
Abstract
Humans greatly differ in how they cope with stress, a natural behavior learnt through negative reinforcement. Some individuals engage in displacement activities, others in exercise or comfort eating, and others still in alcohol use. Across species, adjunctive behaviors, such as polydipsic drinking, are used as a form of displacement activity that reduces stress. Some individuals, in particular those that use alcohol to self-medicate, tend to lose control over such coping behaviors, which become excessive and compulsive. However, the psychological and neural mechanisms underlying this individual vulnerability have not been elucidated. Here we tested the hypothesis that the development of compulsive adjunctive behaviors stems from the functional engagement of the dorsolateral striatum (DLS) dopamine-dependent habit system after a prolonged history of adjunctive responding. We measured in longitudinal studies in male Sprague Dawley rats the sensitivity of early established vs compulsive polydipsic water or alcohol drinking to a bilateral infusion into the anterior DLS (aDLS) of the dopamine receptor antagonist α-flupentixol. While most rats acquired a polydipsic drinking response with water, others only did so with alcohol. Whether drinking water or alcohol, the acquisition of this coping response was insensitive to aDLS dopamine receptor blockade. In contrast, after prolonged experience, adjunctive drinking became dependent on aDLS dopamine at a time when it was compulsive in vulnerable individuals. These data suggest that habits may develop out of negative reinforcement and that the engagement of their underlying striatal system is necessary for the manifestation of compulsive adjunctive behaviors.
Similar content being viewed by others


Introduction
When facing the distress generated by a challenging, emotionally taxing, aversive situation, individuals greatly differ in the emotion regulation strategy they use, which is influenced by situational demands [1,2,3,4,5,6]. Emotion regulation and associated coping strategies [7], such as exercise, comfort eating, shopping, or displacement behavior [8, 9], all of which are driven by the negative reinforcing effects of stress reduction, have long been suggested to be a prerequisite for adaptive functioning, the promotion of resilience and well-being [10, 11].
Several species cope with stress using a form of displacement called adjunctive behavior [12, 13]. One such adjunctive anxiolytic response, schedule-induced polydipsia (SIP) [12,13,14,15], manifests itself as polydipsic water intake in the face of intermittent food delivery in food-restricted animals [16,17,18]. At the population level, non-regulatory polydipsic drinking develops over a week and remains stable for long periods of time during which it selectively decreases the levels of stress-related hormones that had been increased by the associated intermittent food delivery [14, 15, 19,20,21].
However, some humans [22,23,24,25,26,27,28,29] and individuals of other species [30] characterized, for instance by a high impulsivity trait [31], lose control over these coping strategies which become excessive and promote the development of compulsive disorders such as obsessive compulsive and substance use disorders.
During emotion regulation challenges under circumstances often perceived as overwhelming, such as were posed by the COVID-19 pandemic [32,33,34,35], some individuals resort to alternative means to cope with stress, such as drinking alcohol [36,37,38,39,40] or use of other drugs [41]. This in turn is associated with a greater vulnerability to develop compulsivity [40, 42,43,44,45,46,47,48,49,50]. Across species the individual tendency to rely on the anxiolytic effects of alcohol to cope with stress [42, 50, 51], including that induced by the SIP procedure [52], has been associated with an increased vulnerability to switch from controlled to compulsive alcohol use, a key feature of alcohol use disorder (AUD) [42, 49, 50, 52,53,54].
However, the psychological and neural basis of the individual vulnerability to lose control over coping behavior, whether involving alcohol use or not, and the ensuing development of compulsivity are unclear.
Increasing evidence suggests that the transition from controlled, goal-directed, behavior to compulsion, including the compulsive seeking and drinking of alcohol [55, 56] and the development of compulsive adjunctive polydipsic drinking, is dependent on a shift in the locus of control over behavior from the ventral to the dorsolateral striatum (DLS)-dependent habit system [57]. Thus, while the reinforcing properties of alcohol, mediated by the mesolimbic dopamine (DA), system support recreational alcohol use [58, 59], it is the engagement of anterior DLS (aDLS) DA dependent alcohol seeking habits that promotes the transition to compulsive alcohol seeking and drinking [55, 56, 59]. Similarly, the development of adjunctive polydipsic water drinking behavior, but not its compulsive manifestation, is dependent on the mesolimbic system [60, 61]. In contrast, well-established excessive adjunctive water drinking (hyperdipsia) in vulnerable individuals has been associated with an increased in spine density in DLS medium spiny neurons [62].
Here we tested the hypothesis that the development of compulsive coping behavior, manifested as excessive polydipsic drinking, whether of alcohol or water, is associated with the functional engagement of, and an increased reliance on, the DLS DA-dependent habit system.
To test this hypothesis, we assessed the sensitivity of the polydipsic water or alcohol drinking response of each individual in large cohorts of outbred rats to bilateral aDLS infusions of the DA receptor antagonist α-flupentixol [52, 56, 63] over the course of the acquisition of each coping behavior and the subsequent transition to compulsivity.
Methods and materials
Subjects
One hundred and forty-one male Sprague Dawley rats (Charles River, UK), weighing 300–350 g at the start of the experiments, were used in this study as described in detail in the SOM. All experimental protocols were conducted under the project license 70/8072 held by David Belin in accordance with the regulatory requirement of the UK Animals (Scientific Procedures) Act 1986, amendment regulations 2012, following ethical review by the University of Cambridge Animal Welfare and Ethical Review Body (AWERB).
Experimental procedures
The series of experiments are schematically summarized in Fig. 1.

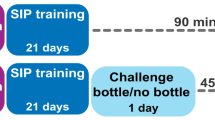
Timeline of the four experiments carried out on independent cohorts of male Sprague Dawley rats. Experiment 1: Following a week of habituation to the animal facility, thirty-six rats received bilateral cannulation of their anterior dorsolateral striatum (aDLS). A week later, rats were food restricted to 85% of their theoretical free-feeding weight and trained in a schedule-induced polydipsia (SIP) procedure with water (SIPw). The reliance of the acquisition of adjunctive water drinking on aDLS dopaminergic mechanisms was assessed after 5 SIPw sessions as the sensitivity of drinking behavior to bilateral infusion of the dopamine (DA) receptor antagonist α-flupentixol (α-flu, 0, 6, 12 μg/side, between-subject design) into the aDLS (Test 1, α-flu Early SIPw). Experiment 2: Following a week of habituation to the animal facility, forty-eight rats were food restricted for a week before being trained in the SIPw procedure for 20 sessions, e.g., until the establishment of hyperdipsia in vulnerable individuals. Then, rats were implanted with bilateral cannulae targeting the aDLS and, after at least a week, they were re-baselined under SIPw for 10 sessions. Then, the reliance of well-established, compulsive, adjunctive water drinking behavior on aDLS DA was assessed as the sensitivity of water drinking behavior to bilateral infusion of α-flupentixol (0, 6, 12 μg/side, within-subject design) into the aDLS (Test 2, α-flu Late SIPw). Subsequently, water was replaced by 10% alcohol (SIPa) and the reliance of early and well-established adjunctive alcohol drinking on aDLS DA was measured after 3 (Test 3, α-flu Early SIPa) or 20 (Test 4, α-flu Late SIPa) SIPa sessions, respectively. Experiment 3: In order to test the stability of water intake levels once adjunctive water drinking has been established, ten rats were food restricted after a week of habituation to the colony, and then trained in the SIP procedure for 40 sessions, a period of training similar to that used in experiment 2, but with water as the only available solution throughout. Experiment 4: In order to establish the anxiolytic properties of polydipsic drinking under SIP at the population level but also, specifically in individuals that relied on alcohol to engage in a coping strategy, forty-seven rats were habituated to the animal facility and food restricted for a week before their anxiety levels were assessed as the percentage of time spent in the open arms of an elevated plus maze (EPM) prior to (naïve, EPM 1), and immediately following a session after at least 20 days of training under SIPw (EPM 2) or SIPa (EPM 3).W: week; s: session.
Experiments 1 and 2 aimed to establish the reliance of coping behavior on a circuit that involves aDLS DA-dependent mechanisms (referred to subsequently as aDLS DA), an established assay that reveals the engagement of habitual control over behavior reinforced by addictive drugs, including alcohol [55, 56, 63]. Thus inactivation of, or DA receptor blockade in, the aDLS does not influence early performance of alcohol, cocaine or heroin seeking behavior [63,64,65,66,67], which instead is mediated by the posterior dorsomedial striatum (pDMS)-dependent goal-directed system [63, 64, 67]. In contrast, the later emerging habitual responding for these drugs becomes sensitive to the same aDLS manipulations that had no effect on early performance [55, 56, 63, 64, 67,68,69]. These data reveal a double dissociation in the control over behavior by the aDLS- and pDMS-dependent corticostriatal networks mediating habitual and goal-directed control over behavior, respectively [63, 67, 70].
As detailed in Fig. 1 and the SOM, the first experiment aimed to determine the involvement of aDLS DA in the acquisition of a coping adjunctive water drinking response (SIPw) (Test 1, Effect of α-flu on Early SIPw).
The second experiment aimed to test the reliance on aDLS DA of well-established adjunctive water drinking (Test 2, Effect of α-flu on Late SIPw) vs that of early (Test 3, Effect of α-flu on Early SIPa) and well-established adjunctive alcohol drinking (SIPa) (Test 4, Effect of α-flu on Late SIPa), as previously described [52].
The third experiment aimed to test the stability of polydipsic water intake levels over a period of training in the SIP procedure similar to that of rats in experiment 2.
The fourth experiment aimed to test the anxiolytic nature of polydipsic alcohol drinking at the population level but also, specifically in individuals that relied on alcohol to engage in a coping strategy.
Schedule-induced polydipsia (SIP)
SIP training was carried out as previously described [31, 71, 72] and detailed in the SOM.
