
Abstract
Background
Special AT-rich sequence-binding protein 2 is essential for the development of cerebral cortex and key molecular node for the establishment of proper neural circuitry and function. Mutations in the SATB2 gene lead to SATB2-associated syndrome, which is characterized by abnormal development of skeleton and central nervous systems.
Methods
We generated Satb2 knockout mouse model through CRISPR-Cas9 technology and performed RNA-seq and ChIP-seq of embryonic cerebral cortex. We conducted RT-qPCR, western blot, immunofluorescence staining, luciferase reporter assay and behavioral analysis for experimental verification.
Results
We identified 1363 downstream effector genes of Satb2 and correlation analysis of Satb2-targeted genes and neurological disease genes showed that Satb2 contribute to cognitive and mental disorders from the early developmental stage. We found that Satb2 directly regulate the expression of Ntng1, Cdh13, Kitl, genes important for axon guidance, synaptic formation, neuron migration, and Satb2 directly activates the expression of Mef2c. We also showed that Satb2 heterozygous knockout mice showed impaired spatial learning and memory.
Conclusions
Taken together, our study supportsroles of Satb2 in the regulation of axonogenesis and synaptic formation at the early developmental stage and provides new insights into the complicated regulatory mechanism of Satb2 and new evidence to elucidate the pathogen of SATB2-associated syndrome.
Impact
-
1363 downstream effector genes of Satb2 were classified into 5 clusters with different temporal expression patterns.
-
We identified Plxnd1, Ntng1, Efnb2, Ephb1, Plxna2, Epha3, Plxna4, Unc5c, and Flrt2 as axon guidance molecules to regulate axonogenesis.
-
168 targeted genes of Satb2 were found to regulate synaptic formation in the early development of the cerebral cortex.
-
Transcription factor Mef2c is positively regulated by Satb2, and 28 Mef2c-targeted genes can be directly regulated by Satb2.
-
In the Morris water maze test, Satb2+/− mice showed impaired spatial learning and memory, further strengthening that Satb2 can regulate synaptic functions.
Similar content being viewed by others

Introduction
Special AT-rich sequence-binding protein 2(SATB2) is a transcription factor that plays essential roles in chromatin remodeling and regulation of gene expression in a nuclear-matrix-attachment regions (MARs) dependent manner.1 Satb2 has been demonstrated to function in multiple biological processes, including craniofacial patterning, bone formation, cortical regionalization, development of corpus callosum and neuron projection in the neocortex.2,3,4 Satb2 is also important as a regulator of synaptic plasticity in the hippocampus that underlies memory functions.5 In humans, microdeletions or mutations of the SATB2 gene lead to SATB2-associated syndromes (SAS) or Glass syndrome (OMIM 612,313), which are clinically manifested as craniofacial malformation, teeth anomalies, developmental delay, poor speech development, hyperactivity, intellectual disability, seizures, and symptoms of autism.6,7
In developing mouse brain, Satb2 expressing neurons extend axons across the corpus callosum. While in Satb2 knockout mice, axons cannot pass through the corpus callosum instead descend along the corticospinal tract and neurons failed to reach their targeted regions.3 Satb2 was reported to regulate the differentiation of both callosal and subcerebral projection neurons in the developing cerebral cortex.8 Generation and outgrowth of axons, termed as axonogenesis, are critical for establishing and maintaining the polarized structure of the neurons.9 Proper axon guidance is essential in neuron migration and projection to their targets.10,11 Satb2 was known to regulate genes involved in axonogenesis, such as Ctip2, Unc5c, and Epha7.12,13,14 The cerebral cortex plays a central role in high-level cognitive functions such as learning, memory, thinking and decision-making.15 Synaptic transmission between neurons, commonly termed synaptic plasticity, are key for primarily learning and memory.16 Many researches demonstrate functions of Satb2 in neuronal projection, axonogenesis and synaptic functions in the cerebral cortex, but molecular mechanisms of Satb2 underlining these functions are still not fully understood.
In this study, to further understand molecular mechanisms of Satb2 in the early development of cerebral cortex, we generated Satb2 knockout mice, and performed RNA-seq and Chromatin Immunoprecipitation sequence (ChIP-seq) and functional study to understand functions of Satb2 in the development of the cortical cortex.
Methods
Experimental animals
All animal research was approved by the Children’s Hospital of Fudan University (2014-025). All experimental mice were fed in a specific pathogen-free room with automatically controlled temperature (16 °C), humidity (40%), ventilation and light conditions. Satb2 knock out model was generated through CRISPR-Cas9 technology by Biocytogen Pharmaceuticals (Beijing) Co., Ltd. Genotypes were determined by polymerase chain reaction (PCR) and agarose gel electrophoresis with two pairs of primers near the deleted sequence (Table S1).
RNA-seq analyses
RNA-seq data were processed through the Nextflow nf-core/rnaseq pipeline (v3.4).17 Differential expression analysis was performed for replicates using DESeq2 (v1.30)18 and genes with a false discovery rate (FDR) < 0.1 were considered as differentially expressed genes, to be consistent with FDR of 0.1 for cutoff in P0 RNA-seq analysis. P0 cortex data from McKenna et al. (GSE68911)19 and adult cortex data from Cera et al. (GSE123992)20 were re-analyzed using the same pipeline for E17.5 cortex and the same threshold cutoffs for differential expression were applied.
