
Abstract
The glutamatergic neurotransmission system and the N-methyl-D-aspartate receptor (NMDAR) have been implicated in smoking and alcohol consumption behavior. Preclinical studies have demonstrated that nicotine and ethanol influence NMDAR functionality, which may have a role in tendencies to consume these substances. Nonetheless, little is known about concentrations of NMDAR coagonists in the cerebrospinal fluid (CSF) and plasma of individuals who smoke or consume alcohol. Glycine and L- and D-stereoisomers of alanine, serine, and proline were therefore measured using ultra-high-performance liquid chromatography-tandem mass spectrometry in 403 healthy subjects. Nicotine and alcohol consumption were quantified using questionnaires. Possible differences in NMDAR coagonist concentrations in plasma and CSF were investigated using ANCOVA with age, body mass index, and storage duration as covariates. The significance threshold was Bonferroni corrected (α=0.00625). Compared with non-smokers, smokers displayed lower levels of D-proline in plasma (p=0.0027, Cohen’s d=−0.41) and D-proline in CSF (p=0.0026, Cohen’s d=−0.43). D-Serine in CSF was higher in smokers than in non-smokers (p=0.0052, Cohen’s d=0.41). After subdividing participants based on smoking quantity, dose-dependent decreases were demonstrated in smokers for D-proline in plasma (F=5.65, p=0.0039) and D-proline in CSF (F=5.20, p=0.0060). No differences in NMDAR coagonist levels between alcohol consumption groups were detected. To our knowledge, this is the first report to implicate D-amino acids in smoking behavior of humans. Whether such concentration differences lie at the root of or result from smoking habits may be addressed in prospective studies.
Similar content being viewed by others
INTRODUCTION
In Europe, the prevalence of smoking was ∼28.6% in 2005 (WHO, 2007, 2012). Smoking decreases life expectancy by 12–20 years and is one of the major mortality risk factors in the world: more than 5 million people worldwide die of the consequences of tobacco consumption yearly (WHO, 2012).
Although genetic variants in genes encoding neuronal nicotinic acetylcholine receptor subunits (eg, CHRNA3 and CHRNA5) are associated with smoking quantity, the explained variances by such single-nucleotide polymorphisms are low (Tobacco and Genetics Consortium, 2010; Liu et al, 2010). This suggests that other pathways, such as the glutamatergic neurotransmission system, have a role in smoking behavior. Glutamate is the prime excitatory neurotransmitter in the central nervous system (CNS) and binds to several receptors, including the ionotropic N-methyl-D-aspartate receptor (NMDAR). Animal studies have reported that nicotine increases glutamate concentrations in the ventral tegmental area (VTA) (Fu et al, 2000; Schilstrom et al, 2000) and exerts an excitatory effect on the NMDAR in dopaminergic neurons in the VTA (Fu et al, 2000; Grillner and Svensson, 2000; Mansvelder and McGehee, 2000), the nucleus accumbens (Schilstrom et al, 1998), and the central nucleus of the amygdala (Kenny et al, 2009). In addition, nicotine self-administration in rats upregulates the expression of NMDAR subunits in the VTA and amygdala (Kenny et al, 2009). In vivo blockade of the NMDAR by NMDAR antagonists diminishes nicotine-induced dopamine release (Schilstrom et al, 1998) and nicotine self-administration (Kenny et al, 2009), but promotes tolerance to nicotine administration in rats (Shoaib et al, 1994; Shoaib and Stolerman, 1992). NMDAR antagonism could therefore constitute a target in the treatment of nicotine dependence, although to date this has not been successful in humans (Liechti and Markou, 2008).
The other substance frequently associated with burden of disease in Western society is alcohol. The World Health Organization estimated that alcohol is currently the world’s third largest risk factor for burden of disease (WHO, 2009). In contrast to nicotine, ethanol has been reported to block the NMDAR (Hoffman et al, 1989; Little, 1991; Lovinger et al, 1989; Weight et al, 1991). Chronic alcohol administration increases the number of NMDARs, thus playing a role in alcohol dependence and withdrawal seizures (Grant et al, 1990; Gulya et al, 1991; Lovinger et al, 1989). Moreover, NMDAR antagonists (eg, MK-801 and ketamine) cause ethanol-like effects in animals and humans (Hundt et al, 1998; Kotlinska and Liljequist, 1997; Krystal et al, 1998).
