JA. Modeling enteric bacterial die-off: a review. Water, Air, Soil Pollut. 1986;27:411–39.
Finkel SE, Kolter R. Evolution of microbial diversity during prolonged starvation. Proc Natl Acad Sci USA. 1999;96:4023–7.
Patra P, Klumpp S. Population dynamics of bacterial persistence. PLoS One. 2013;8:e62814.
Pruitt KM, Kamau DN. Mathematical models of bacterial growth, inhibition and death under combined stress conditions. J Ind Microbiol. 1993;12:221–31.
Van Boekel MAJS. On the use of the Weibull model to describe thermal inactivation of microbial vegetative cells. Int J Food Microbiol. 2002;74:139–59.
Aguilo-Ferretjans MDM, Bosch R, Puxty RJ, Latva M, Zadjelovic V, Chhun A, et al. Pili allow dominant marine cyanobacteria to avoid sinking and evade predation. Nat Commun. 2021;12:1–10.
Hood RD, Higgins SA, Flamholz A, Nichols RJ, Savage DF. The stringent response regulates adaptation to darkness in the cyanobacterium Synechococcus elongatus. Proc Natl Acad Sci USA. 2016;113:E4867–E4876.
Bergkessel M, Basta DW, Newman DK. The physiology of growth arrest: uniting molecular and environmental microbiology. Nat Rev Microbiol. 2016;14:549–62.
Finkel SE. Long-term survival during stationary phase: evolution and the GASP phenotype. Nat Rev Microbiol. 2006;4:113–20.
Koch H, Dürwald A, Schweder T, Noriega-Ortega B, Vidal-Melgosa S, Hehemann JH, et al. Biphasic cellular adaptations and ecological implications of Alteromonas macleodii degrading a mixture of algal polysaccharides. ISME J. 2019;13:92–103.
Elena F, Daniel S, Daniel S. Metabolic phenotyping of marine heterotrophs on refactored media reveals diverse metabolic adaptations and lifestyle strategies. mSystems. 2022;e00070–22.
Teeling H, Fuchs BM, Becher D, Klockow C, Gardebrecht A, Bennke CM, et al. Substrate-controlled succession of marine bacterioplankton populations induced by a phytoplankton bloom. Science. 2012;336:608–11.
Becker JW, Berube PM, Follett CL, Waterbury JB, Chisholm SW, DeLong EF, et al. Closely related phytoplankton species produce similar suites of dissolved organic matter. Front Microbiol. 2014;5:1–14.
Domínguez-Martín MA, López-Lozano A, Clavería-Gimeno R, Velázquez-Campoy A, Seidel G, Burkovski A, et al. Differential NtcA responsiveness to 2-oxoglutarate underlies the diversity of C/N balance regulation in Prochlorococcus. Front Microbiol. 2018;8:1–16.
Lindner AB, Madden R, Demarez A, Stewart EJ, Taddei F. Asymmetric segregation of protein aggregates is associated with cellular aging and rejuvenation. Proc Natl Acad Sci USA. 2008;105:3076–81.
Reich T, Ben-Ezra T, Belkin N, Tsemel A, Aharonovich D, Roth-Rosenberg D, et al. A year in the life of the Eastern Mediterranean: Monthly dynamics of phytoplankton and bacterioplankton in an ultra-oligotrophic sea. Deep Res Part I Oceanogr Res Pap. 2022;182:103720.
Ma L, Calfee BC, Morris JJ, Johnson ZI, Zinser ER. Degradation of hydrogen peroxide at the ocean’s surface: The influence of the microbial community on the realized thermal niche of Prochlorococcus. ISME J. 2018;12:473–84.
Malmstrom RR, Coe A, Kettler GC, Martiny AC, Frias-Lopez J, Zinser ER, et al. Temporal dynamics of Prochlorococcus ecotypes in the Atlantic and Pacific Oceans. ISME J. 2010;4:1252–64.