ChIP-seq
ChIP reactions were used 22 μg of embryonic mouse brain tissue chromatin and 4 μg of anti-SATB2 antibody (Novus, cat# NBP176912). The library of Satb2+/+ ChIP DNA fraction was generated according to the manufacture protocol (Active Motif). The 75nt single-end sequence reads were obtained by Illumina sequencing NextSeq500. Reads were aligned to the mouse genome (mm10) by BWA (v 0.7.17),21 and after removal of duplicate and non-uniquely mapped reads by Picard (v2.23.1) (http://broadinstitute.github.io/picard/). A signal map capturing fragment densities along the genome was generated and visualized in the Integrated Genome Browser (IGB). MACS2 (v2.2.7)22 was used to call narrow peaks with FDR < 0.001. Peaks were annotated using the R package ChIPseeker (1.31.3).23
Gene function and disease enrichment analyses
Gene ontology (GO), kyoto encyclopedia of genes and genomes (KEGG) and human phenotype ontology (HPO) enrichment analysis was performed using the R package clusterProfiler (v4.3.4).24 Sets with less than 10 (5 for HPO) or more than 500 genes were omitted. p-values were corrected using the Benjamini–Hochberg (BH) method. Gene sets with FDR < 0.05 (FDR < 0.2 for KEGG) were considered to be significant and the top ten sets were plotted. Ontology diagrams were visualized using ontologyPlot (v1.6). KEGG pathways were plotted by the R package pathview (v1.31.3).25 HPO annotations for mouse genes were acquired from Molecular Signatures Database (MSigDB) through the R package msigdbr (v7.4.1). 707 neurological disease genes were downloaded from OMIM website (https://www.omim.org/) using “Head & neck, CS, Neurologic” as the clinical synopsis search quires. Disease genes were mapped to the orthologous mouse genes through annotations from the Mouse Genome Informatics (MGI) database (http://www.informatics.jax.org/). The list of genes encoding mouse transcription factors was downloaded from the AnimalTFDB database.26 The p-values for the overlapping gene sets were calculated by the Fisher exact test.
Results
Generation of Satb2 knockout mouse model
Satb2 knockout mouse model was generated through CRISPR-Cas9 technology (Fig. 1a). Real-time quantification PCR(RT-qPCR) showed significantly decreased mRNA level of Satb2 in Satb2−/− (p < 0.001) and Satb2+/− (p < 0.001) mice when compared with Satb2+/+ mice (Fig. 1b). Western blot analysis of cerebral cortex lysates showed a lower expression of SATB2 protein in Satb2+/− mice and nearly no expression in Satb2−/− mice (Fig. 1c). Relative quantification of bands also verified knockout effects (Fig. 1d). We performed immunofluorescence staining of frozen sections of mouse brain to further test the knockout efficiency of SATB2 in situ in the cerebral cortex. Consistent with previous reports, most SATB2-positive cells reside in layers 2/3/4 and a few numbers of SATB2-positive cells are present in layer 5 and even fewer in deep layer 6 in wild-type mice (Fig. 1e). In contrast, SATB2 was not obviously expressed in neocortex region of Satb2−/− mice brain. Satb2 was reported to promote the development of callosal neurons through repressing expression of CTIP2, which is expressed in Layer 5/6 neurons and expanded into layer 2/3/4 in the absence of SATB2.3,12 Taken together, we successfully generated a Satb2 knockout mouse model for further study.

a Knockout strategy through targeting intron 6 and intron 8 of Satb2 gene by CRISPR-Cas9 technology. b qRT-PCR of Satb2 in the cerebral cortex of E17.5 Satb2+/+, Satb2+/−, and Satb2−/− mice. Unpaired t-test, ∗∗∗p < 0.001, n = 3, error bars represent standard error of mean. c Representative western blot images of SATB2 protein in the cerebral cortex of E17.5 Satb2+/+, Satb2+/−, and Satb2−/− mice. d Relative quantitative analysis of western blot bands. Unpaired t-test, ∗∗∗p < 0.001, ∗∗∗∗p < 0.0001, n = 3. e Representative images of SATB2 and CTIP2 immunostaining in E17.5 Satb2+/+ and Satb2−/− cortex. Scale bar, 100 μm.
Identification of dysregulated Satb2-targeted genes in the developing cerebral cortex
To decipher mechanisms of Satb2 in callosal projection and neuronal development, we performed RNA-seq of cerebral cortex samples from wild-type and homozygous mutant mice at E17.5. Compared with Satb2+/+ mice, 144 genes were significantly upregulated and 168 genes were downregulated in Satb2−/− mice (Fig. S1A and Table S5). GO of these differentially expressed genes mainly enriched on axonogenesis, axon guidance, neuron differentiation, neuron projection extension, synapse function, dendrite development, cell adhesion and cognition (Fig. S1B). Satb2 has been reported to promote the development of callosal and subcerebral neurons in a cell context-dependent manner. We analyzed the expression of genes important for the development of callosal projection neurons(CPN) and subcerebral projection neurons(ScPN) (Fig. S1C, D). Most CPN marker genes, such as Cux1/2 and Pou3f2/3, were significantly decreased, while most ScPN marker genes, such as Ctip2 and Sox5, were significantly increased, which means the transcriptome characteristics of cerebral cortex were transformed from CPN to ScPN in Satb2−/− mouse.
ChIP-seq of fresh cortices dissected from E17.5 embryos helped us identify 8719 genomic regions (also referred to as peaks) enriched in SATB2-precipitated DNA, which mapped to a total of 4834 gene loci in the Satb2+/+ group (Fig. S2A). All peak regions distributed ±3 kb upstream of transcription start sites (TSS) (Fig. S2B). The detailed location of peaks relative to genomic annotations was presented in a pie chart (Fig. S2C).
