
Abstract
Bernard–Soulier syndrome (BSS) is a rare bleeding disorder of autosomal recessive inheritance characterized by macrothrombocytopenia. We report the case of a 14-year-old girl diagnosed with BSS who is a fourth-generation Brazilian of Japanese descent and has a compound heterozygote mutation as the responsible gene. The compound heterozygosity would have occurred from the global and long-term racial migration that brought about an accidental encounter of two rare mutant alleles of different origins.
Similar content being viewed by others
Bernard–Soulier syndrome (BSS, OMIM #231200), a rare congenital disorder of hemostasis, is marked by quantitative and functional abnormality of the platelets. It is characterized by moderate thrombocytopenia and giant platelets in a peripheral blood smear (markedly larger than red blood cells).1 The underlying cause is congenital defects in the glycoprotein (GP) Ib-IX-V complex, a platelet receptor for von Willebrand factor (vWF), impairing primary hemostasis. BSS has an autosomal recessive inheritance. The estimated frequency of homozygous cases is as low as one per million population based on case reports,2 and the frequency of the heterozygous genotype is 1 in 500 as projected using the Hardy–Weinberg equilibrium (1998/1 000 000).3 Several founder mutations with a distribution deflection have been identified worldwide.4 Herein, we describe a pediatric case of BSS with compound heterozygosity from different races, a fascinating example signifying historical population movements in molecular anthropology.
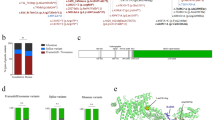
The patient is a 14-year-old girl whose mother is a third-generation Brazilian of Japanese descent (Figure 1a). There was no family history of bleeding tendency and her parents’ marriage was not consanguineous. At age 6, mild thrombocytopenia was indicated at a Brazilian hospital. She came to Japan at the age of 7; however, no medical investigations for thrombocytopenia were performed until age 12, when she informed our hospital, for the first time, about headaches and light-headedness. The symptoms had started just after initiation of menstruation, when she experienced excessive bleeding. Apart from conjunctival pallor, no other remarkable physical findings existed on initial examination. Blood examination revealed iron-deficient anemia, assumed to be due to hypermenorrhea. Her platelet count was moderately low (76×109/l) and giant platelets were observed in the peripheral blood smear. Bleeding time (Duke’s method) was mildly prolonged to 9 min (normal range, 2–5 min). vWF ristocetin cofactor activity was 92% (normal range, 60–170%). Platelet aggregation analysis could not be performed because of the unavailability of the test at our facility. Bone marrow examination performed 5 months after the first visit showed normal cellularity and no signs of dysplasia or megakaryocytosis (megakaryocytes 0.2%, 50/μl). No chromosomal abnormality was detected. Initial platelet flow cytometry (FCM) for platelet membrane glycoproteins by outsourcing showed an apparently normal percentage of positiveness as judged against the cutoff level in the negative control: GPIIb 97.4%, GPIIIa 99.0%, GPbα 95.8%, and GPIX 93.0% (data not shown). On the other hand, the results showed low mean fluorescence intensity (MFI) for GPIbα and GPIX. Repeat analysis revealed a significant decrease in MFI for GPIbα (20.0%) and GPIX (12.9%) compared with that in healthy persons. Regarding the GPIIb/IIIa analysis, the MFI was higher than the reference data that reflected the large surface area of giant platelets (278.6%; Figure 2). After obtaining approval from the ethical review boards of our facilities and the parents’ written informed consent, we performed DNA sequencing analysis of the patient and her asymptomatic parents (Figure 1b). All the coding exons for GP1BA, GP1BB, and GP9 were PCR amplified and directly sequenced by the Sanger method. The results showed that the patient was compound heterozygous for two missense mutations in GP9, NM_000174.3:c.182A>G and NM_000174.3:c.266G>A. Her father was a heterozygous carrier of the c.182A>G mutation, and her mother was a heterozygous carrier of the c.266G>A mutation. No mutations were found in GP1BA and GP1BB. A BSS diagnosis was genetically confirmed, and she began a monthly follow-up regime. Further, she was treated for iron-deficient anemia. Thereafter, no severe bleeding episode has occurred to date.

(a) Family tree. The proband is subject IV-2 with solid symbols. c.182A>G and c.266G>A in GP9 were transmitted individually from the parents (subject III-1,2), which indicates that the patient was compound heterozygous for these missense mutations. The patient’s parents are heterozygous and are shown by half-solid symbols. (b) DNA sequence analysis of GP9. c.182A>G and c.266G>A are predicted to lead to amino acid substitutions of asparagine to serine at codon 61 and of cysteine to tyrosine at codon 89, respectively.

