
Abstract
The effects of glucagon and epinephrine on gluconeogenesis in young (4 month) and old (24 month) Fisher 344 rat hepatocytes were compared. In contrast to glucagon, which had a similar effect on gluconeogenesis in both young and old cells, epinephrine caused a smaller increase in gluconeogenesis in old rat hepatocytes than in young hepatocytes. β2 adrenergic receptor (β2-AR) expression slightly decreased in aged rat liver, and there were differences between young and old hepatocytes in their patterns of G protein coupled receptor kinases, which are involved in the activation of β2-AR receptor signal desensitization. The major isoform of the kinase changed from GRK2 to GRK3 and the expression of β-arrestin, which is recruited by the phosphorylated β2-AR for internalization and degradation, increased in aged rat liver. GRK3 overexpression also decreased the glucose output from young rat hepatocytes. We conclude that an age-associated reduction in epinephrine-induced gluconeogenesis occurs through the epinephrine receptor desensitizing system.
Similar content being viewed by others

Introduction
Since glucose is the main energy source for cells, maintaining blood glucose homeostasis is essential for life. Insulin down-regulates blood glucose levels by enhancing glucose uptake into muscle and adipocytes, as well as by reducing liver gluconeogenesis. When blood glucose levels fall below a certain threshold or during times of extreme physical stress, glucose levels are restored by de novo synthesis from other sources and glycogen degradation. Gluconeogenesis is enhanced by hormones, such as glucagon, epinephrine, and glucocorticoid (Short et al., 1986, Wynshaw-Boris et al., 1986). When blood glucose levels fall below the normal range, glucagon is secreted from the islets of Langerhans in the pancreas. Epinephrine secreted from the adrenal medulla prepares the body to meet an emergency. Circulating glucagon and epinephrine activate cellular signals that catalyze components of adenylate cyclase via the stimulatory guanine nucleotide binding regulatory protein Gαs (Jelinek et al., 1993, Erraji-Benchekroun et al., 2005). This signal increases formation of cAMP and other transcription factors. Phosphoenolpyruvate carboxykinase (PEPCK), the key regulatory enzyme for gluconeogenesis (Valera et al., 1994; Burgess et al., 2004) responds to signals from glucagon and epinephrine (Hanson and Reshef, 1997), which activate PKA via cAMP leading to phosphorylation of the transcription factor cAMP response element binding (CREB). The result is activation of phospho-CREB binding to the PEPCK promoter (Liu et al., 1991, Pilkis et al., 1975). Adenylate cyclase cascade activated CREB phosphorylation synergistically induces strong transcription of peroxisome proliferator-activated receptor gamma cofactor 1 α (PGC-1α), the cofactor for PEPCK (Herzig et al., 2001, Yoon et al., 2001).
There is disagreement regarding the gluconeogenesis response during aging. The catecholamine challenge in aged individuals shows deteriorated response (Morgan et al., 1983; Mabry et al., 1995; Podolin et al., 1996; Marker et al., 1998; Podolin et al., 2001). A deterioration of the catecholamine response with aging was suggested to be due to an age related decrease in the β2 adrenergic receptor (AR), adenylate cyclase, or to Gαs expression (Dax et al., 1981 Fraeyman et al., 2000). In contrast, it has been suggested that expressions of the β2-AR, adenylate cyclase or Gαs either increase or do not change with aging (Dax, 1987; Eakes, 1996; Marker, 1998; Sumida, 2003; Jang, 2006). Thus, the questions of why and how epinephrine becomes less potent in the gluconeogenesis response in aging remain unanswered. We compared the effects of glucagon and epinephrine on gluconeogenesis in young and old rat hepatocytes to investigate the underlying mechanism of the diminished gluconeogenesis response of epinephrine in aging.