To identify Satb2-targeted genes in the 4834 candidate genes, we integrated our E17.5 RNA-seq data and P0 RNA-seq data published by Mckenna et al.19 Re-analysis of P0 RNA-seq data with identical bioinformatics procedures applied in the above E17.5 RNA-seq dataset helped us identify 4209 differentially expressed genes (DEGs) in either E17.5 or P0 Satb2-mutant cortices. 1363 genes overlapped between DEGs and ChIP peaks associated genes, suggesting these genes could be Satb2-targeted genes (Fig. 2a and Table S6). According to the GO and KEGG pathway analysis, there is a significant enrichment for genes with roles in synapse organization, axonogenesis, cell junction assembly, cognition, learning or memory and cell adhesion, indicating that Satb2 regulates various aspects of axongenesis and synaptic function in cerebral cortex (Fig. 2b, c). Next, we want to know whether these Satb2-targeted genes contribute to SAS, so we downloaded neurological disease genes from OMIM website, and performed correlation analysis with Satb2-targeted genes. We found Satb2 targets were enriched for 178 transcription factor (p = 1.269e-12) and 73 neurological disease genes (p = 0.0007568) (Fig. 2d).

a Venn diagram shows the 1363 overlapping genes between SATB2-binding genes and E17.5 DEGs or P0 DEGs. b GO enrichment of Satb2-targeted genes (adjusted p-value < 0.05). c KEGG pathway enrichment of Satb2-targeted genes (adjusted p-value < 0.05). d Venn diagram shows relations between Satb2-targeted genes, transcription factors and neurological disease genes. Satb2 targets are enriched for transcription factors (178 genes, p = 1.269e-12) and neurological disease genes (73 genes, p = 0.0007568).
Developmental expression pattern of Satb2-targeted genes
To explore the developmental functions of the Satb2-targeted genes, we investigated their developmental expression trajectories in the wild-type cerebral cortex based on previous published RNA-seq data.27 The expression profile of the 1363 gene showd noticeable temporal changes and can be grouped into five clusters using soft-clustering method28 (Fig. 3a, b). The five cluster genes regulated by Satb2 were further classified into three groups with different biological functions that were C1 and C2 genes that regulate neurodevelopment, C3 genes that regulate cell adhesion, C4 and C5 genes that regulate synaptic related functions. Functional differences in the three groups are consistent with the role of Satb2 as a cell fate and neuron projection determinant at neonatal stage and regulator of synaptic plasticity/physiology at the adult stage.20 GO analysis of each cluster is consistent with differences in biological processes of Satb2 in different developmental stages (Fig. 3c).

a Five clusters of Satb2-targeted genes with different temporal expression patterns. b Expression heatmap of Satb2-targeted genes. Bottom, developmental stage of RNA-seq samples; Left, gene cluster with up- or downregulated features; Right, DEG Satb2-targeted genes in E17.5 and P0. c Biological process, molecular function, and cellular component in GO analysis of each cluster. d Increased expression of Kitl in Satb2-/- mouse cortex. e SATB2 binds to conserved intronic sequences of Kitl gene. Red arrows indicate a binding peak. f Relative luciferase activity changes when co-transfection of SATB2 or SATB2 R239X and Kitl-S-pEZX. **p < 0.01, ***p < 0.001, n = 3.
Kitl encodes KITL (stem cell factor, SCF), a ligand growth factor for c-kit, and functions in neuronal migration and survival, so we choose Kitl as a representative of C3 genes to verify whether they are directly regulated by Satb2.29,30 Both RNA-seq data and RT-qPCR results showed significantly increased expression of Kitl in Satb2−/− mouse cerebral cortex (Fig. 3d, p-value < 0.01). ChIP-seq analysis identified a strong and specific binding of SATB2 to intron 1 of Kitl gene (Fig. 3e). To test whether SATB2 can directly regulate the expression of Kitl, we performed luciferase reporter assay. When co-transfected with SATB2, luciferase activity significantly decreased in Kitl-S-pEZX group compared with the empty pEZX group (Fig. 3f). We found a de novo SATB2 c.715 C > T (p.R239X) mutation in a 3-year-old boy who was diagnosed with SAS. The mutation had been reported in literatures and was thought of as a hot spot mutation. Individuals with the R239X mutation usually exhibited craniofacial dysmorphism, generalized osteoporosis, profound mental retardation and epilepsy.31 When co-transfected mutant SATB2 R239X with Kitl-S-pEZX, the activity of luciferase increased compared with SATB2 and Kitl-S-pEZX group, suggesting that mutant SATB2 R239X weakened the repression activity of SATB2 for Kitl. The results indicated that SATB2 could negatively regulate the expression of Kitl in the developing cerebral cortex through a conserved regulatory element in intron 1 of Kitl.
Satb2-targeted genes may be involved in axonogenesis in the developing cerebral cortex
We used the R package “ontologyPlot” to generate the ontology diagram of the enriched descendant GO terms to “axonogenesis”. Satb2-targeted axonogenesis genes mainly enriched on axon extension and axon guidance processes (Fig. 4a). We detected several axon guidance molecules dysregulated in Satb2−/− mice compared with Satb2+/+ group (Fig. 4b) and mRNA level changes of these axon guidance molecules were verified by RT-qPCR (Fig. 4c). Expression of Plxnd1, Ntng1, Efnb2, and Ephb1 were significantly increased, while expression of Plxna2, Epha3, Plxna4, Unc5c, and Flrt2 were significantly decreased. Proper axon guidance is essential in neuron migration and projection to their targets. Disrupted axon guidance cues might lead to attractive or repulsive environmental disorganized, which growth cone of axons can sense, thus causing misprojection of neurons, so we inferred that Satb2-targeted axonogenesis genes may have influences on axon growth.

a Satb2-targeted axonogenesis genes mainly enriched on axon extension and axon guidance included in axonogenesis. The darker yellow color represents the lower p-value in the GO enrichment analysis. b Differentially expressed axon guidance molecules in Satb2-/- mouse cortex. Unpaired t-test, ∗∗p < 0.05, n = 3, error bars represent standard error of mean. c RT-qPCR verification of genes in b in the cerebral cortex of E17.5 Satb2+/+ and Satb2-/- mice. d Increased expression of Ntng1 in Satb2-/- mouse cortex. e SATB2 binds to conserved intronic sequences of Ntng1 gene. Red arrows indicate a binding peak. f Luciferase reporter assay showed regulation of Ntng1-S-pEZX by SATB2. **p < 0.01, ***p < 0.001, n = 3.