The SIP procedure was based on a fixed-time 60 s (FT-60 s) schedule of food delivery, previously shown to induce polydipsic drinking with robust and persistent individual differences in the tendency to develop excessive, compulsive manifestations of this adjunctive behavior [31, 52, 72,73,74]. Individuals having an average water consumption over the last 3 days of training in the upper and lower quartiles of the population were considered as high (HD) and low drinkers (LD), respectively, as previously described [31, 52, 72]. HD rats are therefore individuals who acquire a coping response with water (e.g., water copers) and tend, for some of them, to lose control over it, thereby expressing hyperdipsia. LD rats, on the other hand do not readily acquire a coping response with water. However, fifty to sixty percent of them drink alcohol to do so, these are alcohol copers [52].
Twenty-four hours after the last SIPw session, water was replaced by 10% alcohol and rats were trained for twenty 60 min SIPa sessions. The total amount of alcohol consumed (mL) was calculated daily as the difference between the weights of the bottle before and after the session.
Drugs
The alcohol and DA receptor antagonist α-flupentixol solutions were prepared as previously described [52, 65] and detailed in the SOM.
Surgery: aDLS cannulations
Rats received bilateral aDLS implantation of canulae under stereotaxic surgery either before behavioral training (Experiment 1), or after acquisition of SIPw (Experiment 2) under isoflurane anesthesia (O2: 2 L/min, 5% isoflurane for induction and 2% for maintenance), as previously described [56] and detailed in the SOM.
Intra-striatal infusions
The influence of aDLS DA receptor blockade on adjunctive drinking behavior was tested at early and late stage of training for SIPw and SIPa, namely during the acquisition of SIPw (Experiment 1, SIPw session 5 onwards, Test 1 Effect of α-flu on Early SIPw) and SIPa (Experiment 2, SIPa session 3 onwards, Test 3, Effect of α-flu on Early SIPa) and well-established SIPw (Experiment 2, SIPw session 20 onwards, Test 2, Effect of α-flu on Late SIPw) and SIPa (SIPa session 20 onwards, Test 4, Effect of α-flu on Late SIPa).
As described in the SOM, each test session was preceded by intra-aDLS infusions (0.5 μl/side) of α-flupentixol (0, 6, 12 μg/side, made via 28-gauge steel injectors (Plastics One, Roanoke, VA, USA) over 90 s followed by a 60 s period to allow diffusion of the infused drug or vehicle before injectors were removed and obturators were replaced. Test sessions began 5 min later. The effect of aDLS DA receptor blockade on adjunctive drinking was tested on rats from experiment 1 (Test 1) in a between-subject design, and in rats from experiment 2 (Tests 2, 3 and 4) in a counter-balanced order following a Latin-Square design. Each infusion day was followed by two baseline sessions.
Histology
As described in detail in the SOM, at the end of the experiment, rats were euthanized, perfused transcardially with 10% neutral buffered formalin and their brains were extracted before being cryoprotected and then processed into 60 μm coronal sections that were mounted and stained with Cresyl Violet. Cannulae placements in the aDLS were verified using a light microscope by an experimenter blind to the behavioral results.
Elevated plus maze
Anxiety levels were assessed as the percentage of time spent in the open arms of an elevated plus maze (EPM) prior to (naïve), and immediately following a session after at least 20 days of training under SIPw or SIPa, as previously described [31] and detailed in the SOM.
Data and statistical analyses
Data, presented as means ± SEM, individual data points or box plots [medians ± 25% (percentiles) and Min/Max as whiskers], were analyzed with STATISTICA-10 Software (Statsoft, Inc., Tulsa, OK, USA) or Statistical Package for Social Sciences (IBM SPSS, v 26, USA) as described in the SOM.
Behavioral data on the acquisition or maintenance of SIPw or SIPa were analyzed using repeated measures (RM) analyses of variance (ANOVA).
The effect of aDLS DA receptor blockade on adjunctive drinking was analyzed with one way- (Test 1) or RM-ANOVA (Test 2, 3 and 4).
Two-step K-mean cluster analysis [56, 71] was performed to identify groups of individuals whose reliance of adjunctive drinking on aDLS DA differed across tests. We additionally computed an index of differential reliance on aDLS DA of compulsive drinking of alcohol vs water to analyze whether the emerging reliance on aDLS control predicts the individual tendency to rely on alcohol to cope with stress and the ensuing development of compulsive alcohol drinking [52].
Rats were also ranked according to their fluid intake (mL) before receiving α-flupentixol infusions (i.e., after the last three session of SIPw or SIPa) to identify individuals that increased their intake when alcohol was introduced in the SIP procedure.
Changes in anxiety level over the course of exposure to SIP were computed as the relative change (%) in the percentage of time spent in the open arms of the maze from one test to the next. A one-tailed t-test was used to test the hypothesis that the introduction of alcohol resulted in a decrease in anxiety levels specifically in individuals that relied on alcohol to develop a coping response. Pearson’s correlations were used to investigate relationships between the magnitude of polydipsic fluid intake and the level of anxiety.
On confirmation of significant main effects, the effect sizes of which are reported as partial eta squared (ηp2), differences among individual means were further analyzed using post-hoc tests or planned comparisons, as appropriate. Significance was set at α ≤ 0.05.
Results
Individual differences in the reliance on alcohol to engage in a coping strategy
Sixty rats with cannula placements located in the aDLS were included in the final analysis (Fig. 2A, B).

A, B Rats having completed experiments 1 and 2 with cannula tips (orange and violet for experiment 1 and 2, respectively) located in the anterior dorsolateral striatum (aDLS) according to the rat brain atlas (black arrows) [149] as assessed following staining with Cresyl Violet were included in the final analyses. C, D At the population level, and similarly across each independent group for experiment 1, food restricted rats exposed to intermittent food delivery progressively developed adjunctive polydipsic water drinking over five (Experiment 1, n = 22) and twenty (Experiment 2, n = 38) sessions. Experiment was designed to reveal marked individual differences in the tendency to engage in polydipsic water drinking that emerged early on in training. While at the population level individuals developed a polydipsic water drinking response, some individuals, developed an excessive, compulsive polydipsic behavior while others did not engage in a displacement strategy whatsoever. Thus, high drinker (HD) and low drinker rats (LD), selected respectively in the upper and lower quartiles of the population stratified on the average water consumption over the last 3 sessions of SIP with water (Late SIPw, green rectangle), greatly differed in their trajectory of polydipsic water drinking. HD rats developed a compulsive water drinking behavior that reached more than 16.65 ± 1.08 ml/h by the last session, or two times more than the whole population, whereas LD rats maintained throughout a water drinking behavior similar to that associated with their homeostatic need displayed at baseline (B). The introduction of alcohol resulted in significant changes in coping behavior. Thus, HD and LD rats still differed in their level of adjunctive drinking at the beginning of the SIP training with alcohol (Early SIPa, blue rectangle), mostly due to the fact that HD rats, who had developed an hyperdipsia with water persisted following the introduction of alcohol, albeit to a lower level. However, while HD rats maintained overall a steady level of polydipsic alcohol drinking over time, LD rats acquired a coping response with alcohol and eventually developed compulsive polydipsic alcohol drinking so that they no longer differed from HD rats by the end of training (Late SIPa, orange rectangle). E This increase in alcohol drinking shown by LD rats was not attributable to an increase in fluid intake over time since once adjunctive drinking was established in an independent cohort of rats over 20 sessions (Experiment 3, n = 10), polydipsic water drinking remained stable so that by session 40 (S40 dashed-line rectangle, average of sessions 38–40) rats displayed a similar level of fluid intake as they did by the end of the first 20 session period (S20 dashed-line rectangle, average of sessions 18–20). The reliance of early (Tests 1 and 3) and well-established (Tests 2 and 4) polydipsic water or alcohol drinking on aDLS DA was assessed as the sensitivity of drinking behavior to bilateral infusion of α-flupentixol (α-flu) at each time point identified by an arrow. ♯ p < 0.001 group × time interaction; *** p < 0.001, ** p < 0.01, HD different from LD rats; ns: no significant.
Exposure to the SIP procedure resulted, at the population level, in a progressive development of adjunctive drinking behavior, expressed as an increase in water intake in response to the introduction of a FT- 60 s schedule of food delivery over 5 (for Experiment 1) or 20 sessions (for Experiment 2) [main effect of time: F2.7,55.8 = 5.46, p = 0.003, ηp2 = 0.21 and F4.4,162.4 = 17.89, p < 0.001, ηp2 = 0.33, respectively]. Polydipsic water intake began differing from baseline prandial water drinking from FT-60 s session 4 and 3 onwards, respectively (Fig. 2C).
As previously described [31, 52, 72, 75], marked individual differences in the propensity to lose control over adjunctive water drinking behavior emerged over 20 SIPw sessions [main effect of group: F1,19 = 52.73, p < 0.001, ηp2 = 0.73; time: F20,380 = 16.39, p < 0.001, ηp2 = 0.46 and group × time interaction: F20,380 = 11.74, p < 0.001, ηp2 = 0.38] (Fig. 2C). Thus, HD rats (upper quartile of the population, n = 10) were prone readily to develop hyperdipsia, eventually drinking more than 15 mL/h (15.30 ± 1.21 mL over the last three sessions). This represents more than twice the intake of the population as a whole, and almost ten times more than that of LD rats (lower quartile of the population, n = 11) (1.70 ± 0.32 mL over the last three sessions) [main effect of group: F1,19 = 113.19, p < 0.001, ηp2 = 0.86] (Fig. 2D, left panel). This differential level of polydipsic water intake observed between HD and LD rats after 20 daily SIP sessions remained stable over long periods of time, as previously shown [31].
However, the introduction of the opportunity to drink alcohol instead of water as a means to cope with the stress induced by the SIP procedure resulted in an asymmetrical change in the level of adjunctive drinking displayed by LD and HD rats [main effect of time: F2,38 = 9.86, p < 0.001, ηp2 = 0.34; group: F1,19 = 39.81, p < 0.001, ηp2 = 0.68 and time × group interaction: F2,38 = 22.95, p < 0.001, ηp2 = 0.55] (Fig. 2D). Thus, despite an overall decrease in total fluid intake upon the introduction of alcohol, reflective of its anxiolytic properties [76], HD and LD rats initially maintained their then long established difference in the level of adjunctive drinking [HD (5.14 ± 0.84 mL/h) vs LD (2.54 ± 0.69 mL/h, p = 0.006] (Fig. 2D, middle panel). However, over the 20 sessions of SIPa some LD rats (63.6%) more than doubled their intake of alcohol, eventually reaching 5.14 ± 1.12 mL per hour [LD, Early SIPa vs Late SIPa, p < 0.001] (Fig. 2D). These alcohol coper LD rats that relied on alcohol to develop a coping response contributed greatly to the overall increase in alcohol intake shown at the group level [main effect of time F9.2,340 = 12.49, p < 0.001, η2 = 0.25] (Fig. 2C). Alcohol coper LD rats became as compulsive as HD rats [Late SIPa, HD vs LD, p = 0.09], which only increased their alcohol intake by ~50% over the same period [HD, Early SIPa vs Late SIPa, p < 0.02] (Fig. 2D, right panel).
