
Abstract
The T-cell-mediated immune response is implicated in many clinical hepatic injuries, such as autoimmune hepatitis and acute virus hepatitis. CD24 is widely expressed by different immune cells and plays an important role in the pathogenesis of many autoimmune diseases. However, the role of CD24 in T-cell-mediated liver injury has not been elucidated until now. Here we showed that CD24 deficiency protects mice from concanavalin A (ConA)-induced fulminant liver injury by reducing serum interferon-γ (IFN-γ) levels. CD24 expression by hepatic T cells was markedly increased following ConA challenge. Moreover, decreased IFN-γ production by hepatic CD4+ T cells in CD24-deficient mice was detected, which was correlated with downregulated phosphorylation of STAT1 in hepatic tissue. In vitro experiments also supported the conclusion that CD24 deficiency impaired IFN-γ production by CD4+ T cells following ConA, CD3/CD28 and phorbol myristate acetate/ionomycin stimulation. Our study suggests that CD24 deficiency confers hepatoprotection by decreasing CD4+ T-cell-dependent IFN-γ production in vivo, which suggests that CD24 might be a potential target molecule for reducing clinical hepatitis.
Similar content being viewed by others
INTRODUCTION
Autoimmune hepatitis (AIH) and acute viral hepatitis (VH) are characterized by acute liver injury with a high mortality.1 The mechanisms of acute liver injury, other than those directly related to clinical treatments, are not completely understood, which limits the efficacy of clinical treatments of these diseases. It is well known that AIH and VH are induced by different environmental agents, such as drugs and viruses, which result in the response of CD4+ T cells against liver cells.2, 3, 4, 5, 6, 7 Some cytokines, such as IFN-γ and tumor necrosis factor (TNF), are involved in the pathogenesis of liver injuries, and injury to hepatic tissues or hepatocellular death could occur within a short period of time.8, 9, 10, 11, 12, 13 Increased evidence has demonstrated that the liver maintains a special local immune-tolerogenic microenvironment owing to its unique blood supply.14, 15 It is unclear why immune tolerance and immune responses that result in immune damage to the liver occur in some individuals. As concanavalin A (ConA) stimulation can mimic patterns of fulminant immunological liver injury in vivo,16, 17 the functions of hepatic immune cells were analyzed after the injection of ConA in the present study.
CD24 was first discovered in 1978 as a heat-stable antigen, and it was successfully cloned in 1990.18 CD24 is expressed on non-T lymphocytes, such as dendritic cells and B lymphocytes, and regulates their homeostasis and proliferation. Recently, it was found that CD24 can enhance autoimmunity diseases due to its protective effect in the clonal deletion of autoreactive thymocytes, and according to our observations, the expression of CD24 on hepatic T cells markedly increases after ConA stimulation.
Specific receptors on the cell membranes linked with T-cell-mediated hepatic cell injuries are still being discovered.1 Researchers have found that scavenger receptor A (SRA/CD204)19 and CD49a20 mediate fulminant hepatic failure. With highly variable glycosylation sites, the heat-stable antigen (mouse CD24) binds to different ligands, including SiglecG (mouse), Signlec10 (human21), P-selectin22 and danger-associated molecular patterns (DAMPs).23 These ligands provide various routes by which CD24 can participate in the pathogenesis of human24 autoimmune diseases, such as multiple sclerosis25 and ankylosing spondylitis.26 Some reports have shown that CD24 polymorphism is a risk factor for organ-specific and systemic autoimmune diseases.27, 28 Recently, scientists have found that targeted mutations of CD24 induce resistance to autoimmune encephalomyelitis in mice.29, 30
The functional roles of CD24 expressed by hepatic T cells following the injection of ConA were investigated, and CD24 on activated T lymphocytes could aggravate acute liver injury in AIH by promoting the production of interferon-γ (IFN-γ) by CD4+ T cells. Thus, CD24 could be a novel therapeutic target for the treatment of AIH or AH.
MATERIALS AND METHODS
Mice
All animal experiments were performed in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals, with the approval of the Scientific Investigation Board of the Second Military Medical University. CD24 knockout mice were a gift from Professor Yang Liu. They were bred in specific pathogen-free conditions, and 6–8 weeks old littermates were used in the experiments (body weight and sex balanced). CD45.1+ wild-type mice were acquired from The Jackson Laboratory (Bar Harbor, ME, USA, Stock No.: 002014).