FCM analysis of platelet GPs. The numbers in each panel indicate mean fluorescence intensity (MFI). Percentages indicate the MFI relative to that in normal controls. FCM, flow cytometric; GPs, glycoproteins.
BSS patients have congenital defects in the GPIb-IX-V complex, a platelet receptor for vWF, and present with severe bleeding caused by defective primary hemostasis. The GPIb-IX-V complex is composed of four polypeptide subunits, GPIbα, GPIbβ, GPIX, and GPV, that belong to the leucine-rich repeat family.5 A major ligand-binding domain exists within the N-terminal of GPIbα. With the exception of rare autosomal dominant cases,3 the disorder is usually inherited as an autosomal recessive trait. Many of patients are homozygous for mutations in one of the three genes: GP1BA (NM_000173.5), GP1BB (NM_000407.4), and GP9 (NM_000174.3).
Since the first report by Ware et al.,6 ~60 gene mutations that include missense, deletion, nonsense, and frameshift alterations have been detected in the three culprit genes.7 Without the concurrent expression of three genes, the GPIb-IX-V complex is not expressed efficiently on the platelet surface.8 Our patient with a low count of giant platelets had a significantly lower MFI for GPIbα and GPIX expressions than those in normal controls (Figure 2). Carriers may have slightly large platelets and decreased expression of GPIb-IX-V complex expression, even though they are usually asymptomatic with a normal number of platelets.2,3 In this case, GP expression of the parents, as assessed by FCM, showed slightly increased GpIIb, GPIIIa, and GPIIb/IIIa complex compared with that in normal controls, which is a common finding with giant platelets with a large surface area (paternal GPIIIa expression is the exception, and it will be discussed later). GPIbα and GPIX were expressed normally on the enlarged surface, which suggests that expression per unit area of the two GPs decreased in comparison with a normal control because they are heterozygous carriers.
We identified compound heterozygous GP9 mutations in the proband that consisted of c.182A>G resulting in p.Asn61Ser (N61S) substitution and c.266G>A resulting in p.Cys89Tyr (C89Y); both are already known to be causative mutations for BSS (Figure 1b).9–12
The former missense mutation occurs within the leucine-rich repeat motif, which disturbs the proper folding and reduces the stability of GPIX and ultimately prevents the expression of the GPIb-IX-V complex on the platelet surface.13
The N61S mutation was first described in 1993 by Write et al.9 and is shared by northern and central European Populations as a founder mutation.10,14,15
The patient’s father, who is a heterozygous for N61S, is also heterozygous for the human platelet antigen (HPA)-1ab. This was accidentally discovered by FCM, in which decreased expression of GPIIIa and normal expression of GPIIb and GPIIb/IIIa were observed. This is because the anti-GPIIIa antibody SZ21 used in FCM recognizes the HPA-1a epitope and thus the HPA-1b is not detected. HPA-1 genotyping revealed HPA-1a/b heterozygosity in the father (data not shown). Although HPA-1a is common in Japanese and Caucasian populations, the frequency of HPA-1b is different in the two races: extremely rare in Japanese, but relatively common (10–30%) in Caucasians.16 There is little information about the ancestors of the present case; however, it can be judged from these two facts that her father’s origin was European.
In contrast, maternal C89Y substitution, first described in 1996, is thought to be the Japanese founder mutation.11 The mutation causes a structural instability in the GPIX protein due to a substitution of cysteine necessary for intramolecular disulfide bonding of the protein. In vitro expression studies in heterologous cells implicated the mutation as the basis of BSS.12 A detailed interview revealed that her maternal great-grandfather was a resident of Japan’s Saga Prefecture and first-generation Brazilian (Figure 1a). Since 1908, approximately 130 000 Japanese people have immigrated to Brazil, which is now home to the largest population of Japanese descent outside of Japan.
The global and long-term movement of people, accompanied by genomes, has generated novel genetic encounters between individuals of totally different origins.
In conclusion, we described a girl with BSS who is a compound heterozygote for a combination of known founder mutations. Although this is a single case study, we suggest that the genetic interchange across races may change disease epidemiology. We should diagnose and treat hereditary diseases with a wide perspective.
References
References
Lopez JA, Andrews RK, Afshar-Kharghan V, Berndt MC . Bernard-Soulier syndrome. Blood 1998; 91: 4397–4418.