The sudden increase in fluid consumption shown by alcohol coper LD rats upon the introduction of alcohol cannot be accounted for by a simple increase in fluid intake over time since once established, polydipsic water intake remained stable over protracted periods of time (e.g., at least 40 days) [main effect of session: F40,360 = 9.49, p < 0.001, ηp2 = 0.51; from session 15, between sessions all p values > 0.05] (Fig. 2E). Thus, rats exposed to the SIP procedure displayed similar level of polydipsic water intake over sessions 38–40 as over sessions 18–20 [F1,9 = 2.80, p = 0.13, η2 = 0.24] (Fig. 2E, insert). Instead, as shown in an independent replication on a cohort of 47 individuals [main time × group × liquid interaction: F19,304 = 10.33, p < 0.001, ηp2 = 0.39] (Fig. 3A), the emergence of polydipsic alcohol drinking in the LD rats that relied on alcohol to engage in a coping strategy (alcohol copers) resulted in a decrease in their anxiety level, as assessed on an EPM [One tailed t-test: t = −2.25, p < 0.05] (Fig. 3B). The anxiolytic properties of the development of an adjunctive response using alcohol drinking were further supported at the population level by the emergence of a negative correlation between the level of polydipsic drinking and anxiety measured immediately after a SIP session following the introduction of alcohol (Fig. 3C, D) [R = 0.31, p < 0.05].

A At the population level, individuals exposed to a SIP procedure developed a polydipsic drinking coping response with water that they maintained over time. However, some individuals (High drinker rats or Water coper rats, WC, upper quartile of the population) developed an excessive compulsive polydipsic drinking behavior while others (Low drinker rats, LD) did not acquire this adjunctive behavior. Similarly to experiment 2, while WC rats maintained overall a steady level of polydipsic drinking over time when alcohol was introduced in place of water, some LD rats acquired a coping response with alcohol (alcohol coper rats, AC) and eventually developed compulsive polydipsic alcohol drinking so that they eventually no longer differed from WC rats after 20 SIPa sessions. B Trajectories in anxiety levels (assessed as the percentage of time in the open arms of an elevated plus maze in a naïve state or immediately following a SIP session with water (SIPw) or alcohol (SIPa) after at least 20 daily sessions each) revealed that the polydipsic alcohol drinking displayed by the LD rats that relied on alcohol to engage in a coping strategy (AC) resulted in a decrease in their anxiety levels, as compared to those measured after a SIPw session. C, D Furthermore, at the population level, a negative correlation emerged between the level of polydipsic drinking and anxiety following the introduction of alcohol, which revealed the anxiolytic nature of polydipsic alcohol drinking. *p < 0.05, AC, SIPw/N vs SIPa/w; #p <0.001, time x group x liquid interaction; ns: not significant. B: baseline session; EPM: elevated plus maze; N: naïve state; SIPw/N: change in anxiety levels after being trained in the SIPw vs the naïve state; SIPa/w: change in anxiety levels after being trained in SIPa vs SIPw.
These results together replicate both previous evidence of individual differences in the tendency to lose control over stress reducing polydipsic water drinking [52, 77, 78] and our original demonstration of individual differences in the reliance on alcohol to cope with stress, the latter being a gateway for the development of excessive and compulsive alcohol drinking [52].
We then sought to identify the neural locus of control of these coping behaviors that appear initially to be goal-directed, the goal of the response (drinking) being the alleviation of stress [14, 21, 79], but that eventually become excessive and compulsive [75, 80, 81]. This transition is hypothesized here to reflect the development of maladaptive negative-reinforcement driven habits. Thus, we assessed the reliance of adjunctive fluid drinking on aDLS DA, a signature of habitual control over behavior, including alcohol-related responding [64], and the compulsion to seek and drink alcohol [55, 56].
Hyperdipsia is associated with the functional engagement of the aDLS DA-dependent habit system
As predicted, after a short-term exposure to SIPw, prior to the development of excessive drinking behavior in vulnerable individuals, aDLS DA receptor blockade with α-flupentixol (0, 6, 12 μg/0.5 μl/side; between-subject design) had no effect on water intake [main effect of treatment: F2,19 = 2.11, p = 0.13, ηp2 = 0.10] (Test 1, Early SIPw, Fig. 4A). However, after extended exposure to SIPw, at a time when vulnerable individuals had developed compulsive adjunctive drinking, the same aDLS DA receptor blockade (0, 6, 12 μg/0.5 μl/side α-flupentixol; within-subject counter-balanced design) now resulted in a marked decrease in adjunctive water intake in these HD rats [HD vs LD: main effect of treatment: F2,38 = 4.16, p = 0.023, ηp2 = 0.18; group: F1,19 = 44.85, p < 0.001, ηp2 = 0.70 and treatment × group interaction: F2,38 = 2.96, p = 0.06, ηp2 = 0.13].

Intra-anterior dorsolateral striatum (aDLS) infusion of the dopamine receptor antagonist α-flupentixol (0, 6, 12 μg/side) had no effect on recently acquired polydipsic water drinking (Test 1, A) or on drinking shown by low drinker (LD) rats even after 20 sessions (Test 2, B). In contrast, similar intra aDLS infusions of α-flupentixol (0, 6, 12 μg/side) decreased excessive water intake in high drinkers (HD) rats (Test 2, B). When water was replaced by alcohol, intra aDLS dopamine receptor blockage no longer influenced polydipsic drinking in HD rats while it remained ineffective in LD rats (Test 3, C). In marked contrast, the sensitivity shown by HD rats to intra aDLS dopamine receptor blockade when engaged in excessive polydipsic water intake re-emerged when their polydipsic alcohol intake became well established (Test 4, D). E–H The functional engagement of the aDLS DA-dependent habit system when polydipsic water or alcohol drinking becomes compulsive, namely at tests 2 and 4, is not due to a floor effect inherent to relatively lower levels of non-prandial drinking at tests 1 and 2 since no correlations were observed between the magnitude of the decrease in fluid intake following intra aDLS infusion of either dose of α-flupentixol and the baseline level of water (E, F) or alcohol (G, H) adjunctive drinking. ** p < 0.01, * p < 0.05 different from vehicle (0 μg/side). SIPw: SIP with water; SIPa: SIP with alcohol; α-flu: α-flupentixol. ∆water intake (%bsl/veh): difference in water intake following aDLS DA receptor blockage and baseline (bsl, for test 1) or intra aDLS infusion of vehicle (for test 2). ∆alcohol intake (%bsl): difference in alcohol intake following aDLS DA receptor blockage vs intra aDLS infusion of vehicle (tests 3 and 4). In E, F pink arrows indicate the spread of the reliance on the aDLS DA-dependent system.
Follow-up analyses confirmed that aDLS DA receptor blockade did not influence drinking in LD rats [main effect of treatment: F2,20 = 0.31, p = 0.74, ηp2 = 0.03], while it greatly reduced excessive adjunctive drinking in HD rats [main effect of treatment: F2,18 = 3.55, p = 0.049, ηp2 = 0.28; 0 vs 6 and 12 µg/side: p = 0.007] (Test 2, Late SIPw, Fig. 4B).
In marked contrast, in the same rats, the introduction of alcohol as a means to cope with stress resulted in a disengagement, albeit transient (see below), of aDLS DA control over adjunctive drinking. Thus, intra aDLS infusions of the DA receptor antagonist α-flupentixol no longer decreased fluid intake in HD rats after 3 days of SIPa [HD vs LD: main effect of group: F1,19 = 4.95, p = 0.038, ηp2 = 0.21, treatment: F2,38 = 1.44, p = 0.25, ηp2 = 0.07 and treatment × group interaction: F2,38 = 0.90, p = 0.42, ηp2 = 0.04] (Test 3, Early SIPa, Fig. 4C). These results thereby demonstrate that learning to drink alcohol to cope with stress, even in individuals that had established a compulsive adjunctive drinking behavior with water, is associated with a disengagement of habitual control over behavior.
However, following 20 daily sessions of SIPa when adjunctive alcohol drinking had escalated and become excessive, it became, once again, reliant on aDLS DA, as shown by the emergence of a sensitivity of alcohol drinking to aDLS DA receptor blockade in HD rats [HD vs LD: main effect of group: F1,19 = 1.17, p = 0.29, ηp2 = 0.06 and treatment x group interaction: F2,38 = 4.76, p = 0.014, ηp2 = 0.20; HD rats, 0 vs 12 μg/side: p = 0.044] (Test 4, Late SIPa, Fig. 4D). The functional engagement of the aDLS DA-dependent habit system selectively when polydipsic water or alcohol drinking becomes compulsive cannot be explained by a floor effect in early water or alcohol drinking stages since the magnitude of the decrease in fluid intake following intra aDLS infusion of either dose of α-flupentixol was not related to the baseline level of drinking (Fig. 4E–H).
Differential trajectories of the functional engagement of the habit system predict individual differences in the reliance on drinking alcohol to engage a coping strategy
Building on previous evidence in a positive reinforcement setting that the reliance of habitual alcohol seeking on aDLS DA predicts the severity of the ensuing compulsive behavior [56], here we further investigated whether a similar relationship was observed between the reliance of adjunctive responses, maintained through negative reinforcement, on aDLS DA and the vulnerability to develop excessive, compulsive adjunctive drinking.