Reagents
Anti-mouse CD4- and CD8-coated magnetic beads were obtained from Miltenyi Biotech (Bergisch Gladbach, Germany). C57BL/6 mice were purchased from Joint Ventures Sipper BK Experimental Animal Co. (Shanghai, China) RPMI medium 1640 and FBS were purchased from PAA Laboratories (Freiburg, Germany). PE-CY7-conjugated anti-mouse CD3, APC-conjugated anti-mouse CD8, FITC-conjugated anti-mouse CD4 and PE-conjugated anti-mouse IFN-γ antibodies were purchased from eBioscience. PE-conjugated anti-mouse CD24 antibodies were purchased from Acris Antibodies (Rockville, MD, USA). Antibodies specific to p-ERK (Thr202/Tyr204), p-JNK (Thr183/Tyr185), p-p38 (Thr180/Tyr182), p-p65 (Ser536), p-PLCγ1 (Tyr783), p-Zap70 (Tyr493), p-LCK (Tyr505), LCK and β-actin were obtained from Cell Signaling Technology (Danvers, MA, USA). Trizol reagents were from Invitrogen (Carlsbad, CA, USA). Prime Script RT-PCR Kit and SYBR Premix ExTaq kit were from Takara Bio. Concanavalin A (ConA) was obtained from Sigma-Aldrich (Saint Louis, MO, USA), and α-galactosylceramide (α-GalCer) was obtained from Enzo Life Sciences (Farmingdale, NY, USA). A monoclonal antibody specific to CD4 (GK1.5) was obtained from BioXCell (West Lebanon, NH, USA).
Cell preparations
T cells were enriched from spleen mononuclear cells via positive selection with anti-mouse CD4 or CD8 magnetic beads, the purity of which was confirmed to be over 92% by FACS. Splenocytes or isolated T cells were stimulated with ConA (5-10 μg/ml) for different times as indicated.
ELISA and cytometric bead array immunoassay
IL-6, IFN-γ, IL-10, IL-12p40, TNF and MCP-1 concentrations were measured by cytometric bead array (CBA) immunoassays according to the manufacturer’s protocols (BD Biosciences, San Jose, CA, USA). IL-4 concentration was measured by ELISA manufacturer’s protocols (R&D, Minneapolis, MN, USA).
ConA-induced liver injury model
Mice were intravenously injected with ConA (10 or 20 mg/kg body weight). Liver histology and survival were evaluated to represent disease severity. Serum alanine aminotransferase (ALT) was detected according to the manufacturer’s protocols (Rongsheng Company, Shanghai, China).
Evaluation of α-GalCer-induced fulminant hepatitis model
α-GalCer (100 g/kg) was dissolved in 500 μl saline. Serum ALT levels were detected by α-GalCer injections at different time points as indicated. Histological analysis of hepatic injury was shown with H&E staining.
Adoptive re-transfer experiment
Wild-type (WT) mice (CD45.1+) were injected with a monoclonal antibody against CD4+ T cells (GK1.5, 40 μg/mouse), and the degree of CD4 depletion before the reconstitution experiment was calculated by FACS. CD4+ T cells (6 × 106/mouse) were separated from the spleens of untreated CD24−/− mice or their WT littermates with magnetic beads and then intravenously injected into the mice immediately before ConA injection. The samples were collected for H&E staining, ALT measurements and serum IFN-γ assays.
Flow cytometry and IFN-γ intracellular analysis
For intracellular IFN-γ staining, cells were stimulated with ConA (10 μg/ml), plate-bound anti-CD3 (10 μg/ml) plus anti-CD28 (1 μg/ml) or phorbol myristate acetate (PMA; 25 ng/ml) plus ionomycin (500 ng/ml) for different times at 37 °C. GlogiStop was added for the next 4 h of incubation. The cells were stained with monoclonal antibodies for 30 min at room temperature to evaluate the cell surface expression levels of CD24, CD4 and CD8, then fixed using a Fixation/Permeabilization Solution Kit, and finally stained with anti-mouse IFN-γ antibody. Flow cytometric analysis was performed using an LSR II with FACS Diva software (BD Biosciences).
CD4+ T-cell activation and proliferation
Purified CD4+ T cells were cultured in RPMI 1640 plus 10% FCS. The CD4+ T cells (2 × 105 per well) were activated with ConA (10 μg/ml), plate-bound anti-CD3 mAb (10 μg/ml) plus anti-CD28 mAb (1 μg/ml) or PMA (25 ng/ml) plus ionomycin (500 ng/ml). After the indicated times, the CD69 MFI and IFN-γ expression levels were analyzed by flow cytometry.