Among the upregulated axon guidance genes, we identified a new Satb2 target, Ntng1, encoding Netrin-G1 (Fig. 4d, p-value < 0.01). Netrin-G1 is a vertebrate-specific guidance molecule, mediating several aspects of neural-circuit formation, such as neurite elongation and laminar organization of dendrites in mice.32 ChIP-seq analysis identified a strong and specific binding of SATB2 to highly conserved intron 4 of the Ntng1 gene (Fig. 4e). Luciferase activity significantly decreased when co-transfected SATB2 with Ntng1-S-pEZX (Fig. 4f). When co-transfected mutant SATB2 R239X with Ntng1-S-pEZX, the activity of luciferase increased compared with wild-type SATB2 group. The result indicates that Satb2 can directly and negatively regulate the expression of Ntng1 in the developing cerebral cortex through a conserved regulatory element in intron 1 of Ntng1.
Satb2-targeted genes may be involved in synapse formation in the developing cerebral cortex
To further understand synaptic functions of Satb2 in cerebral development, we next focus on synaptic genes from SynaptomeDB.33 Comparing DEGs at E17.5 and P0 stages with ChIP-targeted genes, we found 168 targeted synaptic genes of Satb2 (Fig. 5a and Table S7). GO analysis showed the 168 synaptic genes were enriched in synapse organization, regulation of synaptic structure or activity or membrane, synaptic vesicle and synaptic transmission (Fig. 5b). Over-represented KEGG pathways included glutamatergic synapse, synaptic vesicle cycle and arginine biosynthesis.

a Venn diagram shows 168 genes overlapping between Satb2-binding synaptic genes and E17.5_DEG synaptic genes or P0_DEG synaptic genes. b Gene Ontology or KEGG pathway analysis of Satb2-targeted synaptic genes. c Localization of Satb2-targeted synaptic genes in the synapses and expression changes in different stages. Genes linked by pink or blue lines are simultaneously upregulated or downregulated in Satb2 knockout mice at E17.5, P0 and adult stages. d Venn diagram shows gene overlap between Satb2-binding synaptic genes and E17.5_P0 DEG synaptic genes or adult_DEG synaptic genes. e Gene ontology enrichment of genes in d. f RT-qPCR of Cdh13 in the cerebral cortex of E17.5 Satb2+/+ and Satb2−/− mice. Unpaired t-test, ∗∗p < 0.05, n = 3. g SATB2 binds to conserved promoter sequences of Cdh13 gene. Red arrows indicate binding peak. h Luciferase reporter assay showed regulation of Cdh13-S-pEZX by SATB2. **p < 0.01, ***p < 0.001, n = 3.
To find out whether Satb2 regulates different groups of synaptic genes during developmental stages, we re-analyzed RNA-seq data of Satb2-mutant cortices from adult mice published previously.20 Comparing of DEGs detected from E17.5, P0 and adult samples revealed that only 72 synaptic genes overlapped between the early developmental stage and the adult stage while most differential expressed synaptic genes were stage specific (Fig. S3). Most of the synaptic genes were differentially expressed in P0 stage, and only a small number of genes were simultaneously upregulated or downregulated in Satb2 knockout mouse at E17.5, P0 and adult stages (genes linked by pink or blue lines in different developmental stages) (Fig. 5c). The patterns were similar among presynaptic, postsynaptic, preactivezone and vesicles genes. When considering Satb2 ChIP-targeted synaptic genes, we found 25 are consistently regulated among different stages and 143 genes are E17.5/P0 specific (Fig. 5d), including Cdh10, Cdh13, Cacna2d1, Cntnap2, Epha4 and Gabra1 (Labeled in Fig. 3b). E17.5/P0-specific Satb2-targeted synaptic genes mainly enriched on calmodulin binding, cell adhesion molecule binding and actin filament binding (Fig. 5e), which were essential for initiation of synaptic formation.34 Adult-specific synaptic genes mainly enriched on enzyme activator and postsynaptic neurotransmitter receptor activity and these processes were important for synapse specification and synaptic plasticity. These results demonstrated that Satb2 can also regulate synaptic formation at early developmental stage by targeting different sets of genes compared to adult stage.
Cadherin-13 (CDH13) is critical in the regulation of cell migration,35 neurite outgrowth and synapse formation in neurons with monoaminergic or GABAergic specification.36,37,38 Cdh13 was an E17.5/P0-specific Satb2-targeted synaptic gene and showed increased expression in Satb2−/− mouse cerebral cortex compared with Satb2+/+ group from RNA-seq and RT-qPCR (Fig. 5f, p-value < 0.01). ChIP-seq analysis identified a strong and specific binding of SATB2 to the highly conserved promoter region of the Cdh13 gene (Fig. 5g). Luciferase activity significantly decreased when co-transfected SATB2 with Cdh13-S-pEZX (Fig. 5h). However, when co-transfected mutant SATB2 R239X with Cdh13-S-pEZX, activity of luciferase increased compared with SATB2 and Cdh13-S-pEZX group. These results showed that Satb2 can directly and negatively regulate expression of Cdh13 in the developing cerebral cortex through the conserved element in Cdh13 promoter region.