We capitalized on the within-subject design of experiment 2 to determine, for each individual, the differential sensitivity of excessive adjunctive drinking of water (Test 2) vs alcohol (Test 4) to intra aDLS infusions of α-flupentixol (mean % decrease of intake following DA receptor blockade relative to baseline, i.e., following vehicle infusions).
A K-mean cluster analysis [55, 71] revealed three sub-populations that differed in the reliance of their excessive water or alcohol adjunctive drinking on aDLS DA (Fig. 5A). A first cluster (Cluster 1, 28.9% of the total population) comprised individuals that had all developed an aDLS DA-dependent adjunctive water drinking behavior before they switched to alcohol drinking, which also became reliant on aDLS DA.

A, B Marked individual differences in the reliance of compulsive drinking on anterior dorsolateral striatum (aDLS) dopamine (DA) were revealed when normalizing the influence of aDLS DA receptor blockade on polydipsic water (Late SIP water) or alcohol drinking (Late SIP alcohol) to the baseline levels of drinking following vehicle infusions (α-flupentixol 0 μg/side). A cluster analysis identified three subpopulations of rats, one (Cluster 1) comprised individuals whose polydipsic behavior, irrespective of the fluid drank, was heavily reliant on aDLS DA. These highly aDLS reliant water copers (WC) represented 28.9% of the overall population and consisted predominantly (82%) of high drinkers (HD) and intermediate individuals. A second cluster (Cluster 2) comprised individuals whose polydipsic water or alcohol drinking behavior was overall marginally sensitive to aDLS DA receptor blockade. These marginally aDLS reliant WC represented 50% of the population and consisted predominantly (84%) of low drinkers (LD) and intermediate rats. The third cluster (Cluster 3) comprised individuals whose polydipsic alcohol drinking was much more reliant on aDLS DA than their polydipsic water drinking behavior. These highly aDLS reliant alcohol coper (AC) rats represented 21% of the population and consisted predominantly (87.5%) of LD and intermediate rats. A systematic analysis of the respective reliance of polydipsic water vs alcohol drinking behavior on aDLS DA confirmed that only highly aDLS reliant AC rats displayed a selective increased in the engagement of aDLS DA-dependent control over behavior when they used alcohol as a mean to cope with stress. C Retrospective analysis of the acquisition and expression of polydipsic water and/or alcohol drinking of these three clusters confirmed that Cluster 1 WC rats developed hyperdipsia with water while Cluster 3 AC rats did so with alcohol. D–F Marked differences were observed in the time course of the engagement of the aDLS DA-dependent habit system in mediating compulsive coping behavior in the individuals of these three groups that map perfectly those observed on HD and LD rats (Fig. 3), in that rats from Cluster 1 showed sensitivity to aDLS DA receptor blockade when SIPw (Test 2 Late SIPw) and SIPa (Test 4 Late SIPa) were well-established whereas rats from Cluster 3 showed such sensitivity only when SIPa was well-established, neither showing reliance on aDLS DA during the acquisition of SIPa. Rats from the heterogenous Cluster 2 never showed a decrease in adjunctive drinking following aDLS that reached statistical significance throughout the training history. *** p ≤ 0.001, ** p ≤ 0.01, compared with the vehicle treatment (0 μg/side); # p < 0.001 compared with cluster 1 and 2; @: main cluster × liquid × period interaction, p < 0.001.
These aDLS reliant water coper rats, which comprised 82% of HD and intermediate individuals, differed from rats of Cluster 2 (50% of the population; which instead comprised 84% of intermediate and LD rats) whose water drinking was marginally reliant on aDLS DA and who did not engage their habit system when subsequently exposed to alcohol.
In contrast, individuals in Cluster 3 (21.1% of the population, which comprised 87.5% of LD and intermediate rats) were the only ones to develop an adjunctive response reliant on aDLS DA when they used alcohol, as confirmed by an analysis of the ratio of sensitivity of adjunctive fluid intake to aDLS DA receptor blockade between SIPa and SIPw [main effect of cluster: F2,35 = 13.33, p < 0.001, ηp2 = 0.43; Cluster 3 vs 1 and 2, p < 0.001, respectively) (Fig. 5B, right panel).
Retrospective investigation of the individuals comprising each cluster revealed that most of those belonging to Cluster 1 were water coper HD rats whereas the majority of individuals in Cluster 3 were alcohol coper LD rats that had instead developed hyperdipsia after the introduction of alcohol. This was further supported by the analysis of the acquisition and expression of polydipsic water and/or alcohol drinking of these three clusters (Fig. 5C): aDLS DA reliant water copers and aDLS DA reliant alcohol copers (cluster 1 and 3, respectively) had different coping styles accompanied by different trajectories in the development of their hyperdipsia [main cluster × fluid (water vs alcohol) × period (acquisition vs expression) interaction: F18,315 = 2.64, p < 0.001, ηp2 = 0.13]. Cluster 1 water coper rats developed hyperdipsia with water while cluster 3 alcohol coper rats did so with alcohol, increasing their polydipsic alcohol intake by 60% over the 3 weeks of SIPa, eventually to reach the level of intake shown by water coper rats.
The time course of the engagement of aDLS DA in the control over adjunctive behavior of Cluster 3 alcohol coper rats was therefore very different to that shown by Cluster 1 water coper rats (Fig. 5D–F) [treatment x time x cluster interaction: F5,87.3 = 4.48, p = 0.001, ηp2 = 0.20; treatment × time interaction: F2.5,87.3 = 4.48, p = 0.005, ηp2 = 0.12], with profound differences in their sensitivity to aDLS DA receptor antagonism at Late SIPw (Test 2, Fig. 5D) [main effect of treatment: F1.5,52.2 = 5.01, p = 0.017, ηp2 = 0.12; treatment × cluster interaction: F3,52.2 = 6.16, p = 0.001, ηp2 = 0.26] and at Late SIPa (Test 4, Fig. 5F) [main effect of treatment: F1.7,60.1 = 6.49, p = 0.004, ηp2 = 0.16; treatment × cluster interaction: F3.4,60.1 = 7.55, p < 0.001, ηp2 = 0.30], but not Early SIPa (Test 3, Fig. 5E) [main effect of treatment: F2,70 = 1.22, p = 0.30, ηp2 = 0.03; treatment × cluster interaction: F4,70 = 0.21, p = 0.93, ηp2 = 0.01].
The differences at Late SIPw were driven exclusively by Cluster 1 water coper individuals [0 vs 6 μg/side: p < 0.001, 0 vs 12 μg/side: p < 0.001] (Fig. 5D), whereas the effects at Late SIPa were driven by Cluster 1 water coper and Cluster 3 alcohol coper individuals [Cluster 1, 0 vs 12 μg/side: p = 0.001; Cluster 3, 0 vs 12 μg/side: p = 0.010] (Fig. 5F).
Together these data demonstrate that the development of compulsive adjunctive drinking depends on the engagement of aDLS DA-dependent control over behavior that eventually occurred in 81.6% of the individuals in this study.
Discussion
At the population level, individuals in the present study developed an adaptive coping response under a SIP procedure [75], expressed as non-regulatory adjunctive polydipsic drinking behavior. While exposure to intermittent food delivery in food restricted individuals activates the hypothalamic-pituitary-adrenal (HPA) axis [13] and increases anxiety [31], the development of polydipsic water drinking, which was shown here to remain stable over up to 42 days, has long been shown to reduce the activity of the HPA axis [52, 77, 78].
However, within 2 weeks some individuals progressively lost control over the polydipsic drinking response and developed hyperdipsia, a compulsive coping strategy [74, 75, 80,81,82]. In these HD rats, no changes in anxiety levels were seen following SIPw, revealing that in these individuals, even though it has become compulsive, coping behavior remains acutely anxiolytic [77] and mitigates the otherwise anxiety-inducing effects of the procedure [31]. This observation lends additional support to the construct validity of hyperdipsia as a model of compulsive behavior, since in compulsive disorders compulsions remain acutely anxiolytic in the context of an overall stress surfeit [83].
In contrast, some individuals did not acquire a coping response with water, but only did so when they had access to alcohol. These alcohol coper LD rats learnt to drink alcohol under the anxiogenic effects of intermittent food delivery, but had not done so by engaging in polydipsic water drinking. Moreover, they increased their daily intake of alcohol more rapidly than any other group, and eventually developed compulsive coping behavior. Engaging in polydipsic alcohol drinking, which was shown here to decrease anxiety in alcohol copers, has previously been shown to decrease plasma corticosterone concentration [84] at the population level. It has also been shown to be unrelated to ethanol preference or a tendency to drink more alcohol outside the SIP context [52, 84] and also cannot be solely accounted for by any direct effect of alcohol on the HPA axis [84]. Since it is the stress-reducing properties of the adjunctive response and not the fluid ingested that maintains polydipsic drinking under negative reinforcement, exposure to SIPa does not influence the reinforcing properties of alcohol in a different context [85].
The results of the present study replicate previously reported individual variability in the tendency both to engage in polydipsic drinking [52] and to develop compulsive adjunctive behaviors [31, 52, 72,73,74,75, 80, 81, 86], and more recent evidence that some individual rats that do not develop a coping response with water, do so readily when drinking alcohol [52].
In addition, the results of the present study provide causal evidence of a progressive engagement of aDLS DA in the control over adjunctive behavior, especially when it becomes compulsive in vulnerable individuals. While the acquisition of polydipsic drinking of water or alcohol was not dependent on aDLS DA, its excessive and compulsive manifestation in vulnerable individuals was selectively decreased by bilateral aDLS DA receptor blockade at a time it has previously been shown no longer to be sensitive to dopaminergic manipulations of the mesolimbic system [61].
In alcohol copers, this engagement of aDLS DA habit system [87, 88] in the control over polydipsic alcohol drinking was specific to a facilitated transition to compulsion by acquiring alcohol use as a self-medication strategy. It could not be attributed to the emergence of habitual control over behavior by protracted training [89] because:
-
(i)
the individual differences observed in the tendency to develop excessive polydipsic drinking after 20 sessions of SIPw did not evolve further across an additional period of 20 sessions when animals continued to be given access to water instead of alcohol, see [31],
-
(ii)
and some individuals that had not engaged aDLS DA when drinking under SIPw also did not do so when subsequently given access to alcohol (Fig. 5).