Results
Senescent rat hepatocytes respond differently to glucagon and epinephrine
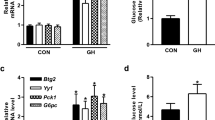
How gluconeogenesis is influenced by different stimuli during aging probably reflects an aged individual's ability to adapt to senescence because glucose is the main energy source for cells. Since gluconeogenesis is enhanced by hormones, such as glucagon, and epinephrine, we compared the effects of these hormones on gluconeogenesis in young and old rat primary hepatocytes. Glucagon is secreted from the islets of Langerhans alpha cells during hypoglycemic conditions, such as fasting, while epinephrine is mainly secreted from the adrenal glands under conditions of physical stress. Glucagon and epinephrine share cAMP as their secondary messenger, although the conditions of their secretion are different. Both glucagon and epinephrine activate PEPCK, the key regulatory enzyme in gluconeogenesis (Figure 1A and C) and its regulatory cofactor PGC-1α (Figure 1B and D) in young rat primary hepatocytes. Glucagon activates PEPCK and its cofactor PGC-1α in old (Figure 1C and D) and young hepatocytes. In contrast, epinephrine induced PEPCK and PGC-1α in old rat hepatocytes to a lesser extent than in young rat hepatocytes (Figure 1 A and B). As shown in Figure 1, PEPCK expression was marginally enhanced by epinephrine in a relatively short period. These relative effects of glucagon and epinephrine are reflected in the glucose output in both young and old rat hepatocytes. Glucagon significantly increased the glucose output in both young and old rat hepatocytes, while epinephrine increased the secreted glucose level only slightly in old rat hepatocytes (Figure 2).

Epinephrine induced PEPCK and PGC-1α expressions differ in young and old rat primary hepatocytes. (A) PEPCK mRNA expression due to 10 µM epinephrine in young and old rat primary hepatocytes. (B) PGC-1α mRNA expression due to 10 µM epinephrine in young and old rat primary hepatocytes. (C) PEPCK mRNA expression due to 10 nM glucagon in young and old rat primary hepatocytes. (D) PGC-1α mRNA expression due to 10 nM glucagon in young and old rat primary hepatocytes. PEPCK and PGC-1α mRNA expression was measured using real time PCR with β-actin mRNA expression as an internal control. Data are graphed as mean ± S.E. from triplicate experiments. An open square represents values from young (4 month) rat hepatocytes and a black square represents values from old (24 month) rat hepatocytes.

Glucose formation in response to epinephrine in young and old rat primary hepatocytes is different, unlike the response to glucogan. Glucose formation measured after 10 nM glucagon and 10 µM epinephrine treatments in young and old rat primary hepatocytes after a 6 h incubation with glucose formation buffer using the enzymatic method. Values were normalized with the total protein concentration and are shown as a percentage of the control after glucagon and epinephrine treatments. Data represent mean ± S.E. from three separate duplicated experiments. An open square represents values from young (4 month) rat hepatocytes and a black square represents values from old (24 month) rat hepatocytes. An asterisk *indicates a statistically significant difference (P < 0.05) based on a two-tailed Student's t-test.
Epinephrine-induced CREB phosphorylation is different in old and young rat hepatocytes
Even though glucagon and epinephrine share the same intracellular messenger cAMP, glucagon and epinephrine-induced gluconeogenesis responses in senescent cells are quite different. We compared intracellular signals in young and old rat hepatocytes in response to glucagon and epinephrine. As shown in Figure 3, glucagon activates signals for CREB phosphorylation in young rat hepatocytes quite early and continues for 6 h. Old rat hepatocytes maintain glucagon-induced CREB phosphorylation almost to the same extent as do young hepatocytes, but not epinephrine-induced CREB phosphorylation. Consistent with the diminished transcriptional activation of PEPCK and PGC-1α, old rat hepatocytes maintained the phospho-CREB level for only 5 min following epinephrine treatment. We compared CREB phosphorylation in young and old hepatocytes after treatment with a nondegradable form of cAMP. The degree of Bt-cAMP-induced CREB phosphorylation in old hepatocytes was not different from the degree in young hepatocytes (Figure 3). Insulin treatments were used as a negative control for CREB phosphrylation. These results indicate that the observed age related differences in epinephrine-induced gluconeogenesis may not be related to PEPCK or PGC-1α activation at the transcriptional level, or to CREB phosphorylation.