Glycine, D-serine, D-alanine, and L-proline are coagonists at the NMDAR (Henneberger et al, 2010; Henzi et al, 1992; Kleckner and Dingledine, 1988; Martin et al, 1992; Matsui et al, 1995; McBain et al, 1989; Mothet et al, 2000; Nong et al, 2003; Panatier et al, 2006; Sakata et al, 1999; Snyder and Kim, 2000). In particular, D-serine and glycine were demonstrated to strongly impact synaptic and extrasynaptic NMDAR signaling, whereas manipulation of both these coagonists has yielded a more nuanced understanding of NMDAR functionality (Papouin et al, 2012). To date, neither in plasma nor cerebrospinal fluid (CSF) have NMDAR coagonists been investigated for their role in smoking or alcohol consumption tendencies. Elucidating possible substance use-associated abnormalities in NMDAR coagonist levels may deepen the understanding of NMDAR involvement in substance use and susceptibility to nicotine and alcohol dependence. Moreover, the success rates of pharmacotherapies in the treatment of nicotine (Eisenberg et al, 2008) and alcohol (Rosner et al, 2008) dependence are limited and therefore novel agents may contribute to treatment optimization. Elucidating NMDAR coagonist functionality in subjects who consume these substances will potentially set the stage for further research into pharmacological modulations of NMDAR gating in preclinical settings and human consumers.
As the NMDAR is involved in smoking and alcohol use, we hypothesized that consumers of nicotine and alcohol display altered levels of NMDAR coagonists in CSF and plasma. Given the demonstrated increase in glutamate concentrations in the rat brain and upregulation of the NDMAR after nicotine administration (Grillner and Svensson, 2000; Mansvelder and McGehee, 2002; Schilstrom et al, 2000), we postulated that coagonist levels are elevated in smokers. As ethanol blocks the NMDAR (Hoffman et al, 1989; Little, 1991; Lovinger et al, 1989; Weight et al, 1991), we expected NMDAR coagonists in alcohol consumers to be decreased. Enantiomers of the above-mentioned coagonists were included in the current study as synthesis and degradation of stereoisomers are likely to be interdependent. We thus measured concentrations of glycine (that is not chiral) and the enantiomers of alanine, serine, and proline in a unique study population with CSF and plasma available for 403 subjects. We then compared these levels across categories of nicotine and alcohol consumption to detect possible concentration differences associated with the number of cigarettes or alcoholic beverages consumed.
MATERIALS AND METHODS
Subjects
Subject recruitment has been described in detail previously (Luykx et al, 2013). In brief, from August 2008 to November 2011, 403 subjects were recruited at outpatient preoperative screening services in and around Utrecht, the Netherlands. At these services, subjects are advised by the anesthesiologist to start fasting at least 6 h preoperatively and refrain from smoking and alcoholic beverages at least 24 h before the procedure (no biochemical verification). This enabled us to study sustained effects—ie, effects persisting for at least 24 h—of smoking and alcohol on amino-acid concentrations. We included patients (i) undergoing spinal anesthesia for minor elective surgical procedures, (ii) ranging between 18 and 60 years of age, and (iii) with four grandparents born in the Netherlands or other North-Western European countries (Belgium, Germany, the UK, France, and Denmark). Each candidate participant received a personal telephone interview to exclude subjects with psychotic or major neurological disorders (stroke, brain tumors, neurodegenerative diseases) and to record any self-reported history of other psychiatric illness or any use of psychotropic medication. Informed consent was obtained from the participants and the Ethics Committee of the UMCU and all local ethics committees approved the study.
CSF and Plasma Collection and Chemical Analyses
Whole blood was collected in EDTA tubes for plasma extraction. Plasma was extracted by centrifuging whole blood at ambient temperature for 10 min at 2500 g, after which plasma was stored at −80 °C. The standardized operating procedures adopted to collect 6 ml of CSF from each subject were described previously (Luykx et al, 2012). Chemical analyses of the L- and D-isomers of alanine, serine, proline, and glycine (no D-isomer) were conducted using ultra-high-performance liquid chromatography-tandem mass spectrometry according to a validated method (Visser et al, 2011).
Questionnaires
During a 2-week period after the elective procedure, subjects filled out online questionnaires about their health. The following questions regarding current smoking and alcohol consumption habits were asked:
-
Do you smoke? If so, how many cigarettes do you smoke per day?