Isolation of liver lymphocytes and Kupffer cells
Mouse livers were cut into pieces and incubated in collagenase-containing buffer (Type IV, 0.05%) for 30 min followed by pressing with a 200-gauge steel mesh. Then, the cells were collected, suspended with saline, and centrifuged at 1000g for 5 min. The cell pellets were collected and resuspended in 40% Percoll (GE Healthcare, Freiburg, Germany), overlaid gently with 20% Percoll on the 40% Percoll, and then centrifuged for 17 min at 2800g. Liver mononuclear cells were taken from the interphase and centrifuged at 1000g for 5 min before collected as liver mononuclear cells. Hepatic Kupffer cells were isolated as previously described.31
Quantitative real-time PCR
Total RNA was extracted using Trizol Reagent (Invitrogen) followed by cDNA synthesis using Reverse Transcriptase M-MLV (Takara, Dalian, China). Subsequently, cDNA was used to measure the mRNA levels of TNF, IFN-γ, IL-2, IL-4, IL-6, IL-10 and IL-12p40 using a Real-Time PCR System (Roche, Basel, Switzerland). The relative quantifications were measured by the comparative CT method. The primer sequences used were as follows:
IFN-γ
forward: CACAGTCATTGAAAGCCTAGA,
reverse: TTGCCAGTTCCTCCAGATAT;
IL-4
forward: CTTGGGACTGATGCTGGTGACAA,
reverse: TCATTTCCACGATTTCCCAGAGAA;
IL-6
forward: CTTGGGACTGATGCTGGTGACAA,
reverse: TCATTTCCACGATTTCCCAGAGAA;
IL-10
forward: CTTGGGACTGATGCTGGTGACAA,
reverse: TCATTTCCACGATTTCCCAGAGAA;
IL-12p40
forward: GGCTGGTGCAAAGAAACATGGACTTGA,
reverse: TGCAGACAGAGACGCCATTCCACAT;
TNF
forward: CACAGTCATTGAAAGCCTAGA,
reverse: TTGCCAGTTCCTCCAGATAT;
β-actin
forward: AGTGTGACGTTGACATCCGT
reverse: GCAGCTCAGTAACAGTCCGC
Western blot analysis
To evaluate the different expression levels of proteins, cells were washed with cold PBS and resuspended in lysis buffer on ice for half an hour. The lysed cells were centrifuged at 12 000g for 5 min at 4 °C, and the supernatant was collected. The proteins were separated through 10% SDS–polyacrylamide gel electrophoresis and transferred onto nitrocellulose membranes. After blocking for 2 h at room temperature in 5% milk, the blots were incubated with the indicated monoclonal antibody overnight at 4 °C. Secondary antibodies were incubated at room temperature for 1.5 h.
Statistics
All statistical analyses were performed with SPSS 17 statistical software for Windows (SPSS, Armonk, NY, USA). The survival curves were assessed by the Kaplan–Meier method, and Statistical significance between two groups was evaluated using unpaired Student’s t-tests. P-values <0.05 were considered significant.
Results
CD24 deficiency reduces ConA-induced liver injury in mice
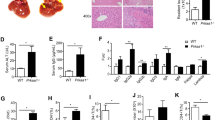
To verify the function of CD24 in acute hepatic injury, the survival times of WT and CD24−/− mice in a model of ConA-induced liver injury was investigated. The WT and CD24−/− mice were intravenously injected with a high dose of ConA (20 μg/g). The WT mice began to die 3 h after injection. All mice had died after 24 h. The survival rate of the CD24−/− mice was 50% at 18 h after injection and did not change after extension to 36 h. On the basis of the survival curve, it was confirmed that CD24 deficiency greatly reduced the mouse death rate due to ConA-induced fulminant liver injury (Figure 1a). Then, ConA was adjusted to a medium dose (10 μg/g), and serum ALT was tested at various time points post injection. In the WT mice, the level of ALT increased significantly at 6 h post injection and reached the highest level at 12 h post injection (Figure 1b). In the CD24−/− mice, serum ALT was maintained at physiological levels before the injection of ConA. At 6 and 12 h post injection, the serum ALT levels in the CD24−/− mice were lower than those in the WT mice (Figure 1b). Before and after the injection of ConA, liver specimens were obtained and stained with H&E. Before the injection of ConA, there was no pathological injury in the CD24−/− or WT mice. At 12 h after injection, marked liver hyperemia, lymphocytic infiltration, swelling, degeneration, rupture and necrosis of liver cells were observed in the WT mice (Figure 1c). However, there was less liver hyperemia, lymphocytic infiltration and swelling of liver cells and no clear rupture or necrosis of liver cells in the CD24−/− mice after injection (Figure 1c).