Activation of CREB phosphorylation by epinephrine in old rat hepatocytes is short-term compared to young hepatocytes. Western blot analysis of CREB and its phosphorylation after indicated drug treatments. CREB phosphorylation stimulated with 10 nM glucagon, 10 µM epinephrine, 1 mM dibutyric cAMP, and 1 µg/µl of insulin in young and old rat primary hepatocytes.
β2-AR signaling in aged rat hepatocytes
The decreased gluconeogenesis response to epinephrine in aging could originate prior to CREB phosphorylation since the phosphoration of CREB and the subsequent activation of PGC-1α and PEPCK occur in parallel. In addition, Bt-cAMP activation of CREB phosphorylation was not different in young and old rat hepatocytes. Thus, the epinephrine receptor activation process might be different in young and old hepatocytes, so we compared epinephrine receptor expression and its desensitization in young and old cells. As previously reported, β2-AR expression decreased slightly in old liver cells and the patterns of the GRK2/3 kinases involved in the activation of the β2-AR signal desensitizing (Goodman et al., 1996; Pierce et al., 2002) were different in young and old liver cells. As shown in Figure 4, GRK2, the major isoform in young rat liver, changes to GRK3 in aged rat isolated liver cells. In addition, the level of the desensitizing internalization point regulator β-arrestin (Goodman et al., 1996) also increased in old rat liver cells. These data support our hypothesis that epinephrine desensitizes the epinephrine receptor faster in old cells.

GRK2 expression changes to GRK3 and β-arrestin expression increases in aging. Western blot analysis of β2-AR internalization related GRK2/3 and β-arrestin in young and old rat liver cells.
The role of GRK in epinephrine-induced gluconeogenesis
β2-AR desensitization starts with kinase activation (Pierce et al., 2002). The major isoform of GRK involved in the β2-AR changes from GRK2 to GRK3 during the aging process and this probably influences glucose formation. When GRK3 was overexpressed in young hepatocytes, the glucose output decreased regardless of whether the cells were treated with epinephrine or not (Figure 5). However, GRK2 overexpression did not decrease the glucose output (data not shown).

Grk3 overexpression decreases the glucose output in young rat primary hepatocytes. The glucose output after Grk3 overexpression significantly decreased regardless of 10 µM epinephrine treatment. Data are represented as mean ± S.E from duplicated experiments. A glucose formation assay was performed after a 6 h incubation with glucose formation buffer and normalized to the total cell protein content. Western blot analysis showed GRK3 overexpressed in young rat hepatocytes.

A model for gluconeogenesis based on glucagon and epinephrine treatments in young and old rat primary hepatocytes.
Discussion
We studied the effects of glucagon and epinephrine on the de novo glucose synthetic pathway in aged cells. These two hormones share the same internal signaling pathway, but are synthesized under distinctly different physiological conditions. Glucagon increased the output of glucose, PEPCK, and its cofactor PGC-1α with similar activation patterns in both young and old hepatocytes. In addition, the intermediate signaling cascades for CREB phosphorylation appear to be almost the same in both groups. However, there were some differences between young and old rat hepatocytes for certain stress-activated epinephrine responses. Enhancement of PEPCK and PGC-1α expression was limited and glucose output was significantly lower in old epinephrine- treated hepatocytes. Phospho-CREB, which directly enhances PEPCK and PGC-1α expression, only activated till 5 min in old hepatocytes, compared to nearly 2 h necessary for its disappearance in young hepatocytes. This does not appear to be due to any internal system default since the non-degradable form of cAMP maintains phospho-CREB in both groups. These results point to upper stream signaling molecules, such as the epinephrine receptor and the receptor desensitizing system as causes for the observed differences in old hepatocytes. As shown in Figure 3, epinephrine receptor expression was slightly decreased in old cells. However, the major isoform of β2-adrenergic receptor kinase involved in internalizing its receptor for degradation (Benovic et al., 1991, Parruti et al., 1993), changed from GRK2 to GRK3 in aged rat hepatocytes. In addition, the expression of β-arrestin, which is recruited by phosphorylated β2-AR for ubiquitination and degradation (Goodman et al., 1996, Pierce et al., 2002), increased in aged rat hepatocytes. These results indicate that a possible mechanism for the decreased epinephrine-induced gluconeogenesis in aged rat hepatocytes is fast ligand-activated β2-AR desensitization.