Finally, the emerging reliance on aDLS DA of alcohol drinking as a coping behavior cannot be explained by a difference in the level of fluid intake: while the level of compulsive fluid intake shown by HD rats at the end of the 20 sessions of SIPa was much lower than that shown at the end of the 20 sessions of SIPw, in both cases their excessive adjunctive behavior was equally sensitive to aDLS DA receptor blockade. In addition, as shown in Fig. 4, the reliance on aDLS control is not predicated on the level of fluid intake.
Importantly, although they decreased the volume of fluid they drank when alcohol was introduced, HD rats still ingested 5–8 ml/h of alcohol, which is more than double the volume drunk by the same strain of rats (1–3 ml/h) following at least 12 sessions of intermittent access to 10% alcohol in a two-bottle choice procedure, data from [52], a time when rats had already escalated their alcohol intake. These observations reveal the excessive nature of their alcohol intake under SIP [90], which has previously been shown to result in blood alcohol levels higher than 0.6 g.l−1 [52].
These results are consistent with a body of evidence suggesting marked differences in the psychological and neural mechanisms underlying the acquisition of adjunctive coping responses in males and its subsequent compulsive manifestation in vulnerable individuals. However, women differ from men in coping strategies and in the impact of stress on their alcohol use or misuse [91, 92], and female rodents develop higher levels of SIP for water [93, 94] or alcohol [95] than males. Thus, further research is warranted to investigate whether the psychological and neural mechanisms identified here that result in the development of compulsive adjunctive behaviors in males may be even more readily engaged in females.
Because adjunctive behaviors do not lead to the delivery of food or help to meet homeostatic need, for review see [79], they have long been considered not to be instrumental. However, since the outcome of coping strategies is not the procurement of an external outcome, but a change in an interoceptive state, namely anxiolysis [96], adjunctive behavior, manifested here as polydipsic drinking, which superficially may appear to be consummatory, is in fact an instrumental response [79, 97, 98]. However, this instrumental response relies, like comfort eating, on the association between the ingestive behavior and stress reduction. Hence, the polydipsic drinking is elicited and maintained by the negative reinforcing properties of stress/anxiety reduction (Fig. 3 and [77]) and its associated interoceptive mechanisms [99].
This view is supported by evidence that the engagement in polydipsic drinking during the first week of exposure to SIP depends on the interoceptive cortex, namely the anterior insula [31], and results in a decrease in the activation of the HPA axis provoked by intermittent food delivery [14, 15, 19,20,21, 77]. Furthermore, at the same early stage of training, prevention of drinking in the SIP context results in an increase in corticosterone levels [13].
Early polydipsic water intake has features of goal-directedness in that it is sensitive to outcome devaluation (anti-anxiolysis), since there is a decrease in responding following administration of anxiogenic drugs such as CRF [100] or amphetamine [61, 101]. In contrast, following prolonged exposure to SIP, the same acute amphetamine challenge no longer results in a decrease in drinking [101]. These observations suggest that polydipsic drinking is initially a response tied to the motivational value of its outcome, namely anxiolysis, but that it eventually becomes habitual [102].
Development of habitual control over polydipsic drinking is consistent with a progressive shift in the striatal locus of control over behavior under SIP from a network involving the ventral and dorsomedial striatum to one involving the DLS [55, 56, 63, 70, 103]. Selective 6-hydroxydopamine lesions of the mesolimbic system prevent the acquisition of SIP [104,105,106], which is otherwise associated with a ramping of extracellular levels of DA in the nucleus accumbens core across sessions [107]. In contrast, DA transporter deficient mice and rats, which have elevated levels of DA, display an impaired acquisition of polydipsic drinking [108, 109], which is also decreased in wild type animals by the acute administration of psychostimulants such as cocaine and D-amphetamine directly into the nucleus accumbens [30, 110]. Together these results show that the acquisition of adjunctive drinking behavior depends on DA signalling in the mesolimbic system, but that it is impaired by an increase in tonic extracellular levels of DA.
In marked contrast, when this coping behavior is well established and becomes excessive in vulnerable individuals, it is no longer impaired by similar causal manipulations of the mesolimbic DA system [61]. Furthermore, repeated amphetamine exposure, which facilitates habitual responding under positive reinforcement [111], exacerbates polydipsic drinking [112] and renders it independent of food delivery [112, 113], suggesting the development of stimulus-response control over behavior [114] when it has become compulsive in vulnerable individuals. These observations are in line with the evidence that polydipsic drinking during early SIPw training is not accompanied by neuronal activation in the DLS [115], whereas its compulsive manifestation after extended exposure is associated with increased spine density in this region [62].
The apparent shift from a network involving the ventral striatum to one involving the DLS in the locus of control over adjunctive behaviors when they become compulsive [116] agrees with previous evidence that the development of compulsive alcohol seeking in vulnerable individuals depends on the prior functional engagement of aDLS DA [56]. This lends further support to the hypothesis that the development of compulsion stems from a loss of control over aDLS-dependent habits [117,118,119,120].
While stress shifts the balance toward aDLS-dependent habits from ventral striatal-dependent, goal-directed, behavior [121,122,123], it is unlikely that individual differences in the tendency to develop compulsive polydipsic water drinking observed here and elsewhere [31, 72,73,74] are due to a differential sensitivity to stress. Individuals that did not acquire a coping response with water did so readily when they had access to alcohol instead. This suggests that the lack of development of an adjunctive behavior under SIPw in these individuals was not due to their lack of sensitivity to the stress engendered by the SIP procedure. Instead, these individuals needed alcohol in order to develop this coping response.
In contrast, the tendency of water copers to develop a coping response with water and the subsequent loss of control over drinking in vulnerable individuals may reflect an interplay between a heightened sensitivity to negative urgency [124] and an inherent propensity generally to rely on the aDLS habit system. Hence, on the one hand, the development of compulsive SIP depends on both insula-dependent interoceptive mechanisms, the noradrenaline stress system and their interaction with pre-existing impulsivity [31, 72]. Whereas on the other hand, it is also predicted by a pre-existing tendency to use the habit system across a wide array of tasks, from response learning in spatial navigation [125] to resistance to devaluation in instrumental reinforcement and perseverative responses in reversal learning task [115, 126]. Overall, these results support the view that the inflexibility and increased responding under operant schedules that promote habitual behavior shown by high water drinker (HD) rats [74, 127] arises from an interaction between an inherent tendency to rely on the habit system and its recruitment by negative reinforcement during the development of a coping response.
Using the same SIP procedure, we have previously shown that alcohol enables the development of adjunctive responses in a specific subpopulation of rats otherwise unable to cope with stress by drinking water. In this subpopulation of LD rats, the phenotype of which was also identified independently in experiment 4 of the present study, it is the acquisition of alcohol use as a self-medication strategy, and not the overall level of intoxication (alcohol coper rats did not differ from water copers in their overall level of alcohol intake or the blood alcohol levels they reached) that determined their greater vulnerability to develop compulsive, quinine resistant, alcohol drinking [52]. This subpopulation of alcohol coper rats is very similar to that identified as Cluster 3 in the present study which only engaged aDLS DA when alcohol was introduced in the SIP procedure.
The engagement of an aDLS DA-dependent circuit in these compulsive alcohol drinkers is consistent with previous evidence of the recruitment of this circuitry following chronic exposure to alcohol and other addictive drugs in humans [59, 128], non-human primates [129, 130] and rodents [56, 63,64,65,66, 103, 131,132,133]. This may in turn be attributable to the adaptations that alcohol exposure causes in the aDLS, including its disinhibition by the dampening of GABAergic transmission onto its principal medium spiny projection neurons [134, 135].
Together, these observations support the view that the engagement of habitual control over negative-reinforcement driven behavior, such as the acquisition of alcohol drinking as a self-medication strategy, is an important determinant of the vulnerability to develop compulsive drinking [50, 54, 70, 118, 136].
It will be important to determine whether such negative reinforcement-driven, stress-bound maladaptive coping habits develop similarly when individuals learn to take drugs to alleviate negative states of withdrawal [137], and the extent to which they facilitate relapse under states of heightened stress [138, 139], as suggested by Marlatt’s taxonomy [140,141,142].
It remains also to be established why some individuals can only develop a coping response with alcohol but not water. One hypothesis is that the anxiolytic properties of alcohol (Fig. 3B–D) [143, 144] facilitate the acquisition of the adjunctive response in these individuals [145, 146]. This is also supported by the marked decrease in total fluid intake shown by water copers after the introduction of alcohol, which is quantitatively similar to that seen following administration of benzodiazepines [147].
The introduction of alcohol in these water coper rats resulted in a transient disengagement of the aDLS DA-dependent habit system. This reveals that the same behavioral expression of a coping response can be mediated by either the goal-directed or the habit system, as is the case for instrumental responses for positive reinforcers [65]. But perhaps more importantly it suggests that incentive learning can influence even a well-established, negative reinforcement-based habitual coping response [102, 148] when alcohol is adjunctively drunk, as has also been shown in rats self-administering heroin under a state of withdrawal [137].
Together these observations provide evidence for a role of negative reinforcement-based habits in the development of compulsive coping behaviors.
References
John OP, Gross JJ (eds). Handbook of Emotion Regulation. New York:The Guilford Press; 2007.
Kobylinska D, Kusev P. Flexible Emotion Regulation: How Situational Demands and Individual Differences Influence the Effectiveness of Regulatory Strategies. Front Psychol. 2019;10:72.
Lazarus RS, Folkman S. Stress, appraisal, and coping. New York: Springer; 1984.
Gross JJ. Emotion regulation: Past, present, future. Cogn Emot. 1999;13:551–73.
Gross JJ, John OP. Individual differences in two emotion regulation processes: implications for affect, relationships, and well-being. J Pers Soc Psychol. 2003;85:348–62.
Bonanno GA, Burton CL. Regulatory Flexibility: An Individual Differences Perspective on Coping and Emotion Regulation. Perspect Psychol Sci. 2013;8:591–612.
Ray C, Lindop J, Gibson S. The concept of coping. Psychol Med. 1982;12:385–95.