Satb2 positively regulates Mef2c expression
To further investigate mechanisms of Satb2 in the regulation of synaptic function in the early developmental stage, we analyzed transcription factors among E17.5 DEG genes, Satb2-targeted genes and neurological disease genes. We found 4 transcription factors overlapping among the three dataset, that is Satb2, Mef2c, Zbtb18 and Zeb2 (labeled by red color in Fig. S1A). Mef2c has been demonstrated to be essential for synaptic function and regulate excitatory/inhibitory synapse density predominantly as a cell-autonomous, transcriptional repressor.39 We identified decreased expression of Mef2c in Satb2−/− mouse cerebral cortex from RNA-seq data and verified by RT-qPCR (Fig. 6a, p-value < 0.01). Western blot and relative quantification of bands also help us verified decreased expression of MEF2C at protein level in Satb2−/− mouse cerebral cortex (Fig. 6b, c). Immunofluorescence staining of SATB2 and MEF2C showed that MEF2C had a high expression in the cerebral cortex especially in layer 2/3/4 in wild-type mouse brain and expression level of MEF2C was reduced significantly in the cerebral cortex of Satb2−/− mouse (Fig. 6d). Co-staining of SATB2 and MEF2C also showed both SATB2 and MEF2C localized in nucleus, and there were cells showed co-localization of SATB2 and MEF2C in layer 2/3 in E17.5 Satb2+/+ cerebral cortex (Fig. S4). These results showed that knockout of Satb2 resulted in reduced expression of MEF2C.

a RT-qPCR of Mef2c in the cerebral cortex of E17.5 Satb2+/+ and Satb2−/− mice. Unpaired t-test, ∗∗p < 0.05, n = 3, error bars represent standard error of mean. b Representative western blot images of MEF2C protein in the cerebral cortex of E17.5 Satb2+/+ and Satb2−/− mice. c Relative quantitative analysis of western blot bands. Unpaired t-test, ∗∗p < 0.05, n = 4. d Representative images of SATB2 and MEF2C immunostaining in E17.5 Satb2+/+ and Satb2−/− cortex. e SATB2 binds to conserved intronic sequences of Mef2C gene. Red arrows indicate two binding peaks. f Luciferase reporter assay showed regulation of Mef2c-S1-pEZX or Mef2c-S2-pEZX by SATB2. **p < 0.01, ***p < 0.001, n = 3. g Venn diagram shows that 28 Satb2-targeted genes overlap with the Mef2c direct targeted genes and 345 Satb2-targeted genes overlap with the Mef2c secondary targeted genes.
ChIP-seq analysis identified strong and specific bindings of SATB2 to highly conserved intron 2 regions of the Mef2c gene. Two sequences (Mef2c-S1 and Mef2c-S2) were inserted into the luciferase reporter plasmid to test whether Satb2 directly regulates the detected regions of Mef2c (Fig. 6e). Co-transfection of SATB2 or SATB2 R239X with empty pEZX-FR01 plasmids slightly increased luciferase activity. However, luciferase activity significantly increased when co-transfection SATB2 with Mef2c-S1-pEZX or Mef2c-S2-pEZX (Fig. 6f). Moreover, the activity of luciferase didn’t increase when co-transfection mutant SATB2 R239X with Mef2c-S1-pEZX or Mef2c-S2-pEZX. The result indicated that mutant SATB2 R239X lead to a weakened regulatory activity. We also observed similar pattern of luciferase activity in Mef2c-S2- pEZX group (Fig. 6f). These results suggested that Satb2 can regulate the conserved intron regions of Mef2c and Satb2 likely directly regulate the expression of Mef2c in the developing cerebral cortex.
To find out whether Satb2 can regulate the expression of synaptic function-related genes through Mef2c, we compared Satb2-targeted genes and Mef2c-targeted genes from the study of Allaway et al.40 We found 28 genes overlap between Satb2-targeted genes and Mef2c_direct genes in the early developmental stage, indicating that expression of the 28 genes can be regulated by Satb2 directly or by Mef2c which is regulated by Satb2 (Fig. 6g). Among the 28 genes, Arhgef28, Dtna, Cdh7, Cacna2d1 and Caln1 have been reported to be associated with synaptic functions.41,42,43,44,45 Additionally, we found 332 genes overlapping between Satb2-targeted genes and Mef2c_2nd genes. We inferred that these genes were regulated by Satb2 directly or by other transcription factors that Mef2c regulates. By integrating of Satb2 ChIP-seq data and RNA-seq data of adult Satb2 and Mef2c knockout cortical cortex,20,39 we identified 206 genes were directly regulated by Satb2 or Mef2c in the adult stage (Fig. S5A). The 206 genes are enriched mainly on synapse organization, synapse assembly, forebrain development and cell junction assembly (Fig. S5B). These results suggested that the numbers of synaptic genes regulated by Satb2 or Mef2c increased when synapse and cognitive function getting more important and complex from early developmental stage to adult stage.
Impaired spatial learning and memory in Satb2 +/− mice
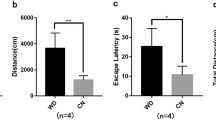
Since Satb2−/− mice die immediately after birth,2 we performed behavioral tests with Satb2+/+ and Satb2+/− mice. We recorded the weights of Satb2+/+ and Satb2+/− mice after birth and found Satb2+/− mice were significantly lighter than Satb2+/+ mice since D12 (Fig. S6A). Open field test, passive avoidance test, elevated plus-maze test and novel object recognition test showed no differences between Satb2+/+ and Satb2+/− mice (data not shown). In the Morris water maze test (MWM), Satb2+/− mice exhibited a longer latency in finding the platform than Satb2+/+ mice during the learning phase (Fig. S6B). The result indicated that the Satb2+/− mice may have impaired learning capacity during the consecutive 4-days training phase of the MWM task than Satb2+/+ mice. During the memory test, the number of platform crossings was lower in the Satb2+/− mice compared to Satb2+/+ mice (Fig. S6C). The distance or time to the platform showed no obvious difference between Satb2+/+ and Satb2+/− mice (Fig. S6D, E). Thus, we thought Satb2+/− mice showed impaired spatial learning and memory, further strengthening that Satb2 regulates functions of synapse.