-
Do you drink alcohol? If so, how many units of alcohol do you drink per day?
Smokers were asked to choose between the following divisions of number of cigarettes smoked per day: <1; 1–10; 11–20; 21–30; and >30. Alcohol consumption was quantified using the following cutoffs: <1; 1–3; 4–6; and >6 alcoholic beverages.
We chose web-based symptom questionnaires as these have been validated as reliable assessment tools in a range of epidemiological studies (Ekman et al, 2006; Gosling et al, 2004) and may decrease socially desirable responses compared with face-to-face interviews or questionnaires that are filled out in clinical settings (Buchanan and Smith, 1999; Joinson, 1999).
Statistical Analyses
Regarding smoking habits, the participants were divided into two groups: non-smokers (no current smoking) and smokers (any number of cigarettes smoked daily). Alcohol users were divided into mild alcohol consumers (<1 alcohol unit/day) and moderate alcohol consumers (⩾1 alcohol units/day) as only 11.2% of the study population proved abstinent.
Outliers were defined as subjects having at least one measurement with 3 or more standard deviations (SDs) from the mean and were excluded from further analyses. As no generally accepted covariates for amino-acid measurements are available, we comprehensively assessed possible covariates during the study period. The following variables were tested: age, sex, the rostrocaudal concentration gradient (reflected by the participants’ height), lumbar puncture level (binary, ie, lumbar vertebrate levels ⩽3 vs >3 as estimated by the anesthesiologist), time elapsed before storage (continuous, in hours), time of the day of lumbar puncture (continuous, rounded to the half hour), storage duration until chemical analyses (continuous, in months), body mass index (BMI (continuous), and amount of CSF drawn (continuous, as in 8% of the cases >7 or <5 ml of CSF were drawn). Relevant covariates were defined as variables that correlated with more than one amino acid at a Spearman’s ρ p-value <0.05. If covariates were collinear (r>0.5), the covariate with most missing data was excluded from the model. Only for height and weight, data were missing (for 25 out of 403 subjects, ie, 6.2%) and replaced by the mean per sex (183.4 cm and 86.8 kg for men; 170.5 cm and 74.6 kg for women). Using ANOVA and χ2 tests possible smoking and alcohol consumption-dependent differences were assessed.
Normality of the distributions was verified with a Kolmogorov–Smirnov (K-S) test and defined as a two-tailed asymptotic p-value⩾0.05. Non-normally distributed amino acids were logarithm (log) transformed. Homogeneity of variances (defined as a Levene’s test p-value>0.05) and homogeneity of regression slopes (by visual inspection of the scatterplots between the amino acids and the covariates) between groups were verified. To unravel possible interaction effects of alcohol and smoking behavior on amino-acid levels, we applied a generalized linear model and tested possible interaction effects of the above-mentioned dichotomous smoking and alcohol traits on amino-acid concentrations (Bonferroni-corrected type III Wald χ2 α=0.05/8=0.00625).
A one-way ANCOVA was conducted correcting for all relevant covariates (defined above). Significance was Bonferroni corrected (α=0.05/8=0.00625, as four NMDAR coagonists were tested for both nicotine and alcohol consumption, whereas stereoisomers in CSF and plasma were highly correlated, Supplementary 1). Stratification by sex was not performed due to the relatively small number of female participants (N=99). In the event Bonferroni-corrected significance was attained, we tested whether concentration differences were substance dose dependent. To that end, covariates that correlated at p<0.05 with that amino acid were determined and ANCOVA correcting for these covariates (or ANOVA in the event no covariate correlated with that amino acid) was conducted for the following substance consumption categories: <1 consumption a day (category 1); 1–10 cigarettes or 1–3 alcoholic beverages a day (category 2); and >10 cigarettes or >3 alcoholic beverages a day (category 3). Given the small numbers of subjects smoking >20 cigarettes a day or consuming >4 alcoholic beverages daily, this subdivision resulted in the most equal numbers of subjects per category. All statistical analyses were conducted using SPSS version 20 (SPSS, Chicago, IL).