CD24 deficiency protects mice from ConA-induced liver injury. (a) Survival was followed for 36 h after ConA (20 μg/g body weight) injection. (b) Serum levels of ALT in CD24-deficient mice and their WT littermates after intravenous injection of ConA (10 μg/g body weight). (c) Liver sections from CD24−/− mice and their WT littermates were stained with H&E 12 h post ConA injection. Saline-treated mice were used as a control group. The percentages of the necrotic area were calculated with Image J. Values are presented as the mean±s.d. *P⩽0.05; **P⩽0.01. Similar results were obtained for three independent experiments. ALT, alanine aminotransferase; ConA, concanavalin A; WT, wild type.
Cytokines play a key role in ConA-mediated liver injury
At three time points (0, 6, and 24 h) after ConA injection, the levels of serum cytokines in the CD24−/− mice and WT littermates were detected. The serum levels of IFN-γ (Figure 2Aa) and TNF (Figure 2Ab) were markedly lower in the CD24−/− mice than in the WT mice. However, the differences in IL-4 (Figure 2Ac), IL-6 (Figure 2Ad), IL-10 (Figure 2Ae) and IL-12p40 (Figure 2Af) between the CD24−/− mice and their WT littermates were not statistically different.

Reduced inflammatory cytokines in CD24-deficient mice. (A) Mice were injected with ConA (10 μg/g) and killed. Serum levels of IFN-γ (a), TNF (b), IL-4 (c), IL-6 (d), IL-10 (e) and IL-12p40 (f) in WT (black bars) and CD24−/− mice (white bars) were measured and analyzed via ELISA or CBA at the indicated time points (6 and 24 h, n=5). ND, not detected. (B) The levels of IFN-γ (a), TNF (b), IL-4 (c), IL-6 (d), IL-10 (e) and IL-12p40 (f) in WT (black bars) and CD24−/− mice (white bars) following ConA (10 μg/g) injection (n=5) were measured and analyzed via real-time quantitative PCR. Values are presented as the mean±s.d. *P⩽0.05; **P⩽0.01. The data are representative of at least three independent experiments with similar results, and triplicate samples were used for the statistical analysis. CBA, cytometric bead array; ConA, concanavalin A; ELISA, enzyme-linked immunosorbent assay; NS, not significant; TNF, tumor necrosis factor; WT, wild type.
The hepatic expression levels of the Ifng (IFN-γ), Tnf (TNF), Il4 (IL-4), Il6 (IL-6), Il10 (IL-10) and Il12b (IL-12p40) genes were also evaluated by real-time quantitative PCR. Compared with the saline-injected group of WT littermates, at 6 h post injection, the mRNA levels of IFN-γ (Figure 2Ba), TNF (Figure 2Bb), IL-4 (Figure 2Bc), IL-6 (Figure 2Bd), IL-10 (Figure 2Be) and IL-12p40 (Figure 2Bf) were increased in the ConA-injected group of WT littermates. After 24 h, the levels of cytokines decreased substantially. In a comparison between the CD24−/− mice and their WT littermates, IFN-γ mRNA (Figure 2Ba) and TNF mRNA (Figure 2Bb) in the livers of the CD24−/− mice were 44% and 20% lower, respectively, and no marked differences were found in the mRNA levels of IL-4 (Figure 2Bc), IL-6 (Figure 2Bd), IL-10 (Figure 2Be) or IL-12p40 (Figure 2Bf) between the two groups of mice. These data suggested that CD24 signaling aggravates ConA-induced liver injury via increased pro-inflammatory factors.