Discussion
Here, we identified downstream targeted genes of Satb2 and showed that Satb2 directly regulated genes important in axonogenesis, synaptic organization and cell adhesion during the early development of the cerebral cortex.
Satb2 has been demonstrated to regulate many biological processes as a transcription factor. Integration of RNA-seq and ChIP-seq data helped us identified 1363 targeted genes of Satb2 with different temporal expression patterns (Fig. 2). Interestingly, 73 Satb2-targeted genes are associated with neurological disease, which could explain the mental abnormalities in SAS patients.6 The growth of axons towards their targets is critical during the establishment of neuronal connections under the regulation of axon guidance ligands and receptors.46 Satb2 and Ctip2 directly repress the expression of two Netrin1 receptors- DCC and Unc5C to control the interhemispherical projection in a subset of early born, deep layer callosal neurons.13 Satb2 also has been reported to regulate EphA7 to control soma spacing and self-avoidance of cortical pyramidal neurons.14 In this study, we found eight additional downstream axon guidance genes regulated by Satb2, that is Plxna2, Plxna4, Plxnd1, Ntng1, Epha3, Ephb1, Efnb2, and Flrt2 (Fig. 4c). Plxna2, Plxna4 and Plxnd1 code cell surface receptors for class 3 semaphorins (SEMA3A/3C/3E), SEMA4A and SEMA6A, and have been demonstrated to regulate neuronal migration or ensure specificity of synapse formation.47,48,49 PLXNA2 has recently been shown as a candidate gene in autism spectrum disorder and intellectual disability.50,51 Epha3, Ephb1, Efnb2 code ligand or receptors of the ephrins family, which inhibit axon growth by mediating growth cone collapse through regulation of Ras and Rho GTPases.52 Epha3 also promotes axon growth through RhoA signaling and NMIIA in developing neurons.53 We provided experimental evidence of Ntng1 to be a new axon guidance molecule regulated by Satb2 (Fig. 4d–f). Therefore, these evidences provided new insights into the complexity of Satb2 in regulating axonogenesis.
During neuronal development, axons and dendrites establish initial synaptic contacts mostly during development and the early postnatal period, although synapse formation continues throughout life at a lower rate.34 It has been revealed that Satb2 has different roles as a cell fate and neuron projection determinant at the neonatal stage while a regulator of synaptic plasticity/physiology at the adult stage.20 Our study identified 168 Satb2-targeted synaptic genes and 143 early-stage-specific Satb2-targeted synaptic genes, suggesting that Satb2 can also regulate synaptic formation at the early developmental stage (Fig. 5). 168 synaptic genes enriched on synapse organization, regulation of synaptic structure or activity or membrane, synaptic vesicle and synaptic transmission. Cdh13, one of the 143 early-stage-specific Satb2-targeted synaptic genes, had an increased expression in the absence of Satb2. Several studies revealed Cdh13 variants associated with memory impairment54 and hyperactivity/impulsivity, specifically during childhood and adolescents.55 Luciferase reporter assay provides evidence that Satb2 can directly regulate the expression of Cdh13 (Fig. 5h). Satb2 was thought as a cell fate and neuron projection determinant at neonatal stage and regulator of synaptic plasticity/physiology at the adult stage. Our study firstly demonstrated the function of Satb2 in synaptic formation at the embryonic stage; our findings enriched the functions of Satb2 in the early development stage.
Myocyte enhancer factor 2 C (MEF2C) is crucial for programming early neuronal differentiation and regulation of dendritic spine density, synapse development, synapse number, and AMPA-mediated postsynaptic strength.39,56,57 We provided experimental evidence that Mef2c was downregulated in Satb2−/− mice (Fig. 6a–d). Our study firstly suggests Mef2c as a downstream effector of Satb2 to regulate synaptic transmission in the developing cerebral cortex (Fig. 6e, f). Compared analysis of targeted genes of Satb2 and Mef2c showed that several targets of Mef2c were regulated directly or indirectly by Satb2 (Fig. 6f), indicating a complex regulatory network of Satb2.
Satb2 cKO mice showed hyperactivity, increased impulsivity, abnormal social novelty, and impaired spatial learning and memory.58 MWM test of global Satb2+/− mice showed impaired spatial learning and memory (Fig. S6), further strengthening that Satb2 regulates synapse functions. Considering SATB2 heterozygous variations in humans cause disease and SATB2 expression in mice and human has similar patterns during prenatal cortex development, our results showed that Satb2 heterozygous knockout mice had similar phenotypes and were more consistent with human disease model.
Conclusion
Our study has increased the knowledge of the potential target genes of Satb2 in the developing cerebral cortex and these target genes may be involved in axonogenesis, synapse formation and synaptic plasticity. We also explored the direct regulation of Satb2 in 4 downstream effectors (Kitl, Ntng1, Cdh13, and Mef2c) although these still need further investigation. Our study demonstrates complicated molecular regulatory mechanism of Satb2 in axonogenesis and synaptic function in the early developmental stage and provides new insights to elucidate the pathogen of SAS.
Data availability
RNAseq and ChIP-seq data that support the findings of this study have been deposited to GEO database with the accession number GSE201562.
References
Britanova, O., Akopov, S., Lukyanov, S., Gruss, P. & Tarabykin, V. Novel transcription factor Satb2 interacts with matrix attachment region DNA elements in a tissue-specific manner and demonstrates cell-type-dependent expression in the developing mouse CNS. Eur. J. Neurosci. 21, 658–668 (2005).