Mohiyeddini C, Semple S. Displacement behaviour regulates the experience of stress in men. Stress. 2013;16:163–71.
Mohiyeddini C, Bauer S, Semple S. Public self-consciousness moderates the link between displacement behaviour and experience of stress in women. Stress. 2013;16:384–92.
Tugade MM, Fredrickson BL. Regulation of Positive Emotions: Emotion Regulation Strategies that Promote Resilience. J Happiness Stud. 2007;8:311–33.
Mayordomo T, Viguer P, Sales A, Satorres E, Meléndez JC. Resilience and Coping as Predictors of Well-Being in Adults. J Psychol. 2016;150:809–21.
Dantzer R, Mormede P. Pituitary-adrenal consequences of adjunctive activities in pigs. Horm Behav. 1981;15:386–95.
Tazi A, Dantzer R, Mormede P, Le Moal M. Pituitary-adrenal correlates of schedule-induced polydipsia and wheel running in rats. Behav Brain Res. 1986;19:249–56.
Brett LP, Levine S. Schedule-induced polydipsia suppresses pituitary-adrenal activity in rats. J Comp Physiol Psychol. 1979;93:946–56.
Mittleman G, Jones GH, Robbins TW. The relationship between schedule-induced polydipsia and pituitary-adrenal activity: pharmacological and behavioral manipulations. Behav Brain Res. 1988;28:315–24.
Falk J. Production of polydipsia in normal rats by an intermittent food schedule. Science. 1961;133:195–6.
Killeen PR, Hanson SJ, Osborne SR. Arousal: its genesis and manifestation as response rate. Psychol Rev. 1978;85:571–81.
Levine S, Weinberg J, Brett LP. Inhibition of pituitary-adrenal activity as a consequence of consummatory behavior. Psychoneuroendocrinology. 1979;4:275–86.
Lopez-Grancha M, Lopez-Crespo G, Venero C, Canadas F, Sanchez-Santed F, Sandi C, et al. Differences in corticosterone level due to inter-food interval length: implications for schedule-induced polydipsia. Horm Behav. 2006;49:166–72.
Levine R, Levine S. Role of the pituitary-adrenal hormones in the acquisition of schedule-induced polydipsia. Behav Neurosci. 1989;103:621–37.
Dantzer R, Terlouw C, Mormede P, Le Moal M. Schedule-induced polydipsia experience decreases plasma corticosterone levels but increases plasma prolactin levels. Physiol Behav. 1988;43:275–9.
Horwood S, Anglim J. Problematic smartphone usage and subjective and psychological well-being. Comput Hum Behav. 2019;97:44–50.
Goodwin H, Haycraft E, Meyer C. The relationship between compulsive exercise and emotion regulation in adolescents. Br J Health Psychol. 2012;17:699–710.
Moritz S, Jahns AK, Schroder J, Berger T, Lincoln TM, Klein JP, et al. More adaptive versus less maladaptive coping: What is more predictive of symptom severity? Development of a new scale to investigate coping profiles across different psychopathological syndromes. J Affect Disord. 2016;191:300–7.
Lawrence LM, Elphinstone B. Coping associated with compulsive buying tendency. Stress Health. 2021;37:263–71.
Belin D, Daniel M, Lacoste J, Belin-Rauscent A, Bacconier M, Jaafari N. Insight: New vistas into an aetiological and phenomenological role in the psychopathology of obsessive compulsive disorders. Ann médico-psychologiques. 2011;169:420–5.
Whitehead MR, Suveg C. Difficulties in emotion regulation differentiate depressive and obsessive–compulsive symptoms and their co-occurrence. Anxiety, Stress, Coping. 2016;29:507–18.
Crisson JE, Keefe FJ. The relationship of locus of control to pain coping strategies and psychological distress in chronic pain patients. Pain. 1988;35:147–54.
Moore CF, Sabino V, Koob GF, Cottone P. Neuroscience of Compulsive Eating Behavior. Front Neurosci. 2017;11:469.
Lopez-Grancha M, Lopez-Crespo G, Sanchez-Amate MC, Flores P. Individual differences in schedule-induced polydipsia and the role of gabaergic and dopaminergic systems. Psychopharmacol (Berl). 2008;197:487–98.
Belin-Rauscent A, Daniel ML, Puaud M, Jupp B, Sawiak S, Howett D, et al. From impulses to maladaptive actions: the insula is a neurobiological gate for the development of compulsive behavior. Mol Psychiatry. 2016;21:491–9.
Yao N, Nazari N, Veiskarami HA, Griffiths MD. The role of healthy emotionality in the relationship between fear of COVID-19 and mental health problems: a cross-sectional study. Cogn Process. 2022;23:569–81.
Waugh CE, Leslie-Miller CJ, Cole VT. Coping with COVID-19: the efficacy of disengagement for coping with the chronic stress of a pandemic. Anxiety Stress Coping. 2023;36:52–66.
Koole SL, Rothermund K. Coping with COVID-19: Insights from cognition and emotion research. Cogn Emot. 2022;36:1–8.
Dimanova P, Borbas R, Schnider CB, Fehlbaum LV, Raschle NM. Prefrontal Cortical Thickness, Emotion Regulation Strategy use and Covid-19 Mental Health. Soc Cogn Affect Neurosci. 2022;17:877–89.
Stanton R, To QG, Khalesi S, Williams SL, Alley SJ, Thwaite TL, et al. Depression, Anxiety and Stress during COVID-19: Associations with Changes in Physical Activity, Sleep, Tobacco and Alcohol Use in Australian Adults. Int J Environ Res Public Health. 2020;17:4065.
Ramalho R. Alcohol consumption and alcohol-related problems during the COVID-19 pandemic: a narrative review. Australas Psychiatry. 2020;28:524–6. 1039856220943024
Neill E, Meyer D, Toh WL, van Rheenen TE, Phillipou A, Tan EJ, et al. Alcohol use in Australia during the early days of the COVID-19 pandemic: Initial results from the COLLATE project. Psychiatry Clin Neurosci. 2020;74:542–9.
Dubey MJ, Ghosh R, Chatterjee S, Biswas P, Chatterjee S, Dubey S. COVID-19 and addiction. Diabetes Metab Syndr. 2020;14:817–23.
Tartaglia S, Bergagna E. Alcohol consumption as a maladaptive coping strategy to face low life satisfaction. Drugs: Educ Prev Policy. 2020;27:306–11.
Khantzian EJ. The self-medication hypothesis of addictive disorders: focus on heroin and cocaine dependence. Am J Psychiatry. 1985;142:1259–64.
Cooper ML, Frone MR, Russell M, Mudar P. Drinking to regulate positive and negative emotions: a motivational model of alcohol use. J Pers Soc Psychol. 1995;69:990–1005.
Tice DM, Bratslavsky E, Baumeister RF. Emotional distress regulation takes precedence over impulse control: If you feel bad, do it! J Personal Soc Psychol. 2001;80:53–67.
Weiss NH, Bold KW, Sullivan TP, Armeli S, Tennen H. Testing bidirectional associations among emotion regulation strategies and substance use: a daily diary study. Addiction. 2017;112:695–704.
Cavicchioli M, Kraslavski A, Movalli M, Maffei C, Ogliari A. The investigation of mechanisms underlying addictive behaviors: a case-control study. J Addict Dis. 2022;40:306–25.
Blevins CE, Abrantes AM, Stephens RS. Motivational pathways from antecedents of alcohol use to consequences: a structural model of using alcohol to cope with negative affect. Am J Drug Alcohol Abus. 2016;42:395–403.
Zeidner M, Saklofske D. Adaptive and maladaptive coping. Handbook of coping: Theory, research, applications. Oxford, England: John Wiley & Sons; 1996. p. 505–31.
Ramachandran A, Makhashvili N, Javakhishvili J, Karachevskyy A, Kharchenko N, Shpiker M, et al. Alcohol use among conflict-affected persons in Ukraine: risk factors, coping and access to mental health services. Eur J Public Health. 2019;29:1141–6.
Clay JM, Parker MO. Alcohol use and misuse during the COVID-19 pandemic: a potential public health crisis? Lancet Public Health. 2020;5:e259.
Koob GF, Powell P, White A. Addiction as a Coping Response: Hyperkatifeia, Deaths of Despair, and COVID-19. Am J Psychiatry. 2020;177:1031–7.
Spanagel R, Noori HR, Heilig M. Stress and alcohol interactions: animal studies and clinical significance. Trends Neurosci. 2014;37:219–27.
Fouyssac M, Puaud M, Ducret E, Marti-Prats L, Vanhille N, Ansquer S, et al. Environment-dependent behavioral traits and experiential factors shape addiction vulnerability. Eur J Neurosci. 2021;53:1794–808.
Clay JM, Parker MO. The role of stress-reactivity, stress-recovery and risky decision-making in psychosocial stress-induced alcohol consumption in social drinkers. Psychopharmacol (Berl). 2018;235:3243–57.
Khantzian EJ. Addiction as a self-regulation disorder and the role of self-medication. Addiction. 2013;108:668–9.
Giuliano C, Puaud M, Cardinal RN, Belin D, Everitt BJ. Individual differences in the engagement of habitual control over alcohol seeking predict the development of compulsive alcohol seeking and drinking. Addict Biol. 2021;26:e13041.
Giuliano C, Belin D, Everitt BJ. Compulsive Alcohol Seeking Results from a Failure to Disengage Dorsolateral Striatal Control over Behavior. J Neurosci. 2019;39:1744–54.
Lipton DM, Gonzales BJ, Citri A. Dorsal Striatal Circuits for Habits, Compulsions and Addictions. Front Syst Neurosci. 2019;13:28.
Li TK, Spanagel R, Colombo G, McBride WJ, Porrino LJ, Suzuki T, et al. Alcohol reinforcement and voluntary ethanol consumption. Alcohol Clin Exp Res. 2001;25:117S–126S.
Vollstadt-Klein S, Wichert S, Rabinstein J, Buhler M, Klein O, Ende G, et al. Initial, habitual and compulsive alcohol use is characterized by a shift of cue processing from ventral to dorsal striatum. Addiction. 2010;105:1741–9.