CD24 deficiency is dispensable for the recruitment of mononuclear lymphocytes in the liver after ConA injection
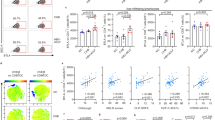
Investigations of ConA-induced liver injury have mainly focused on mononuclear lymphocytes (including T and NKT cells) raised in the liver.32, 33 To determine the main subsets that contributed in our model, T cells (CD3+), B cells (CD19+), natural killer cells (NK1.1+), Kupffer cells (F4/80+ & CD11b+) and neutrophils (LY6G+ and CD11b+) in the liver were detected in vivo after ConA injection. No marked differences in the proportions of NK cells, T cells and NKT cells (Figure 3a) were found between the CD24−/− mice and their WT littermates after ConA injection. In addition, no differences in the proportions of Kupffer cells, neutrophils, monocytes and B cells (Supplementary Figure 1) were found. Some studies have reported that CD4+ T cells mediate the process of ConA-induced liver injury.34 Therefore, the proportions of CD4+ (Figure 3b) and CD8+ T (Figure 3c) cells and their activation based on the expression of CD69 were evaluated. However, these results confirmed that no marked differences in the proportions of cells with positive expression or the activation marker between the CD24−/− mice and their WT littermates were found.

CD24 deficiency does not affect T-cell activation following ConA injection. (a) CD24 deficiency does not affect T, NK and NKT cell recruitment following ConA injection. Mononuclear cells were stained with CD3 and NK1.1. The right bar chart shows the absolute number of NK, NKT and T cells. (b) CD24 deficiency does not affect CD4 activation. The right bar chart shows the percentage of CD69 in liver CD4+ T cells from the WT and CD24−/− mice at different times. (c) CD24 deficiency does not affect CD8 activation. The right bar chart shows the percentage of CD69 in liver CD8+ T cells from the WT and CD24−/− mice. Values are presented as the mean±s.d. The data are representative of three independent experiments. ConA, concanavalin A; NS, not significant; WT, wild type.
NKT cells make a critical difference on effectiveness of immune responses to liver injury.35 To evaluate the effect of the CD24 molecule on NKT cells, we established a liver injury model induced by α-GalCer (an activator of NKT cells). Histological results (Supplementary Figure 2A) and ALT levels (Supplementary Figure 2B) were compared between the CD24−/− mice and their WT littermates. There were no significant differences between the groups of mice, indicating that NKT cell activation and function were not influenced by CD24 deficiency. Therefore, it was concluded that the reduced liver injury in the CD24-deficient mice was not a result of differences in the numbers of CD4+ and CD8+ T cells in the liver or in the activation of NKT cells.
Expression of CD24 on T cells in the liver increased after stimulation with ConA
It was confirmed that CD4+ T cells play a key role in ConA-induced liver injury.8, 36 Thus, the expression levels of CD24 on CD4+ and CD8+ T cells from the liver and spleen were investigated 2 h after ConA injection. Before stimulation with ConA, the expression of CD24 by CD4+ T cells was low, but after the injection of ConA, the expression of CD24 increased significantly (Figure 4a). In addition, the expression of CD24 on CD8+ T cells from the liver and spleen (Figure 4b) and the absolute amount of CD24+CD4+ and CD24+CD8+ cells from the liver (Figure 4c) increased markedly. These data demonstrated that CD24 expression by T cells was upregulated after ConA injection.

The expression of CD24 in T cells following ConA challenge. (a) CD24 expression in CD4+ T cells of the spleen and liver treated with saline or ConA for 2 h. (b) CD24 expression in CD8+ T cells of the spleen and liver treated with saline or ConA for 2 h. (c) Absolute number of CD24+CD4+ and CD24+CD8+ cells in the liver after saline (white bar) or ConA treatment for 2 h; n=8 per group (mean±s.d, P⩽0.05). The data are representative of at least three independent experiments with similar results. ConA, concanavalin A.
CD24 deficiency specifically decreased the secretion of IFN-γ by CD4+ T cells
IFN-γ is a main factor in ConA-induced liver injury.36 Therefore, the levels of IFN-γ secreted by CD4+ and CD8+ T cells as well as by NKT cells from the livers of the CD24−/− mice and their WT littermates were detected. The expression of IFN-γ secreted by CD4+ T cells in the CD24−/− mice was lower than that of their WT littermates at 24 h after ConA injection. Unlike CD4+ T cells, no differences were found in CD8+ T cells and NKT cells (Figure 5a). CD69 expression was also investigated, and the results revealed that there was no difference in CD69 expression between the CD4+ and CD8+ T cells originating from the spleens of the two types of mice (Supplementary Figure 4). A previous study suggested that TNF derived from macrophages and T cells also contributes to the progression of ConA-induced liver injury.37 We further examined TNF expression in CD4+ T cells and Kupffer cells after ConA injection in vivo (Supplementary Figures 3A and B). The TNF mRNA levels were reduced in both CD4+ T cells and Kupffer cells in the CD24−/− mice.