Age related epinephrine responses have been studied. Electric shock induced catecholamine secretion from the adrenal medulla together with decreased blood glucose responses in old rats (Mabry et al., 1995). Catecholamine-stimulated lipolysis, glycogenolysis, and β2-AR mediated vasorelaxation all declined in aged animal aortic tissues (Dax et al., 1981; Fraeyman and van Ermen, 1993; Dobson et al., 2003). To understand the reasons for the reduced response to β2-AR-stimulated intracellular function, aged rat hepatocyte viability, receptor density, receptor binding affinity, and adenylate cyclase activities were studied (Morgan et al., 1983; Fraeyman et al., 2000). The β2-AR number, the density, the Gs activity, and the protein levels were all unchanged in aging (Dax et al., 1987; Eakes et al., 1996; Marker et al., 1998; Sumida et al., 2003).
An earlier study using the livers of rats up to 18 months old showed that the β2-AR binding capacity is directly proportional to the isoproterenol stimulated adenylate cyclase activity. In livers older than 24 months the binding capacity increased, but there was no further increase in the adenylate cyclase activity. However, activation of adenylate cyclase by the nonhormonal stimulators ADP ribosylation factor and forskolin did not change with aging, indicating that the catalytic unit was not the limiting factor (Eakes et al., 1996). Since the relationship between the glucagon receptor and adenylate cyclase also remained unaltered, the uncoupling apparently lies in an alteration of β2-AR desensitization. Results of a study using aortic tissues may offer a possible explanation for the alteration of β2-AR desensitization in aging (Schutzer et al., 2001). Similar to our results, β2-AR mediated vasorelaxation in whole aortic tissue declined with advancing age, whereas the β2-AR density was maintained. β2-AR desensitization related GRK activity and expression of GRK2, GRK3, but not GRK5 and β-arrestin, increase as an organism ages (Schutzer et al., 2001). In addition, the gene expression profiles for aged myocardial tissues showed a reduction in β2-AR-induced metabolic and contractile responsiveness, and an increase in β-arrestin mRNA expression (Dobson et al., 2003). Our results also show that GRK3 and β-arrestin desensitization systems are associated with decreased epinephrine-induced gluconeogenesis. Besides, although glucagon and the β2 adrenergic receptor share the same Gs and adenylate cyclase, the glucagon receptor does not use GRKs for desensitization (Tobias et al., 1997).
We have shown here that a decreased epinephrine-induced gluconeogenesis response in old hepatocytes is associated with fast desensitization of the epinephrine activated receptor in response to GRK3 and β-arrestin. Aging associated reduction in epinephrine-induced gluconeogenesis seems to occur via the receptor desensitizing system of epinephrine and not via any internal messenger molecules that epinephrine shares with glucagon.
Methods
Hepatocyte Isolation
Four and 24 month old male Fisher 344 rats were purchased from Samtaco (Seoul, Korea). They were housed and maintained under conditions approved by the Animal Care and Use Committee of Seoul National University Medical School.
Primary hepatocytes were isolated using the two-step collagenase perfusion system described by Seglen (1976). Isolated hepatocytes (2 × 106 cells) were seeded in William's E medium (Sigma, St Louis) containing 10% FBS (Invitrogen, Carlsbad) in p60 culture dishes and left standing for 3 h, after which the medium was changed to serum free DMEM.