RESULTS
Subject and NMDAR Coagonist Characteristics
Information about smoking and alcohol consumption habits in addition to NMDAR coagonist levels in either CSF or plasma were available for 403 subjects. Exclusion of outliers (55 subjects) brought the study population to 348 subjects (249 men and 99 women). Characteristics of these 348 subjects (N=325 for whom CSF was available; N=307 for whom plasma was available) are summarized in Table 1. The only significant differences in subject characteristics across substance use categories were found for psychiatric comorbidity and psychotropic medication (both increased in smokers vs non-smokers; χ2 p-values of 0.030 and 0.018, respectively). Six NMDAR coagonists were normally distributed (L-serine and D-serine in CSF; L-alanine, glycine, D-serine, and L-proline in plasma; Supplementary 2). The other NMDAR coagonists were log transformed, resulting in a formally normal distribution by K-S testing for all concentrations except L-proline in CSF (that approximated normality upon visual inspection, Supplementary 2). Supplementary 3 presents the mean NMDAR coagonist concentrations parsed by substance use category and body fluid.
NMDAR Coagonist Concentrations in Smokers and Non-Smokers
The covariates age, BMI, and storage duration were the only covariates that correlated with >1 amino acid. These showed correlations (p<0.05) with several amino acids (Supplementary 4). For these covariates no data were missing. No interaction effects between alcohol and smoking habits on amino-acid levels were detected.
After correction for these three covariates, all D-enantiomers in plasma were lower in smokers than in non-smokers, but for plasma only D-proline reached Bonferroni-corrected significance: the D-proline concentration in plasma was lower in smokers than in non-smokers (F1,302=9.61, p=0.0027, Cohen’s d=−0.41; Table 2 and Figure 1a). The plasma D-alanine difference between smokers and non-smokers was nominally significant (p=0.021) and plasma D-serine was only slightly decreased in smokers (p=0.21).

D-Amino-acid concentration differences between smokers and non-smokers: interquartile ranges (boxes) with medians (lines in boxes), whiskers (10–90 percentiles), and dots (values falling outside the 10–90 percentiles). (a) D-Proline in plasma in smokers vs non-smokers. (b) D-Proline in CSF in smokers vs non-smokers. (c) D-Serine in CSF in smokers vs non-smokers.
D-Proline in CSF was also lower in smokers than in non-smokers (F1,320=9.22, p=0.0026, Cohen’s d=−0.43; Figure 1b), whereas D-serine in CSF was higher in smokers than in non-smokers (F1,320=7.91, p=0.0052, Cohen’s d=0.41; Figure 1c).
We then assessed whether the concentration differences were substance dose dependent for the Bonferroni-corrected significant results. Although for D-serine in CSF no smoking dose-dependent effect was detected, we found significant differences between the three categories for D-proline (Table 3):
-
D-Proline in plasma (covariates: age and BMI): F2,302=5.65, p=0.0039 (Figure 2a).
Figure 2 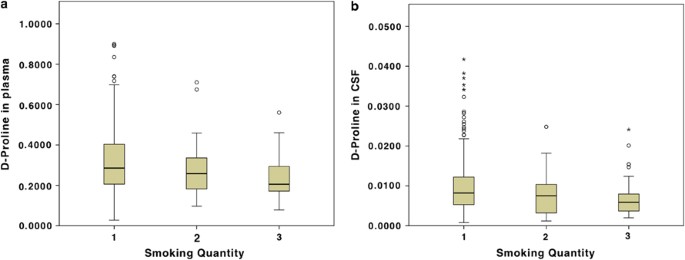
D-Proline in plasma (a) and D-proline in CSF (b) concentrations significantly differ by smoking quantity category (1=<1 cigarette; 2=1–10 cigarettes; 3=>10 cigarettes per day): interquartile ranges (boxes) with medians (lines in boxes), whiskers (10–90 percentiles), and dots (values falling outside the 10–90 percentiles).
-
D-Proline in CSF (covariate: age): F2,321=5.20, p=0.0060 (Figure 2b).

Finally, as psychiatric comorbidity and psychotropic medication significantly differed between smokers and non-smokers, we tested whether these variables were correlated with any of the above-mentioned amino acids. D-Serine in plasma correlated with psychiatric comorbidity (Spearman’s rho=0.16, p=0.005), but correcting for this variable did not change the results (data not shown). Variances and regression slopes were homogeneous between groups for all reported differences.
NMDAR Coagonist Concentrations in Mild and Moderate Alcohol Consumers
No differences in NMDAR coagonist levels between mild (<1 unit/day) and moderate alcohol consumers (⩾1 units/day) were detected (Supplementary 5).