Decreased production of IFN-γ from CD24-deficient CD4+ T cells. (a) Liver lymphocytes from the WT and CD24−/− mice were isolated after ConA injection. Intracellular staining of IFN-γ in liver CD4+ T cells, CD8+ T cells and NKT cells were performed. The right bar chart displays the IFN-γ expressions in CD4+ T cells and CD8+ T cells at 24 h. (b–d) As indicated in the Methods, 6 × 106 CD4+ T cells from the WT and CD24−/− mice were transferred into WT mice (CD45.1+) depleted of CD4+ T cells. Twelve hours after ConA injection, H&E staining of liver tissues (b) and the serum levels of ALT (c) and IFN-γ (d) were analyzed. The data are representative of two independent experiments, n=6 per group. Values are presented as the mean±s.d. of the mean. *P⩽0.05; **P⩽0.01. ConA, concanavalin A; NS, not significant; WT, wild type.
To determine whether the reduction of liver injury in the CD24−/− mice was mediated by CD24-deficient CD4+ T cells, CD4+ T cells of the CD24−/− and their WT littermates were isolated using magnetic beads, and an adoptive re-transfer experiment was performed. WT (CD45.1+) mice were injected with a monoclonal antibody against CD4+ T cells (GK1.5, 40 μg each, Supplementary Figure 2C). At 3 days post injection, the mice were treated with the isolated CD4+ T cells from the CD24−/− or WT mice (CD45.2+). The liver injury after the injection of ConA was evaluated. On the basis of liver tissue slices stained with H&E (Figure 5b) and the serum levels of ALT and IFN-γ (Figures 5c and d), the liver injury in the mice that received CD4+ T cells from the WT mice worsened compared with the mice treated with CD4+ T cells from the CD24−/− mice.
To confirm the effect of CD24 on the functions of T cells in vitro, CD4+ and CD8+ T cells from the spleens of the CD24−/− mice or their WT littermates were stimulated with ConA, CD3/CD28 or PMA/ionomycin. The level of IFN-γ secreted by the CD4+ T cells from the CD24−/− mice was visibly lower than that of their WT littermates after stimulation (Figure 6a). However, no differences were found in CD8+ T cells between the CD24−/− mice and their WT littermates (Supplementary Figure 5). In addition, the IFN-γ mRNA levels of CD4+ T cells from the CD24−/− mice and the levels of IFN-γ in the culture supernatants were relatively low (Figure 6b). These results all confirmed that, after stimulation in vitro, the levels of IFN-γ secreted by CD4+ T cells originating from the CD24−/− mice were markedly lower than the IFN-γ levels observed for CD4+ T cells originating from their WT littermates. We also detected reduced TNF production in ConA-activated CD4 T cells (Supplementary Figures 3C and D) at both the mRNA and protein levels. Thus, we hypothesized that the reduced IFN-γ and TNF productions may be the result of reduced activation of CD4+ T cells. Thus, we analyzed the effect of CD24 loss on TCR signaling. After TCR ligation, the downstream signaling molecules Zap70, LCK and PLCγ1 were accessed with respect to their activation. CD24 deficiency weakened the phosphorylation of PLCγ1, Zap-70 and LCK (Figure 6c).

CD24-deficient CD4+ T cells have impaired IFN-γ production in vitro. (a) Splenic CD4+ T cells from the WT and CD24−/− mice stimulated with ConA, plate-bound anti-CD3 plus anti-CD28 or PMA plus ionomycin for 4 h. The percentage of intracellular IFN-γ in the CD4+ T cells was analyzed. (b) Splenic CD4+ T cells were stimulated with plate-bound anti-CD3 plus anti-CD28. The left figure shows the IFN-γ mRNA levels of CD4+ T cells assessed via real-time quantitative PCR. The right figure shows the supernatant levels of IFN-γ assessed via CBA assays. (c) Immunoblot analysis of the total proteins and phosphorylated proteins in the whole-cell lysates of splenic WT and CD24−/− CD4+ T cells treated for various times with ConA (10 μg/ml) in vitro. Values (a, b) are presented as the mean±s.d. *P⩽0.05. The data are representative of at least three independent experiments with similar results, and triplicate samples were used for the statistical analysis. ConA, concanavalin A; WT, wild type.