Dobreva, G. et al. SATB2 is a multifunctional determinant of craniofacial patterning and osteoblast differentiation. Cell 125, 971–986 (2006).
Alcamo, E. A. et al. Satb2 regulates callosal projection neuron identity in the developing cerebral cortex. Neuron 57, 364–377 (2008).
Britanova, O. et al. Satb2 is a postmitotic determinant for upper-layer neuron specification in the neocortex. Neuron 57, 378–392 (2008).
Li Y, et al. Satb2 ablation impairs hippocampus-based long-term spatial memory and short-term working memory and immediate early genes (IEGs)-mediated hippocampal synaptic plasticity. Mol. Neurobiol. https://doi.org/10.1007/s12035-017-0531-5 (2017).
Zarate, Y. A. & Fish, J. L. SATB2-associated syndrome: mechanisms, phenotype, and practical recommendations. Am. J. Med Genet. A 173, 327–337 (2017).
Lewis, H. et al. Epilepsy and electroencephalographic abnormalities in SATB2-associated syndrome. Pediatr. Neurol. 112, 94–100 (2020).
Leone, D. P. et al. Satb2 regulates the differentiation of both callosal and subcerebral projection neurons in the developing cerebral cortex. Cereb. Cortex 25, 3406–3419 (2015).
Hall, A. & Lalli, G. Rho and Ras GTPases in axon growth, guidance, and branching. Cold Spring Harb. Perspect. Biol. 2, a001818 (2010).
Tessier-Lavigne, M. & Goodman, C. S. The molecular biology of axon guidance. Science 274, 1123–1133 (1996).
Kamiguchi, H. The mechanism of axon growth: what we have learned from the cell adhesion molecule L1. Mol. Neurobiol. 28, 219–228 (2003).
Baranek, C. et al. Protooncogene Ski cooperates with the chromatin-remodeling factor Satb2 in specifying callosal neurons. Proc. Natl Acad. Sci. USA 109, 3546–3551 (2012).
Srivatsa, S. et al. Unc5C and DCC act downstream of Ctip2 and Satb2 and contribute to corpus callosum formation. Nat. Commun. 5, 3708 (2014).
He, C. -H. et al. Satb2 regulates EphA7 to control soma spacing and self-avoidance of cortical pyramidal neurons. Cereb. Cortex 32, 2321–2331 (2021).
Defelipe, J. The evolution of the brain, the human nature of cortical circuits, and intellectual creativity. Front. Neuroanat. 5, 29 (2011).
Takeuchi, T., Duszkiewicz, A. J. & Morris, R. G. M. The synaptic plasticity and memory hypothesis: encoding, storage and persistence. Philos. Trans. R. Soc. Lond. B Biol. Sci. 369, 20130288 (2014).
Ewels, P. A. et al. The nf-core framework for community-curated bioinformatics pipelines. Nat. Biotechnol. 38, 276–278 (2020).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
McKenna, W. L. et al. Mutual regulation between Satb2 and Fezf2 promotes subcerebral projection neuron identity in the developing cerebral cortex. Proc. Natl Acad. Sci. USA 112, 11702–11707 (2015).
Cera, I. et al. Genes encoding SATB2-interacting proteins in adult cerebral cortex contribute to human cognitive ability. PLoS Genet. 15, e1007890 (2019).
Li, H. & Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 25, 1754–1760 (2009).
Zhang, Y. et al. Model-based analysis of ChIP-Seq (MACS). Genome Biol. 9, R137 (2008).
Yu, G., Wang, L.-G. & He, Q.-Y. ChIPseeker: an R/Bioconductor package for ChIP peak annotation, comparison and visualization. Bioinformatics 31, 2382–2383 (2015).
Wu, T. et al. clusterProfiler 4.0: a universal enrichment tool for interpreting omics data. Innovation (N. Y) 2, 100141 (2021).
Luo, W. & Brouwer, C. Pathview: an R/Bioconductor package for pathway-based data integration and visualization. Bioinformatics 29, 1830–1831 (2013).
Hu, H. et al. AnimalTFDB 3.0: a comprehensive resource for annotation and prediction of animal transcription factors. Nucleic Acids Res. 47, D33–D38 (2019).
Weyn-Vanhentenryck, S. M. et al. Precise temporal regulation of alternative splicing during neural development. Nat. Commun. 9, 2189 (2018).
Kumar, L. & E Futschik, M. Mfuzz: a software package for soft clustering of microarray data. Bioinformation 2, 5–7 (2007).
Jin, K., Mao, X. O., Sun, Y., Xie, L. & Greenberg, D. A. Stem cell factor stimulates neurogenesis in vitro and in vivo. J. Clin. Invest. 110, 311–319 (2002).
Erlandsson, A., Larsson, J. & Forsberg-Nilsson, K. Stem cell factor is a chemoattractant and a survival factor for CNS stem cells. Exp. Cell Res. 301, 201–210 (2004).
Leoyklang, P. et al. Heterozygous nonsense mutation SATB2 associated with cleft palate, osteoporosis, and cognitive defects. Hum. Mutat. 28, 732–738 (2007).
Yin, Y., Miner, J. H. & Sanes, J. R. Laminets: laminin- and netrin-related genes expressed in distinct neuronal subsets. Mol. Cell Neurosci. 19, 344–358 (2002).
Pirooznia, M. et al. SynaptomeDB: an ontology-based knowledgebase for synaptic genes. Bioinformatics 28, 897–899 (2012).
Südhof, T. C. Towards an understanding of synapse formation. Neuron 100, 276–293 (2018).
Philippova, M., Ivanov, D., Tkachuk, V., Erne, P. & Resink, T. J. Polarisation of T-cadherin to the leading edge of migrating vascular cells in vitro: a function in vascular cell motility? Histochem. Cell Biol. 120, 353–360 (2003).