Amino-Acid Reference Values
As reference values for most D- and L-amino acids in CSF and plasma are currently lacking or based on limited study populations, we provide these values before removal of any outliers to give an impression of their naturally occurring variation (Supplementary 6).
DISCUSSION
Here, we demonstrate an increase in CSF D-serine and a decrease in CSF and plasma D-proline in smokers compared with non-smokers (N=348). The D-proline concentration differences proved substance dose-dependent. No differences in NMDAR coagonists between mild and moderate alcohol consumers were found.
The current study being the first to comprehensively compare NMDAR-coagonist levels in the CSF and plasma of smoking and non-smoking subjects, our results cannot be directly compared with previous findings. To our knowledge, only in smoking pregnant women and smoking schizophrenia patients and controls have amino-acid levels been compared with non-smokers. None of these two studies detected smoking behavior-dependent differences in plasma amino-acid levels (Jauniaux et al, 1999; Clelland et al, 2011). However, instead of parsing the results by enantiomers, only total amino-acid concentrations were investigated in those studies. As the primary contribution to total amino-acid concentrations comes from L-enantiomers, such previous findings combined with the data presented here suggest that smoking may be associated only with D-enantiomers of amino acids.
A possible mechanism whereby smoking leads to D-amino acid aberrations relates to the direct effect of nicotine on the NMDAR. Nicotine-dependent activation of nicotinic acetylcholine receptors enhances glutamatergic transmission (Mansvelder and McGehee, 2000). Glutamate can then activate the NMDAR, leading to long-term potentiation (Mansvelder and McGehee, 2000). Conceivably, long-term potentiation may thereafter result in D-serine upregulation in the CNS as D-serine is the most important ligand at the synaptic NMDAR glycine-binding site (Papouin et al, 2012). Such increased D-serine consequential to nicotine effects in the CNS is in line with the high CSF D-serine concentrations we observed in smokers. Relatively high CSF D-serine in smokers is further consistent with a burgeoning body of evidence demonstrating lack of efficacy of D-cycloserine in reducing tobacco consumption (Kamboj et al, 2012; Nesic et al, 2011; Santa Ana et al, 2009; Yoon et al, 2013). Further potentiating D-serine-dependent NMDAR neurotransmission by D-cycloserine administration in smokers would likely be unsuccessful for this indication.
To our knowledge, no significant differences in NMDAR coagonist levels between mild and moderate alcohol consumers have been reported, although increased glutamate and proline levels in plasma of alcohol-dependent patients with positive alcohol breath tests have been reported (Walter et al, 2008). This suggests that alcohol consumption shortly before sampling affects glutamate and proline levels. The subjects included in the current study had fasted before lumbar puncture and were instructed to be abstinent for at least 24 h before lumbar puncture. Possibly, such previously reported effects of alcohol on proline concentrations are short lived. Higher proline levels in plasma in subjects with alcohol abuse or dependence compared with healthy controls have been described (Clelland et al, 2011). Our study population differed in that here analyses were performed on subjects from the general population–– not a patient–control cohort.
A limitation of this study is that no formal assessments of alcohol and nicotine dependence or other measures relevant to smoking habits (eg, vulnerability to relapse and smoking initiation) were conducted. Therefore, the effects of such diagnoses and behavioral characteristics on D-amino acid levels cannot be construed from our data. In addition, the data we provide are cross-sectional and therefore inferences about causality cannot be made. Furthermore, the limited number of study participants precluded us from running linear regression as few smokers fell in high consumption categories. Although designs quantitatively assessing smoking habits and measuring NMDAR coagonists in the plasma of a substantial number of participants are feasible, CSF would be a more cumbersome target. On a similar note, this study was underpowered to stratify analyses by sex given the small number of female subjects. Finally, whether nicotine-dependent influences on NMDAR coagonist levels are sustained over time cannot be deduced from our data, as measurements were not performed at different time points. However, as participants were advised to refrain from smoking at least 24 h preoperatively, it is possible that D-amino acid aberrations in smokers are not short lived.