Reduced activation of CD4+ T cells and phosphorylation of liver STAT1 in CD24−/− mice
Previous studies have demonstrated that STAT-related signal molecules are involved in inflammatory reactions of the liver injury.38 STAT1 dimerizes and translocates to the nucleus to induce the transcription of various genes that promote liver injury and inflammation and inhibit liver regeneration, while STAT1 in hepatocytes is predominately activated by IFN-γ. To this end, the related signal pathways of ConA-induced liver injury mediated by CD4+ T cells were investigated. Four hours after injection of ConA (10 μg/g), the total protein levels of liver STAT1 did not change, but the phosphorylation level of STAT1 was markedly reduced. Changes in the phosphorylation levels of STAT3 and p65 were not detected (Figure 7a). These results suggested that CD24 deficiency could reduce liver injury and decrease the level of IFN-γ, which could be related to decreased activation of the STAT1 signaling pathway in the liver. The phosphorylation levels of STAT1, STAT3, p65 and c-Jun in CD4+ T cells from the spleen were also evaluated. CD24 deficiency resulted in a marked decrease in the phosphorylation levels of STAT1, STAT3 and p65 in CD4+ T cells from the spleen after the injection of ConA (Figure 7b). However, no differences in the phosphorylation levels of c-Jun, ERK, JNK and p38 were detected between the spleen CD4+ T cells of the CD24−/− mice and their WT littermates (Supplementary Figure 6).

Decreased phosphorylation of STAT1 in the CD24-deficient mice following ConA stimulation. (a) Immunoblot analysis of the lysates of hepatic tissue from the WT and CD24−/− mice following ConA injection. The right bar charts displays the relative intensities of p-STAT1, p-STAT3 and p-p65 in the liver. (b) Splenic CD4+ cell lysates were analyzed via western blot (two mice were pooled together for each group). The right bar charts display the intensities of p-STAT1, p-STAT3, p-p65 and p-Jun, which were quantified after correction with STAT1, STAT3, p65 and c-Jun. β-actin served as the input control. Values are presented as the mean±s.d. The data are representative of at least three independent experiments with similar results. ConA, concanavalin A; WT, wild type.
DISCUSSION
Some studies have shown that, after adoptive transfer of CD24-deficient T cells into hosts with syngeneic lymphocytes, the proliferation of T cells is reduced, but their survival and homing function remain unchanged.39 Upon stimulation with PMA plus ionomycin, IFN-γ production by CD24-deficient CD4+ and CD8+ T cells was markedly lower than the production by normal T cells. Some researchers have found that, after stimulation with ConA or anti-CD3 plus anti-CD28 antibodies, the expression of the CD24 antigen on the surface of CD4+ T cells may briefly increase. Co-stimulation of CD3 and CD24 could promote the proliferation of CD4+ T cells.40 However, most previous studies were performed in vitro, and the role of CD24 in acute hepatic injury induced by ConA in vivo needs to be further explored.
ConA is a plant lectin globulin extracted from the seeds of Canavalia gladiata, peas, and other plants, and it has a high affinity for mannose-rich carbohydrates. As a carbohydrate ligand on the surface of T cells, it can promote the mitotic division of T cells.41 Acute hepatic injury in mice induced by the activation of T and NKT cells with ConA is currently a widely used model used to simulate clinical acute immune hepatic injury.8, 33 The role of CD24 in acute hepatic injury induced by ConA has not been reported previously and is worth studying. A better understanding of the role of CD24 in acute hepatic injury is crucial for the treatment of hepatic injury. Many specific T-cell surface receptors related to hepatic injury have been identified, such as SRA19 and CD49a,20 which can mediate the pathological process of ConA induced-acute hepatic injury.
Previous studies have fully demonstrated that CD24 plays an important role in a variety of autoimmune diseases.24, 25, 26 In a study of autogenous meningo-encephalomyelitis, it was found that CD24 could regulate the recruitment of self-reactive T cells. In normal stable situations, the expression of CD24 on peripheral T cells is very low; however, the expression of CD24 by T lymphocytes rapidly increases under stimulation of the TCR–CD3 complex.42, 43
In the study, we demonstrated that CD24 on the surface of CD4+ and CD8+ T cells was markedly increased after treatment with ConA both in vivo and in vitro. It was confirmed that the ConA induced-hepatic injury was weakened in CD24-deficient mice. On the basis of in vivo studies, we also found that the injury was not caused by changes in the number, composition ratio or activation ratio of CD4+ T cells, CD8+ T cells, NKT cells, neutrophils, monocytes, macrophages, and B cells in the liver. Further studies confirmed that CD4+ T cells, rather than CD8+ T cells or NKT cells, were involved in reducing the secretion of IFN-γ in the CD24−/− mice.