Robbins TW, Koob GF. Selective disruption of displacement behaviour by lesions of the mesolimbic dopamine system. Nature. 1980;285:409–12.
Robbins TW, Roberts DC, Koob GF. Effects of d-amphetamine and apomorphine upon operant behavior and schedule-induced licking in rats with 6-hydroxydopamine-induced lesions of the nucleus accumbens. J Pharm Exp Ther. 1983;224:662–73.
Ibias J, Soria-Molinillo E, Kastanauskaite A, Orgaz C, DeFelipe J, Pellon R, et al. Schedule-induced polydipsia is associated with increased spine density in dorsolateral striatum neurons. Neuroscience. 2015;300:238–45.
Murray JE, Belin D, Everitt BJ. Double dissociation of the dorsomedial and dorsolateral striatal control over the acquisition and performance of cocaine seeking. Neuropsychopharmacology. 2012;37:2456–66.
Corbit LH, Nie H, Janak PH. Habitual alcohol seeking: time course and the contribution of subregions of the dorsal striatum. Biol Psychiatry. 2012;72:389–95.
Fouyssac M, Pena-Oliver Y, Puaud M, Lim NTY, Giuliano C, Everitt BJ, et al. Negative Urgency Exacerbates Relapse to Cocaine Seeking After Abstinence. Biol Psychiatry. 2022;91:1051–60.
Hodebourg R, Murray JE, Fouyssac M, Puaud M, Everitt BJ, Belin D. Heroin seeking becomes dependent on dorsal striatal dopaminergic mechanisms and can be decreased by N-acetylcysteine. Eur J Neurosci. 2019;50:2036–44.
Murray JE, Belin-Rauscent A, Simon M, Giuliano C, Benoit-Marand M, Everitt BJ, et al. Basolateral and central amygdala differentially recruit and maintain dorsolateral striatum-dependent cocaine-seeking habits. Nat Commun. 2015;6:10088.
Vanderschuren LJ, Di Ciano P, Everitt BJ. Involvement of the dorsal striatum in cue-controlled cocaine seeking. J Neurosci. 2005;25:8665–70.
Zapata A, Minney VL, Shippenberg TS. Shift from goal-directed to habitual cocaine seeking after prolonged experience in rats. J Neurosci. 2010;30:15457–63.
Belin D, Belin-Rauscent A, Murray JE, Everitt BJ. Addiction: failure of control over maladaptive incentive habits. Curr Opin Neurobiol. 2013;23:564–72.
Marti-Prats L, Belin-Rauscent A, Fouyssac M, Puaud M, Cocker PJ, Everitt BJ, et al. Baclofen decreases compulsive alcohol drinking in rats characterized by reduced levels of GAT-3 in the central amygdala. Addict Biol. 2021;26:e13011.
Ansquer S, Belin-Rauscent A, Dugast E, Duran T, Benatru I, Mar AC, et al. Atomoxetine decreases vulnerability to develop compulsivity in high impulsive rats. Biol Psychiatry. 2014;75:825–32.
Pellon R, Ruiz A, Moreno M, Claro F, Ambrosio E, Flores P. Individual differences in schedule-induced polydipsia: neuroanatomical dopamine divergences. Behav Brain Res. 2011;217:195–201.
Moreno M, Gutierrez-Ferre VE, Ruedas L, Campa L, Sunol C, Flores P. Poor inhibitory control and neurochemical differences in high compulsive drinker rats selected by schedule-induced polydipsia. Psychopharmacol (Berl). 2012;219:661–72.
Moreno M, Flores P. Schedule-induced polydipsia as a model of compulsive behavior: neuropharmacological and neuroendocrine bases. Psychopharmacol (Berl). 2012;219:647–59.
Suzdak PD, Paul SM. Ethanol stimulates GABA receptor-mediated Cl- ion flux in vitro: possible relationship to the anxiolytic and intoxicating actions of alcohol. Psychopharmacol Bull. 1987;23:445–51.
Velazquez-Sanchez C, Muresan L, Marti-Prats L, Belin D. The development of compulsive coping behaviour is associated with a downregulation of Arc in a Locus Coeruleus neuronal ensemble. Neuropsychopharmacology. 2023;48:653–63.
Fouyssac M, Puaud M, Ducret E, Marti-Prats L, Vanhille N, Ansquer S et al. Environment-dependent behavioral traits and experiential factors shape addiction vulnerability. Eur J Neurosci. 2021;53:1794–808.
Killeen PR, Pellon R. Adjunctive behaviors are operants. Learn Behav. 2013;41:1–24.
Platt B, Beyer CE, Schechter LE, Rosenzweig-Lipson S. Schedule-induced polydipsia: a rat model of obsessive-compulsive disorder. Curr Protoc Neurosci. 2008; Chapter 9: Unit 9 27 https://doi.org/10.1002/0471142301.ns0927s43.
Woods A, Smith C, Szewczak M, Dunn RW, Cornfeldt M, Corbett R. Selective serotonin re-uptake inhibitors decrease schedule-induced polydipsia in rats: a potential model for obsessive compulsive disorder. Psychopharmacol (Berl). 1993;112:195–8.
Navarro SV, Gutierrez-Ferre V, Flores P, Moreno M. Activation of serotonin 5-HT2A receptors inhibits high compulsive drinking on schedule-induced polydipsia. Psychopharmacol (Berl). 2015;232:683–97.
Koob G. Neurobiological substrates for the dark side of compulsivity in addiction. Neuropharmacology. 2009;56:18–31.
Ford MM, Steele AM, McCracken AD, Finn DA, Grant KA. The relationship between adjunctive drinking, blood ethanol concentration and plasma corticosterone across fixed-time intervals of food delivery in two inbred mouse strains. Psychoneuroendocrinology. 2013;38:2598–610.
Meisch RA. The function of schedule-induced polydipsia in establishing ethanol as a positive reinforcer. Pharm Rev. 1975;27:465–73.
Ibias J, Pellon R. Schedule-induced polydipsia in the spontaneously hypertensive rat and its relation to impulsive behaviour. Behav Brain Res. 2011;223:58–69.
Yin HH, Knowlton BJ. The role of the basal ganglia in habit formation. Nat Rev Neurosci. 2006;7:464–76.
Yin HH, Knowlton BJ, Balleine BW. Lesions of dorsolateral striatum preserve outcome expectancy but disrupt habit formation in instrumental learning. Eur J Neurosci. 2004;19:181–9.
Dickinson A. Actions and Habits: The Development of Behavioural Autonomy. Philos Trans R Soc Lond. 1985;B 308:67–78.
Colotla VA. Adjunctive polydipsia as a model of alcoholism. Neurosci Biobehav Rev. 1981;5:335–42.
Guinle MIB, Sinha R. The Role of Stress, Trauma, and Negative Affect in Alcohol Misuse and Alcohol Use Disorder in Women. Alcohol Res. 2020;40:05.
Temmen CD, Crockett LJ. Relations of Stress and Drinking Motives to Young Adult Alcohol Misuse: Variations by Gender. J Youth Adolesc. 2020;49:907–20.
Katovic NM, Gresack JE, Spear LP. Schedule-induced polydipsia: gender-specific effects and consequences of prenatal cocaine and postnatal handling. Pharm Biochem Behav. 1999;64:695–704.
Stohr T, Szuran T, Welzl H, Pliska V, Feldon J, Pryce CR. Lewis/Fischer rat strain differences in endocrine and behavioural responses to environmental challenge. Pharm Biochem Behav. 2000;67:809–19.
Martin JR, Baettig K. Schedule induced ethanol polydipsia in psychogenetically selected lines of rats. Pharm Biochem Behav. 1981;14:857–62.
Ahmadi L, Goldman MB. Primary polydipsia: Update. Best Pr Res Clin Endocrinol Metab. 2020;34:101469.
Ibias J, Pellon R, Sanabria F. A microstructural analysis of schedule-induced polydipsia reveals incentive-induced hyperactivity in an animal model of ADHD. Behav Brain Res. 2015;278:417–23.
Alvarez B, Ibias J, Pellon R. Reinforcement of schedule-induced drinking in rats by lick-contingent shortening of food delivery. Learn Behav. 2016;44:329–39.
Schulz A, Vögele C. Interoception and stress. Front Psychol. 2015;6:993.
Cole BJ, Koob GF. Corticotropin-releasing factor and schedule-induced polydipsia. Pharm Biochem Behav. 1994;47:393–8.
Liu YP, Wang HC, Tseng CJ, Tang HS, Yin TH, Tung CS. Effects of amphetamine on schedule-induced polydipsia. Chin J Physiol. 2005;48:176–86.
Dickinson A, Dawson GR. Incentive learning and the motivational control of instrumental performance. Q J ExperimentalPsychol. 1989;41:99–112.
Belin D, Everitt BJ. Cocaine seeking habits depend upon dopamine-dependent serial connectivity linking the ventral with the dorsal striatum. Neuron. 2008;57:432–41.
Mittleman G, Whishaw IQ, Jones GH, Koch M, Robbins TW. Cortical, hippocampal, and striatal mediation of schedule-induced behaviors. Behav Neurosci. 1990;104:399–409.
Wallace M, Singer G, Finlay J, Gibson S. The effect of 6-OHDA lesions of the nucleus accumbens septum on schedule-induced drinking, wheelrunning and corticosterone levels in the rat. Pharm Biochem Behav. 1983;18:129–36.
Koob GF, Riley SJ, Smith SC, Robbins TW. Effects of 6-hydroxydopamine lesions of the nucleus accumbens septi and olfactory tubercle on feeding, locomotor activity, and amphetamine anorexia in the rat. J Comp Physiol Psychol. 1978;92:917–27.
Weissenborn R, Blaha CD, Winn P, Phillips AG. Schedule-induced polydipsia and the nucleus accumbens: electrochemical measurements of dopamine efflux and effects of excitotoxic lesions in the core. Behav Brain Res. 1996;75:147–58.
Mittleman G, Call SB, Cockroft JL, Goldowitz D, Matthews DB, Blaha CD. Dopamine dynamics associated with, and resulting from, schedule-induced alcohol self-administration: analyses in dopamine transporter knockout mice. Alcohol. 2011;45:325–39.