In conclusion, the differences in D-amino acids in CSF and plasma that we detected support their involvement in smoking behavior. Animal studies may determine whether nicotine-dependent activation of the NMDAR is mediated by D-serine and D-proline. Future longitudinal designs in humans that incorporate measurements of D-amino acid levels before and after substance consumption may clarify whether differences in such levels are state or trait dependent. Moreover, given previously highlighted sex-dependent differences in smoking behavior (Townsend et al, 1994), upcoming experiments will hopefully attain sufficient power to stratify by sex. Furthermore, formally diagnosing nicotine dependence and assessments of additional smoking behavior measures (eg, vulnerability to relapse and smoking initiation) in such projects may tease apart associations of substance use phenotypes with D-amino acid aberrations.
References
Buchanan T, Smith JL (1999). Using the Internet for psychological research: personality testing on the World Wide Web. Br J Psychol 90: 125–144.
Clelland CL, Read LL, Baraldi AN, Bart CP, Pappas CA, Panek LJ et al (2011). Evidence for association of hyperprolinemia with schizophrenia and a measure of clinical outcome. Schizophr Res 131: 139–145.
Eisenberg MJ, Filion KB, Yavin D, Belisle P, Mottillo S, Joseph L et al (2008). Pharmacotherapies for smoking cessation: a meta-analysis of randomized controlled trials. Cmaj 179: 135–144.
Ekman A, Dickman PW, Klint A, Weiderpass E, Litton JE (2006). Feasibility of using web-based questionnaires in large population-based epidemiological studies. Eur J Epidemiol 21: 103–111.
Fu Y, Matta SG, Gao W, Brower VG, Sharp BM (2000). Systemic nicotine stimulates dopamine release in nucleus accumbens: re-evaluation of the role of N-methyl-D-aspartate receptors in the ventral tegmental area. J Pharmacol Exp Ther 294: 458–465.
Gosling SD, Vazire S, Srivastava S, John OP (2004). Should we trust web-based studies? A comparative analysis of six preconceptions about internet questionnaires. Am Psychol 59: 93–104.
Grant KA, Valverius P, Hudspith M, Tabakoff B (1990). Ethanol withdrawal seizures and the NMDA receptor complex. Eur J Pharmacol 176: 289–296.
Grillner P, Svensson TH (2000). Nicotine-induced excitation of midbrain dopamine neurons in vitro involves ionotropic glutamate receptor activation. Synapse 38: 1–9.
Gulya K, Grant KA, Valverius P, Hoffman PL, Tabakoff B (1991). Brain regional specificity and time-course of changes in the NMDA receptor-ionophore complex during ethanol withdrawal. Brain Res 547: 129–134.
Henneberger C, Papouin T, Oliet SH, Rusakov DA (2010). Long-term potentiation depends on release of D-serine from astrocytes. Nature 463: 232–236.
Henzi V, Reichling DB, Helm SW, MacDermott AB (1992). L-Proline activates glutamate and glycine receptors in cultured rat dorsal horn neurons. Mol Pharmacol 41: 793–801.
Hoffman PL, Rabe CS, Moses F, Tabakoff B (1989). N-Methyl-D-aspartate receptors and ethanol: inhibition of calcium flux and cyclic GMP production. J Neurochem 52: 1937–1940.
Hundt W, Danysz W, Holter SM, Spanagel R (1998). Ethanol and N-methyl-D-aspartate receptor complex interactions: a detailed drug discrimination study in the rat. Psychopharmacology (Berl) 135: 44–51.
Jauniaux E, Gulbis B, Acharya G, Gerlo E (1999). Fetal amino acid and enzyme levels with maternal smoking. Obstet Gynecol 93: 680–683.
Joinson A (1999). Social desirability, anonymity, and Internet-based questionnaires. Behav Res Methods Instrum Comput 31: 433–438.
Kamboj SK, Joye A, Das RK, Gibson AJ, Morgan CJ, Curran HV (2012). Cue exposure and response prevention with heavy smokers: a laboratory-based randomised placebo-controlled trial examining the effects of D-cycloserine on cue reactivity and attentional bias. Psychopharmacology (Berl) 221: 273–284.
Kenny PJ, Chartoff E, Roberto M, Carlezon WA Jr, Markou A (2009). NMDA receptors regulate nicotine-enhanced brain reward function and intravenous nicotine self-administration: role of the ventral tegmental area and central nucleus of the amygdala. Neuropsychopharmacology 34: 266–281.
Kleckner NW, Dingledine R (1988). Requirement for glycine in activation of NMDA-receptors expressed in Xenopus oocytes. Science 241: 835–837.