Hepatic glucose formation assay
Primary hepatocytes were seeded in 6 well plates (1.4 × 106 cells per well). After incubation for 4 h in William's medium with 10% FBS, the medium was changed to serum free DMEM and left overnight. The medium was replaced with 1 ml of glucose phenol red free DMEM base medium supplemented with 20 mM sodium lactate and 2 mM sodium pyruvate (Sigma). After a 4 h incubation, 0.5 ml of the medium was collected and the glucose concentration was measured using the glucose oxidase/ peroxidase method. The glucose values obtained were normalized by division using the total protein content.
PEPCK and PGC-1α mRNA expression
Total RNA was extracted from young and aged rat primary hepatocytes and liver tissue using an Ultraspec II RNA isolation system (Biotecx Laboratories, Inc., Houston, TX). One µg of total RNA was converted to cDNA via a reverse transcriptase reaction (Invitrogen). One tenth of the cDNA was used for real time PCR with a TaqMan gene expression assay system following standard methods (Applied Biosystems, Foster City, CA). PEPCK and PGC-1α expression levels were estimated as the FAM dye fluorescence intensity from PEPCK or PGC-1α primer expanded products divided by the VIC dye fluorescence intensity from β-actin primer extended products.
Transfection with the Grk3 expression vector
The Grk3 expression vector was provided by Dr. Lefkowitz (Duke University, Durham, NC). Rat hepatocytes were seeded (2 × 106 cells in p60 in each culture dish) and washed after 3 h to remove unattached cells. The following day the hepatocytes were transfected with the Grk3 expression vector using lipofectamin 2000 (Invitrogen) following the manufacturer's protocol.
Western blot analysis
Rat hepatocytes were seeded (2 × 106 cells in p60 in each culture dish) in William's medium with 10% FBS. The medium was changed to serum free DMEM medium to remove unattached cells and left overnight. Hepatocytes were treated with epinephrine, glucagon, insulin, or dibutyric cAMP (Bt-cAMP) (Sigma) and lysed with 20 mM Tris-HCl, pH 7.5, 5 mM EDTA, 10 mM Na4P2O7, 10 mM NaF, 2 mM Na3VO4, 1% NP-40, 1 mM PMSF, and a protease inhibitor cocktail (Sigma) for 30 min Twenty to 50 µg of the cell lysate was loaded onto acrylamide gel, then the proteins were separated by electrophoresis and transferred to nitrocellulose membranes. The membranes were then blocked with 5% skim milk and blotted with anti β2-AR (Santa Cruz Biotechnology, Santa Cruz, CA) anti GRK2/3 (Upstate Biotechnology, Lake Placid, NY), anti GRK3, (Santa Cruz Biotechnology) and anti β-arrestin (ABR, Golden, CO). Protein expression detected chemiluminescent signals activated by SuperSignal West Pico Chemiluminescent Substrate (Pierce Rockford, IL) reacted with horse radish peroxidase tagged secondary antibodies (Jackson ImmunoResearch Laboratories, Inc., West Grove).
Statistics
Data are presented as mean ± S.E. Statistical analysis was done by a two-tailed Student's t-test.