Beckett SJ, Demory D, Coenen AR, Casey JR, Dugenne M, Follett CL, et al. Diel population dynamics and mortality of Prochlorococcus in the North Pacific Subtropical Gyre. bioRxiv. 2021.
Moore SK, Baird ME, Suthers IM. Relative effects of physical and biological processes on nutrient and phytoplankton dynamics in a shallow estuary after a storm event. Estuaries Coasts. 2006;29:81–95.
Murray AG, Parslow JS. The analysis of alternative formulations in a simple model of a coastal ecosystem. Ecol Model. 1999;119:149–66.
Dutkiewicz S, Follows M, Marshall J, Gregg WW. Interannual variability of phytoplankton abundances in the North Atlantic. Deep Sea Res Part II Top Stud Oceanogr. 2001;48:2323–44.
Moore LR, Coe A, Zinser ER, Saito MA, Sullivan MB, Lindell D, et al. Culturing the marine cyanobacterium Prochlorococcus. Limnol Oceanogr Methods. 2007;5:353–62.
Grossowicz M, Roth-Rosenberg D, Aharonovich D, Silverman J, Follows MJ, Sher D. Prochlorococcus in the lab and in silico: The importance of representing exudation. Limnol Oceanogr. 2017;62:818–35.
Martiny AC, Ma L, Mouginot C, Chandler JW, Zinser ER. Interactions between thermal acclimation, growth rate, and phylogeny influence Prochlorococcus elemental stoichiometry. PLoS One. 2016;11:1–12.
Bertilsson S, Berglund O, Karl DM, Chisholm S. Elemental composition of marine Prochlorococcus and Synechococcus: Implications for the ecological stoichiometry of the sea. Limnol Oceanogr. 2003;48:1721–31.
Posacka AM, Semeniuk DM, Maldonado MT. Effects of copper availability on the physiology of marine heterotrophic bacteria. Front Mar Sci. 2019;5:1–19.
Liefer JD, Garg A, Fyfe MH, Irwin AJ, Benner I, Brown CM, et al. The macromolecular basis of phytoplankton C:N:P under nitrogen starvation. Front Microbiol. 2019;10:1–16.
Chan LK, Newton RJ, Sharma S, Smith CB, Rayapati P, Limardo AJ, et al. Transcriptional changes underlying elemental stoichiometry shifts in a marine heterotrophic bacterium. Front Microbiol. 2012;3:1–24.
Browne MW. Cross-validation methods. J Math Psychol. 2000;44:108–32.
Acknowledgements
We thank Sher lab members and especially Natalie Andrawes for help in fluorescence measurements and Daniel Segrè, Hans-Peter Grossart, Zhen Wu and Tal Luzzatto Knaan for critical reading of the manuscript. This work was supported by the Human Frontiers Science Program (grant RGP0020/2016, to DS), the Israel Science Foundation (grant 1786/20 to DS), and the U.S.-Israel Binational Science Foundation (BSF) - U.S. National Science Foundation (NSF) program in Oceanography (grant 1635070/2016532, to DS). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
DA and DS designed the experiment, DA performed experiments, DA, OW, and DS analyzed experimental results. OW designed and executed the data analysis and modeling. DA, OW, and DS wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Weissberg, O., Aharonovich, D. & Sher, D. Phototroph-heterotroph interactions during growth and long-term starvation across Prochlorococcus and Alteromonas diversity. ISME J 17, 227–237 (2023). https://doi.org/10.1038/s41396-022-01330-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41396-022-01330-8
This article is cited by
-
Pseudomonas putida as saviour for troubled Synechococcus elongatus in a synthetic co-culture – interaction studies based on a multi-OMICs approach
Communications Biology (2024)
-
The β-Lactamase Activity at the Community Level Confers β-Lactam Resistance to Bloom-Forming Microcystis aeruginosa Cells
Journal of Microbiology (2023)