Kotlinska J, Liljequist S (1997). The NMDA/glycine receptor antagonist, L-701,324, produces discriminative stimuli similar to those of ethanol. Eur J Pharmacol 332: 1–8.
Krystal JH, Petrakis IL, Webb E, Cooney NL, Karper LP, Namanworth S et al (1998). Dose-related ethanol-like effects of the NMDA antagonist, ketamine, in recently detoxified alcoholics. Arch Gen Psychiatry 55: 354–360.
Liechti ME, Markou A (2008). Role of the glutamatergic system in nicotine dependence: implications for the discovery and development of new pharmacological smoking cessation therapies. CNS Drugs 22: 705–724.
Little HJ (1991). Mechanisms that may underlie the behavioural effects of ethanol. Prog Neurobiol 36: 171–194.
Liu JZ, Tozzi F, Waterworth DM, Pillai SG, Muglia P, Middleton L et al (2010). Meta-analysis and imputation refines the association of 15q25 with smoking quantity. Nat Genet 42: 436–440.
Lovinger DM, White G, Weight FF (1989). Ethanol inhibits NMDA-activated ion current in hippocampal neurons. Science 243: 1721–1724.
Luykx JJ, Bakker SC, Lentjes E, Neeleman M, Strengman E, Mentink L et al (2013). Genome-wide association study of monoamine metabolite levels in human cerebrospinal fluid. Mol Psychiatry e-pub ahead of print 15 January 2013. doi: 10.1038/mp.2012.183.
Luykx JJ, Vinkers CH, Bakker SC, Visser WF, van Boxmeer L, Strengman E et al (2012). A Common variant in ERBB4 regulates GABA concentrations in human cerebrospinal fluid. Neuropsychopharmacology 37: 2088–2092.
Mansvelder HD, McGehee DS (2000). Long-term potentiation of excitatory inputs to brain reward areas by nicotine. Neuron 27: 349–357.
Mansvelder HD, McGehee DS (2002). Cellular and synaptic mechanisms of nicotine addiction. J Neurobiol 53: 606–617.
Martin D, Ault B, Nadler JV (1992). NMDA receptor-mediated depolarizing action of proline on CA1 pyramidal cells. Eur J Pharmacol 219: 59–66.
Matsui T, Sekiguchi M, Hashimoto A, Tomita U, Nishikawa T, Wada K (1995). Functional comparison of D-serine and glycine in rodents: the effect on cloned NMDA receptors and the extracellular concentration. J Neurochem 65: 454–458.
McBain CJ, Kleckner NW, Wyrick S, Dingledine R (1989). Structural requirements for activation of the glycine coagonist site of N-methyl-D-aspartate receptors expressed in Xenopus oocytes. Mol Pharmacol 36: 556–565.
Mothet JP, Parent AT, Wolosker H, Brady RO Jr, Linden DJ, Ferris CD et al (2000). D-Serine is an endogenous ligand for the glycine site of the N-methyl-D-aspartate receptor. Proc Natl Acad Sci USA 97: 4926–4931.
Nesic J, Duka T, Rusted JM, Jackson A (2011). A role for glutamate in subjective response to smoking and its action on inhibitory control. Psychopharmacology (Berl) 216: 29–42.
Nong Y, Huang YQ, Ju W, Kalia LV, Ahmadian G, Wang YT et al (2003). Glycine binding primes NMDA receptor internalization. Nature 422: 302–307.
Panatier A, Theodosis DT, Mothet JP, Touquet B, Pollegioni L, Poulain DA et al (2006). Glia-derived D-serine controls NMDA receptor activity and synaptic memory. Cell 125: 775–784.
Papouin T, Ladepeche L, Ruel J, Sacchi S, Labasque M, Hanini M et al (2012). Synaptic and extrasynaptic NMDA receptors are gated by different endogenous coagonists. Cell 150: 633–646.
Rosner S, Leucht S, Lehert P, Soyka M (2008). Acamprosate supports abstinence, naltrexone prevents excessive drinking: evidence from a meta-analysis with unreported outcomes. J Psychopharmacol 22: 11–23.
Sakata K, Fukushima T, Minje L, Ogurusu T, Taira H, Mishina M et al (1999). Modulation by L- and D-isoforms of amino acids of the L-glutamate response of N-methyl-D-aspartate receptors. Biochemistry 38: 10099–10106.