Abbreviations
- AR:
-
adrenergic receptor
- CREB:
-
cAMP response element binding
- GRK:
-
G protein coupled receptor kinase
- PEPCK:
-
phosphoenolpyruvate carboxykinase
- PGC-1α:
-
peroxisome proliferator-activated receptor gamma cofactor 1 α
References
Benovic JL, Onorato JJ, Arriza JL, Stone WC, Lohse M, Jenkins NA, Gilbert DJ, Copeland NG, Caron MG, Lefkowitz RJ . Cloning, expression, and chromosomal localization of beta-adrenergic receptor kinase 2. A new member of the receptor kinase family . J Biol Chem 1991 ; 266 : 14939 - 14946
Burgess SC, Hausler N, Merritt M, Jeffrey FM, Storey C, Milde A, Koshy S, Lindner J, Magnuson MA, Malloy CR, Sherry AD . Impaired tricarboxylic acid cycle activity in mouse livers lacking cytosolic phosphoenolpyruvate carboxykinase . J Biol Chem 2004 ; 279 : 48941 - 48949
Dax EM, Partilla JS, Gregerman RI . Mechanism of the age-related decrease of epinephrine-stimulated lipolysis in isolated rat adipocytes: beta-adrenergic receptor binding, adenylate cyclase activity, and cyclic AMP accumulation . J Lipid Res 1981 ; 22 : 934 - 943
Dax EM, Partilla JS, Pineyro MA, Gregerman RI . Beta-adrenergic receptors, glucagon receptors, and their relationship to adenylate cyclase in rat liver during aging . Endocrinology 1987 ; 120 : 1534 - 1541
Dobson JG, Fray J, Leonard JL, Pratt RE . Molecular mechanisms of reduced beta-adrenergic signaling in the aged heart as revealed by genomic profiling . Physiol Genomics 2003 ; 15 : 142 - 147
Eakes AT, Hymer TK, Rosenthal MJ, Moss J, Katz MS . Alterations of adenylyl cyclase-linked G proteins in rat liver during aging . Am J Physiol Endocrinol Metab 1996 ; 270 : E126 - E132
Erraji-Benchekroun L, Couton D, Postic C, Borde I, Gaston J, Guillet JG, Andre C . Overexpression of beta2- adrenergic receptors in mouse liver alters the expression of gluconeogenic and glycolytic enzymes . Am J Physiol Endocrinol Metab 2005 ; 288 : E715 - E722
Fraeyman N, Van ermen A . Influence of aging on the betaand glucagon-receptor-mediated glycogenolysis in rat hepatocytes . Mech Ageing Dev 1993 ; 70 : 115 - 126
Fraeyman N, Van de velde E, Van ermen A, Bazan A, Vanderheyden P, Van emmelo J, Vandekerckhove J . Effect of maturation and aging on [beta]-adrenergic signal transduction in rat kidney and liver . Biochemical Pharmacology 2000 ; 60 : 1787 - 1795
Goodman OB, Krupnick JG, Santini F, Gurevich VV, Penn RB, Gagnon AW, Keen JH, Benovic JL . Beta-arrestin acts as a clathrin adaptor in endocytosis of the beta2-adrenergic receptor . Nature 1996 ; 383 : 447 - 450
Hanson RW, Reshef L . Regulation of phosphoenolpyruvate carboxykinase (GTP) gene expression . Annu Rev Biochem 1997 ; 66 : 581 - 611
Herzig S, Long F, Jhala US, Hedrick S, Quinn R, Bauer A, Rudolph D, Schutz G, Yoon C, Puigserver P, Spiegelman B, Montminy M . CREB regulates hepatic gluconeogenesis through the coactivator PGC-1 . Nature 2001 ; 413 : 179 - 183
Jelinek LJ, Lok S, Rosenberg GB, Smith RA, Grant FJ, Biggs S, Bensch PA, Kuijper JL, Sheppard PO, Sprecher CA, Et A . Expression cloning and signaling properties of the rat glucagon receptor . Science 1993 ; 259 : 1614 - 1616
Liu JS, Park EA, Gurney AL, Roesler WJ, Hanson RW . Cyclic AMP induction of phosphoenolpyruvate carboxykinase (GTP) gene transcription is mediated by multiple promoter elements . J Biol Chem 1991 ; 266 : 19095 - 19102
Mabry TR, Gold PE, Mccarty R . Age-related changes in plasma catecholamine responses to acute swim stress . Neurobiol Learn Mem 1995 ; 63 : 260 - 268
Marker JC, Clutter WE, Cryer PE . Reduced epinephrine clearance and glycemic sensitivity to epinephrine in older individuals . Am J Physiol 1998 ; 275 : E770 - E776