Savoia A, Balduini CL, Savino M, Noris P, Del Vecchio M, Perrotta S et al. Autosomal dominant macrothrombocytopenia in Italy is most frequently a type of heterozygous Bernard-Soulier syndrome. Blood 2001; 97: 1330–1335.
Noris P, Perrotta S, Bottega R, Pecci A, Melazzini F, Civaschi E et al. Clinical and laboratory features of 103 patients from 42 Italian families with inherited thrombocytopenia derived from the monoallelic Ala156Val mutation of gpib{alpha} (Bolzano mutation). Haematologica 2012; 97: 82–88.
Savoia A, Kunishima S, De Rocco D, Zieger B, Rand ML, Pujol-Moix N et al. Spectrum of the mutations in Bernard-Soulier syndrome. Hum Mutat 2014; 35: 1033–1045.
Andrews RK, Gardiner EE, Shen Y, Whisstock JC, Berndt MC . Glycoprotein Ib-IX-V. Int J Biochem Cell Biol 2003; 35: 1170–1174.
Ware J, Russell SR, Vicente V, Scharf RE, Tomer A, McMillan R et al. Nonsense mutation in the glycoprotein Ib alpha coding sequence associated with Bernard-Soulier syndrome. Proc Natl Acad Sci USA 1990; 87: 2026–2030.
Berndt MC, Andrews RK . Bernard-Soulier syndrome. Haematologica 2011; 96: 355–359.
Li R, Emsley J . The organizing principle of the platelet glycoprotein Ib-IX-V complex. J Thromb Haemost 2013; 11: 605–614.
Wright SD, Michaelides K, Johnson DJ, West NC, Tuddenham EG . Double heterozygosity for mutations in the platelet glycoprotein IX gene in three siblings with Bernard-Soulier syndrome. Blood 1993; 81: 2339–2347.
Sachs UJH, Kroll H, Matzdorff AC, Berghöfer H, Löpez JA, Santoso S . Bernard-Soulier syndrome due to the homozygous Asn-45Ser mutation in GPIX: an unexpected, frequent finding in Germany. Br J Haematol 2003; 123: 127–131.
Noda M, Fujimura K, Takafuta T, Shimomura T, Fujii T, Katsutani S et al. A point mutation in glycoprotein IX coding sequence (Cys73 (TGT) to Tyr(TAT)) causes impaired surface expression of GPIb/IX/V complex in two families with Bernard-Soulier syndrome. Thromb Haemost 1996; 76: 874–878.
Kunishima S, Tomiyama Y, Honda S, Kurata Y, Kamiya T, Ozawa K et al. Cys97-->Tyr mutation in the glycoprotein IX gene associated with Bernard-Soulier syndrome. Br J Haematol 1999; 107: 539–545.
Sae-Tung G, Dong JF, Lopez JA . Biosynthetic defect in platelet glycoprotein IX mutants associated with Bernard-Soulier syndrome. Blood 1996; 87: 1361–1367.
Liang HPH, Morel-Kopp M-C, Clemetson JM, Clemetson KJ, Kekomaki R, Kroll H et al. A common ancestral glycoprotein (GP) 9 1828A>G (Asn45Ser) gene mutation occurring in European families from Australia and Northern Europe with Bernard-Soulier Syndrome (BSS). Thromb Haemost 2005; 94: 599–605.
Koskela S, Javela K, Jouppila J, Juvonen E, Nyblom O, Partanen J et al. Variant Bernard-Soulier syndrome due to homozygous Asn45Ser mutation in the platelet glycoprotein (GP) IX in seven patients of five unrelated Finnish families. Eur J Haematol 1999; 62: 256–264.
Salem AH, Han K, Batzer MA . Allele frequencies of the human platelet antigen-1 in the Egyptian population. BMC Res Notes 2009; 2: 90.
Data Citations
Kanda, Kenji HGV Database (2017) http://dx.doi.org/10.6084/m9.figshare.hgv.1381
Kanda, Kenji HGV Database (2017) http://dx.doi.org/10.6084/m9.figshare.hgv.1384
Acknowledgements
We are grateful to the patient and her parents for their cooperation.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/4.0/
About this article

Cite this article
Kanda, K., Kunishima, S., Sato, A. et al. A Brazilian case of Bernard–Soulier syndrome with two distinct founder mutations. Hum Genome Var 4, 17030 (2017). https://doi.org/10.1038/hgv.2017.30
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/hgv.2017.30