Santa Ana EJ, Rounsaville BJ, Frankforter TL, Nich C, Babuscio T, Poling J et al (2009). D-Cycloserine attenuates reactivity to smoking cues in nicotine dependent smokers: a pilot investigation. Drug Alcohol Depend 104: 220–227.
Schilstrom B, Fagerquist MV, Zhang X, Hertel P, Panagis G, Nomikos GG et al (2000). Putative role of presynaptic alpha7* nicotinic receptors in nicotine stimulated increases of extracellular levels of glutamate and aspartate in the ventral tegmental area. Synapse 38: 375–383.
Schilstrom B, Nomikos GG, Nisell M, Hertel P, Svensson TH (1998). N-Methyl-D-aspartate receptor antagonism in the ventral tegmental area diminishes the systemic nicotine-induced dopamine release in the nucleus accumbens. Neuroscience 82: 781–789.
Shoaib M, Benwell ME, Akbar MT, Stolerman IP, Balfour DJ (1994). Behavioural and neurochemical adaptations to nicotine in rats: influence of NMDA antagonists. Br J Pharmacol 111: 1073–1080.
Shoaib M, Stolerman IP (1992). MK801 attenuates behavioural adaptation to chronic nicotine administration in rats. Br J Pharmacol 105: 514–515.
Snyder SH, Kim PM (2000). D-Amino acids as putative neurotransmitters: focus on D-serine. Neurochem Res 25: 553–560.
Tobacco and Genetics Consortium (2010). Genome-wide meta-analyses identify multiple loci associated with smoking behavior. Nat Genet 42: 441–447.
Townsend J, Roderick P, Cooper J (1994). Cigarette smoking by socioeconomic group, sex, and age: effects of price, income, and health publicity. BMJ 309: 923–927.
Visser WF, Verhoeven-Duif NM, Ophoff R, Bakker S, Klomp LW, Berger R et al (2011). A sensitive and simple ultra-high-performance-liquid chromatography-tandem mass spectrometry based method for the quantification of D-amino acids in body fluids. J Chromatogr A 1218: 7130–7136.
Walter H, Schlaff WB, Lesch OM, Vitek L, Zima T, Hartl D et al (2008). Breath alcohol level and plasma amino acids: a comparison between older and younger chronic alcohol-dependent patients. Alcohol Alcohol 43: 653–657.
Weight FF, Lovinger DM, White G, Peoples RW (1991). Alcohol and anesthetic actions on excitatory amino acid-activated ion channels. Ann N Y Acad Sci 625: 97–107.
WHO (2007) WHO Tobacco Control Report. World Health Organisation: Copenhaven, Denmark.
WHO (2009) WHO Global Health Risk. World Health Organisation: Geneva, Switserland.
WHO (2012) WHO World Health Statistics. World Health Organisation: Geneva, Switzerland.
Yoon JH, Newton TF, Haile CN, Bordnick PS, Fintzy RE, Culbertson C et al (2013). Effects of D-cycloserine on cue-induced craving and cigarette smoking among concurrent cocaine- and nicotine-dependent volunteers. Addict Behav 38: 1518–1526.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies the paper on the Neuropsychopharmacology website
PowerPoint slides
Rights and permissions
About this article
Cite this article
Luykx, J., Bakker, S., van Boxmeer, L. et al. D-Amino Acid Aberrations in Cerebrospinal Fluid and Plasma of Smokers. Neuropsychopharmacol 38, 2019–2026 (2013). https://doi.org/10.1038/npp.2013.103
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/npp.2013.103
Keywords
This article is cited by
-
Chiral capillary electrophoresis with UV-excited fluorescence detection for the enantioselective analysis of 9-fluorenylmethoxycarbonyl-derivatized amino acids
Analytical and Bioanalytical Chemistry (2018)
-
Determining the presence of asthma-related molecules and salivary contamination in exhaled breath condensate
Respiratory Research (2017)
-
Metabolomic similarities between bronchoalveolar lavage fluid and plasma in humans and mice
Scientific Reports (2017)
-
Evidence that COMT genotype and proline interact on negative-symptom outcomes in schizophrenia and bipolar disorder
Translational Psychiatry (2016)
-
Genome-wide association study of NMDA receptor coagonists in human cerebrospinal fluid and plasma
Molecular Psychiatry (2015)