Morgan NG, Blackmore PF, Exton JH . Age-related changes in the control of hepatic cyclic AMP levels by alpha 1- and beta 2-adrenergic receptors in male rats . J Biol Chem 1983 ; 258 : 5103 - 5109
Parruti G, Ambrosini G, Sallese M, De blasi A . Molecular cloning, functional expression and mRNA analysis of human beta-adrenergic receptor kinase 2 . Biochem Biophys Res Commun 1993 ; 190 : 475 - 481
Pierce KL, Premont RT, Lefkowitz RJ . Seven-transmembrane receptors . Nat Rev Mol Cell Biol 2002 ; 3 : 639 - 650
Pilkis SJ, Claus TH, Johnson RA, Park CR . Hormonal control of cyclic 3':5'-AMP levels and gluconeogenesis in isolated hepatocytes from fed rats . J Biol Chem 1975 ; 250 : 6328 - 6336
Podolin DA, Gleeson TT, Mazzeo RS . Hormonal regulation of hepatic gluconeogenesis: influence of age and training . Am J Physiol 1996 ; 270 : R365 - R372
Podolin DA, Wills BK, Wood IO, Lopez M, Mazzeo RS, Roth DA . Attenuation of age-related declines in glucagon-mediated signal transduction in rat liver by exercise training . Am J Physiol Endocrinol Metab 2001 ; 281 : E516 - E523
Schutzer WE, Reed JF, Bliziotes M, Mader SL . Upregulation of G protein-linked receptor kinases with advancing age in rat aorta . Am J Physiol Regul Integr Comp Physiol 2001 ; 280 : R897 - R903
Seglen PO . Preparation of isolated rat liver cells . Methods Cell Biol 1976 ; 13 : 29 - 83
Short JM, Wynshaw-Boris A, Short HP, Hanson RW . Characterization of the phosphoenolpyruvate carboxykinase (GTP) promoter-regulatory region. II. Identification of cAMP and glucocorticoid regulatory domains . J Biol Chem 1986 ; 261 : 9721 - 9726
Sumida KD, Arimoto SM, Catanzaro MJ, Frisch F . Effect of age and endurance training on the capacity for epinephrine-stimulated gluconeogenesis in rat hepatocytes . J Appl Physiol 2003 ; 95 : 712 - 719
Tobias ES, Rozengurt E, Connell JM, Houslay MD . Cotransfection with protein kinase D confers phorbol-estermediated inhibition on glucagon-stimulated cAMP accumulation in COS cells transfected to overexpress glucagon receptors . Biochem J 1997 ; 326 : 545 - 551
Valera A, Pujol A, Pelegrin M, Bosch F . Transgenic mice overexpressing phosphoenolpyruvate carboxykinase develop non-insulin-dependent diabetes mellitus . Proc Natl Acad Sci USA 1994 ; 91 : 9151 - 9154
Wynshaw-Boris A, Short J, Loose D, Hanson R . Characterization of the phosphoenolpyruvate carboxykinase (GTP) promoter-regulatory region. I. Multiple hormone regulatory elements and the effects of enhancers . J Biol Chem 1986 ; 261 : 9714 - 9720
Yoon JC, Puigserver P, Chen G, Donovan J, Wu Z, Rhee J, Adelmant G, Stafford J, Kahn CR, Granner DK, Newgard CB, Spiegelman BM . Control of hepatic gluconeogenesis through the transcriptional coactivator PGC-1 . Nature 2001 ; 413 : 131 - 138
Acknowledgements
The authors thank Dr. R.J. Lefkowitz (Duke University, Durham, NC) for the GRK3 vector and Dr. Yong-Sung Juhnn (Seoul National University, Seoul, Korea) for critical discussion. This work was supported by the Korea Research Foundation of Health Science (R11-2002-097-05001-0), Korea Science and Engineering Foundation (KOSEF, R11-2002-097-00001-0) for Aging and Apoptosis Research Center at Seoul National University, and The Korea National Cancer Center (0710280-1).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Kim, K., Cho, S., Cova, A. et al. Alterations of epinephrine-induced gluconeogenesis in aging. Exp Mol Med 41, 334–340 (2009). https://doi.org/10.3858/emm.2009.41.5.037
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3858/emm.2009.41.5.037