Paradis, S. et al. An RNAi-based approach identifies molecules required for glutamatergic and GABAergic synapse development. Neuron 53, 217–232 (2007).
Rivero, O. et al. Cadherin-13, a risk gene for ADHD and comorbid disorders, impacts GABAergic function in hippocampus and cognition. Transl. Psychiatry 5, e655 (2015).
Drgonova, J. et al. Cadherin 13: human cis-regulation and selectively-altered addiction phenotypes and cerebral cortical dopamine in knockout mice. Mol. Med. 22, 537–547 (2016).
Harrington, A. J. et al. MEF2C regulates cortical inhibitory and excitatory synapses and behaviors relevant to neurodevelopmental disorders. Elife 5, e20059 (2016).
Allaway, K. C. et al. Genetic and epigenetic coordination of cortical interneuron development. Nature 597, 693–697 (2021).
Kuwako, K.-I., Nishimoto, Y., Kawase, S., Okano, H. J. & Okano, H. Cadherin-7 regulates mossy fiber connectivity in the cerebellum. Cell Rep. 9, 311–323 (2014).
Föcking, M. et al. Proteomic analysis of the postsynaptic density implicates synaptic function and energy pathways in bipolar disorder. Transl. Psychiatry 6, e959 (2016).
Beeson, K. A., Beeson, R., Westbrook, G. L. & Schnell, E. α2δ-2 protein controls structure and function at the cerebellar climbing fiber synapse. J. Neurosci. 40, 2403–2415 (2020).
Di Gregorio, S. E., Volkening, K., Strong, M. J. & Duennwald, M. L. Inclusion formation and toxicity of the ALS protein RGNEF and its association with the microtubule network. Int. J. Mol. Sci. 21, E5597 (2020).
Reshetnikov, V. V. et al. Genes associated with cognitive performance in the Morris water maze: an RNA-seq study. Sci. Rep. 10, 22078 (2020).
Russell, S. A. & Bashaw, G. J. Axon guidance pathways and the control of gene expression. Dev. Dyn. 247, 571–580 (2018).
Sakurai, A. et al. Semaphorin 3E initiates antiangiogenic signaling through plexin D1 by regulating Arf6 and R-Ras. Mol. Cell Biol. 30, 3086–3098 (2010).
Hatanaka, Y. et al. Semaphorin 6A-plexin A2/A4 interactions with radial glia regulate migration termination of superficial layer cortical neurons. iScience 21, 359–374 (2019).
Limoni, G., Murthy, S., Jabaudon, D., Dayer, A. & Niquille, M. PlexinA4-Semaphorin3A-mediated crosstalk between main cortical interneuron classes is required for superficial interneuron lamination. Cell Rep. 34, 108644 (2021).
Altuame, F. D. et al. PLXNA2 as a candidate gene in patients with intellectual disability. Am. J. Med. Genet. A 185, 3859–3865 (2021).
Pijuan, J. et al. PLXNA2 and LRRC40 as candidate genes in autism spectrum disorder. Autism Res. 14, 1088–1100 (2021).
Noren, N. K. & Pasquale, E. B. Eph receptor-ephrin bidirectional signals that target Ras and Rho proteins. Cell Signal 16, 655–666 (2004).
Javier-Torrent, M. et al. Presenilin/γ-secretase-dependent EphA3 processing mediates axon elongation through non-muscle myosin IIA. Elife 8, e43646 (2019).
Salatino-Oliveira, A. et al. Catechol-O-methyltransferase valine158methionine polymorphism moderates methylphenidate effects on oppositional symptoms in boys with attention-deficit/hyperactivity disorder. Biol. Psychiatry 70, 216–221 (2011).
Salatino-Oliveira, A. et al. Cadherin-13 gene is associated with hyperactive/impulsive symptoms in attention/deficit hyperactivity disorder. Am. J. Med. Genet. B Neuropsychiatr. Genet. 168B, 162–169 (2015).
Rajkovich, K. E. et al. Experience-dependent and differential regulation of local and long-range excitatory neocortical circuits by postsynaptic Mef2c. Neuron 93, 48–56 (2017).
Assali, A., Harrington, A. J. & Cowan, C. W. Emerging roles for MEF2 in brain development and mental disorders. Curr. Opin. Neurobiol. 59, 49–58 (2019).
Zhang, Q., Huang, Y., Zhang, L., Ding, Y.-Q. & Song, N.-N. Loss of Satb2 in the cortex and hippocampus leads to abnormal behaviors in mice. Front. Mol. Neurosci. 12, 33 (2019).
Acknowledgements
We are very grateful to Dr. Zilong Qiu and Dr. Man Xiong for their comments and suggestions during the study. We are also grateful to the animal co-facility center of Children’s Hospital of Fudan University and our genetic laboratory teams who contributed to this study.
Funding
This study is funded by the National Natural Science Foundation of China (81471483, 81974237).
Author information
Authors and Affiliations
Contributions
H.W. designed the study and revised the manuscript; Q.G. performed experiment, collected and interpreted the data, drafted the initial manuscript; Y.W. analyzed the RNA-seq and ChIP-seq data; Y.Q., X.C. and X.L. involved experiment, X.D. and H.C. involved data analysis, Q.W., Y.J., S.Y., J.Z., and S.S. involved animal experiment; B.W. and W.Z. revised the manuscript; and all authors read and approved the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Guo, Q., Wang, Y., Wang, Q. et al. In the developing cerebral cortex: axonogenesis, synapse formation, and synaptic plasticity are regulated by SATB2 target genes. Pediatr Res 93, 1519–1527 (2023). https://doi.org/10.1038/s41390-022-02260-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41390-022-02260-z
This article is cited by
-
Sex differences in brain protein expression and disease
Nature Medicine (2023)