Leo D, Sukhanov I, Zoratto F, Illiano P, Caffino L, Sanna F, et al. Pronounced Hyperactivity, Cognitive Dysfunctions, and BDNF Dysregulation in Dopamine Transporter Knock-out Rats. J Neurosci. 2018;38:1959–72.
Jones GH, Hooks MS, Juncos JL, Justice JB Jr. Effects of cocaine microinjections into the nucleus accumbens and medial prefrontal cortex on schedule-induced behaviour: comparison with systemic cocaine administration. Psychopharmacol (Berl). 1994;115:375–82.
Nelson A, Killcross S. Amphetamine exposure enhances habit formation. J Neurosci. 2006;26:3805–12.
Hawken ER, Beninger RJ. The amphetamine sensitization model of schizophrenia symptoms and its effect on schedule-induced polydipsia in the rat. Psychopharmacol (Berl). 2014;231:2001–8.
Murray RM, Lappin J, Di Forti M. Schizophrenia: from developmental deviance to dopamine dysregulation. Eur Neuropsychopharmacol. 2008;18:S129–134.
Robbins TW, Costa RM. Habits. Curr Biol. 2017;27:R1200–R1206.
Gregory JG, Hawken ER, Banasikowski TJ, Dumont EC, Beninger RJ. A response strategy predicts acquisition of schedule-induced polydipsia in rats. Prog Neuropsychopharmacol Biol Psychiatry. 2015;61:37–43.
Merchan A, Mora S, Gago B, Rodriguez-Ortega E, Fernandez-Teruel A, Puga JL, et al. Excessive habit formation in schedule-induced polydipsia: Microstructural analysis of licking among rat strains and involvement of the orbitofrontal cortex. Genes Brain Behav. 2019;18:e12489.
Chamberlain SR, Blackwell AD, Fineberg NA, Robbins TW, Sahakian BJ. The neuropsychology of obsessive compulsive disorder: the importance of failures in cognitive and behavioural inhibition as candidate endophenotypic markers. Neurosci Biobehav Rev. 2005;29:399–419.
Everitt BJ, Robbins TW. Neural systems of reinforcement for drug addiction: from actions to habits to compulsion. Nat Neurosci. 2005;8:1481–9.
Gillan CM, Morein-Zamir S, Urcelay GP, Sule A, Voon V, Apergis-Schoute AM, et al. Enhanced avoidance habits in obsessive-compulsive disorder. Biol Psychiatry. 2014;75:631–8.
Voon V, Derbyshire K, Ruck C, Irvine MA, Worbe Y, Enander J, et al. Disorders of compulsivity: a common bias towards learning habits. Mol Psychiatry. 2015;20:345–52.
Schwabe L, Wolf OT. Stress prompts habit behavior in humans. J Neurosci. 2009;29:7191–8.
Smeets T, van Ruitenbeek P, Hartogsveld B, Quaedflieg C. Stress-induced reliance on habitual behavior is moderated by cortisol reactivity. Brain Cogn. 2019;133:60–71.
Dias-Ferreira E, Sousa JC, Melo I, Morgado P, Mesquita AR, Cerqueira JJ, et al. Chronic stress causes frontostriatal reorganization and affects decision-making. Science. 2009;325:621–5.
Zorrilla EP, Koob GF. Impulsivity Derived From the Dark Side: Neurocircuits That Contribute to Negative Urgency. Front Behav Neurosci. 2019;13:136.
Packard MG. Anxiety, cognition, and habit: a multiple memory systems perspective. Brain Res. 2009;1293:121–8.
Merchan A, Sanchez-Kuhn A, Prados-Pardo A, Gago B, Sanchez-Santed F, Moreno M, et al. Behavioral and biological markers for predicting compulsive-like drinking in schedule-induced polydipsia. Prog Neuropsychopharmacol Biol Psychiatry. 2019;93:149–60.
Navarro SV, Alvarez R, Colomina MT, Sanchez-Santed F, Flores P, Moreno M. Behavioral Biomarkers of Schizophrenia in High Drinker Rats: A Potential Endophenotype of Compulsive Neuropsychiatric Disorders. Schizophr Bull. 2017;43:778–87.
Gremel CM, Lovinger DM. Associative and sensorimotor cortico-basal ganglia circuit roles in effects of abused drugs. Genes Brain Behav. 2017;16:71–85.
Salinas AG, Mateo Y, Carlson VCC, Stinnett GS, Luo G, Seasholtz AF, et al. Long-term alcohol consumption alters dorsal striatal dopamine release and regulation by D2 dopamine receptors in rhesus macaques. Neuropsychopharmacology. 2021;46:1432–41.
Porrino LJ, Lyons D, Smith HR, Daunais JB, Nader MA. Cocaine self-administration produces a progressive involvement of limbic, association, and sensorimotor striatal domains. J Neurosci. 2004;24:3554–62.
Renteria R, Baltz ET, Gremel CM. Chronic alcohol exposure disrupts top-down control over basal ganglia action selection to produce habits. Nat Commun. 2018;9:211.
Corbit LH, Janak PH. Habitual Alcohol Seeking: Neural Bases and Possible Relations to Alcohol Use Disorders. Alcohol Clin Exp Res. 2016;40:1380–9.
Barker JM, Corbit LH, Robinson DL, Gremel CM, Gonzales RA, Chandler LJ. Corticostriatal circuitry and habitual ethanol seeking. Alcohol. 2015;49:817–24.
Patton MH, Roberts BM, Lovinger DM, Mathur BN. Ethanol Disinhibits Dorsolateral Striatal Medium Spiny Neurons Through Activation of A Presynaptic Delta Opioid Receptor. Neuropsychopharmacology. 2016;41:1831–40.
Wilcox MV, Cuzon Carlson VC, Sherazee N, Sprow GM, Bock R, Thiele TE, et al. Repeated binge-like ethanol drinking alters ethanol drinking patterns and depresses striatal GABAergic transmission. Neuropsychopharmacology. 2014;39:579–94.
Koob GF. Negative reinforcement in drug addiction: the darkness within. Curr Opin Neurobiol. 2013;23:559–63.
Hutcheson DM, Everitt BJ, Robbins TW, Dickinson A. The role of withdrawal in heroin addiction: enhances reward or promotes avoidance? Nat Neurosci. 2001;4:943–7.
Breese GR, Chu K, Dayas CV, Funk D, Knapp DJ, Koob GF, et al. Stress enhancement of craving during sobriety: a risk for relapse. Alcohol Clin Exp Res. 2005;29:185–95.
Koob GF, Le Moal M. Drug addiction, dysregulation of reward, and allostasis. Neuropsychopharmacology. 2001;24:97–129.
Larimer ME, Palmer RS, Marlatt GA. Relapse prevention. An overview of Marlatt’s cognitive-behavioral model. Alcohol Res health : J Natl Inst Alcohol Abus Alcohol. 1999;23:151–60.
Lowman C, Allen J, Stout RL, Group TRR. Replication and extension of Marlatt’s taxonomy of relapse precipitants: overview of procedures and results. Addiction. 1996;91:51–72.
Marlatt GA. Taxonomy of high-risk situations for alcohol relapse: evolution and development of a cognitive-behavioral model. Addiction. 1996;91:S37–49.
Wilson GT. Alcohol and anxiety. Behav Res Ther. 1988;26:369–81.
Moberg CA, Curtin JJ. Alcohol selectively reduces anxiety but not fear: startle response during unpredictable versus predictable threat. J Abnorm Psychol. 2009;118:335–47.
Spanagel R, Montkowski A, Allingham K, Stohr T, Shoaib M, Holsboer F, et al. Anxiety: a potential predictor of vulnerability to the initiation of ethanol self-administration in rats. Psychopharmacol (Berl). 1995;122:369–73.
George DT, Nutt DJ, Dwyer BA, Linnoila M. Alcoholism and panic disorder: is the comorbidity more than coincidence? Acta Psychiatr Scand. 1990;81:97–107.
Pellon R, Ruiz A, Lamas E, Rodriguez C. Pharmacological analysis of the effects of benzodiazepines on punished schedule-induced polydipsia in rats. Behav Pharm. 2007;18:81–87.
Balleine B. Instrumental performance following a shift in primary motivation depends on incentive learning. J Exp Psychol Anim Behav Process. 1992;18:236–50.
Paxinos G, Watson C. The rat brain in stereotaxic coordinates, 7th ed. 7th edn. San Diego: Elesvier Academic Press; 2013.
Acknowledgements
This work, carried out at the department of Psychology of the University of Cambridge, was funded by a UKRI grant to DB (MR/W019647/1), a UKRI grant (MR/N02530X/1) to BJE and DB, a Brain and Behavior Research Foundation NARSAD Young Investigator Grant (RG95599 Grant ID: 27353) to CG, a Leverhulme Trust Early Career (ECF-2018-713) and Isaac Newton Trust fellowship (18.08(g)) to LMP, and a Leverhulme Trust Early Career (ECF-2021-451) and Isaac Newton Trust fellowship (21.08(g)) to MF. For the purpose of open access, the author has applied a Creative Commons Attribution (CC BY) licence to any Author Accepted Manuscript version arising. The data can be available at: https://drive.google.com/drive/folders/1_qrEP-ztVDG3IOMR6Ytza5cKnCbbVvzn?usp=sharing We authors would like to thank Drs Aude Belin-Rauscent, Antonio Ferragud and Clara Velazquez-Sanchez, as well as Laetitia Ward, for technical assistance with behavioral training.
Author information
Authors and Affiliations
Contributions
DB, CG, AD, YPO and LMP designed the experiments. CG, AD, YPO, MP, CM and LMP carried out the behavioral experiments. LMP, CG, MF and DB performed the data analysis. LMP, CG, BJE and DB wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Marti-Prats, L., Giuliano, C., Domi, A. et al. The development of compulsive coping behavior depends on dorsolateral striatum dopamine-dependent mechanisms. Mol Psychiatry 28, 4666–4678 (2023). https://doi.org/10.1038/s41380-023-02256-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41380-023-02256-z
This article is cited by
-
From compulsivity to compulsion: the neural basis of compulsive disorders
Nature Reviews Neuroscience (2024)