CD24 was once thought to be a co-stimulatory molecule of T cells that functioned in T-cell homeostasis and proliferation.26 Studies have found that CD24 plays a role in protecting against the clonal deletion of self-reactive thymocytes, thus mediating the occurrence and development of autoimmune diseases.44 CD24 has highly variable glycosylation sites, which can combine with multiple receptors, including DAMPs (such as high-mobility group protein B123 or heat shock protein 70 or 9045), the sialic acid-binding immunoglobulin-like lectin family G21 (SiglecG) and P-selectin.22 It has been confirmed that the interactions of CD24 with different receptors are involved in the pathogenesis of various human autoimmune diseases, such as multiple sclerosis25 and ankylosing spondylitis.26 Recent studies have found that targeted mutations in CD24 can decrease the occurrence of autoimmune cerebrospinal meningitis.29, 30 Additional studies found that an anti-CD24 antibody could block the signal transduction of inflammatory cytokines26 and affect the amplification of bone marrow-derived suppressor cells.45 Furthermore, CD24 could downregulate the activity of antigen-presenting cells to form clones of self-reactive thymocytes.44
We confirmed in vivo at the cellular level that CD24-deficient CD4+ T cells play a key role in alleviating autoimmune liver injury after treatment with ConA. α-GalCer is a ligand on NKT cells; therefore, the hepatic injury model induced by α-GalCer was considered to be an NKT cell-mediated hepatic injury model. Some studies have also confirmed that NKT cells play a key role in the ConA-induced liver injury model.34 NKT cells promote the recruitment of eosinophils in the liver by secreting IL-4 and IL-5 and cause hepatic injury.36 In the α-GalCer-induced hepatic injury model, we confirmed that CD24 does not affect the process of hepatic injury mediated by NKT cells.
Signaling molecules, including tyrosine kinase, serine/threonine protein kinase, phosphoinositide kinase, phospholipase C and guanosine triphosphatase, play important roles in the activation of T cells. Some activated transcription factors, such as NFAT and NF-κB, can initiate T cells. For example, after mitogen activated-protein kinase P38 activation, immature T cells differentiate into Th1 cells to produce IFN-γ, which functions primarily in the regulation of the serine/threonine protein kinase. CD24 regulates the recruitment of auto-reactive T cells and the activation of T cells by regulating the NF-κB,21 PTK,21 MAPK,46, 47 JAK–STAT48 and NFAT549 signaling pathways of immunocytes. CD24 inhibits the activation of NF-κB and mediates injury repair via the CD24–SigG/10 pathway.21 We also considered these signaling pathways to explore the mechanism by which CD24 deficiency mitigates ConA-induced hepatic injury.
Comparing phosphorylated protein levels between the CD24-deficient mice and the wild-type control mice in the liver before and after injection with ConA, the phosphorylation of hepatic STAT1 was significantly reduced in the CD24-deficient mice, but the phosphorylation levels of STAT3 and p65 were not affected. These results indicated that CD24 could be related to the phosphorylation of STAT1. We further investigated CD4+ T-cell-related signaling pathways and confirmed that STAT1, STAT3 and p65 were markedly downregulated, and the activities of these signaling molecules were distinctly inhibited in the CD24-deficient mice. The c-Jun signaling pathway, apoptosis related-proteins ERK, JNK and p38, and the phosphorylation levels of these molecules were not affected. These results also suggested that CD24 is related to the phosphorylation of STAT1, STAT3 and p65 for the secretion of IFN-γ by CD4+ T cells.
We also found a reduced production of TNF in CD24−/− T cells. Thus, we speculate that the reduced productions of both TNF and IFN-γ by CD4+ T cells were related to the reduced activation of TCR, as CD24 has been previously suggested to be a costimulatory molecule for T-cell activation. A pathway analysis revealed that the activation of Zap70, LCK and PLCγ-1 in CD24−/− CD4+ T cells was actually reduced after TCR ligation. As IFN-γ is the main priming agent of macrophages, reduced IFN-γ in CD24−/− mice may lead to reduced priming of macrophages and thus cause reduced TNF production by these macrophages.
Although CD24 was discovered approximately 30 years ago, there is still great research interest in this molecule. Therefore, studies of the mechanisms underlying the role of CD24 in intercellular interactions have become more popular. In fields such as tumor immunology, autoimmune diseases and so on, studies on CD24 have increased markedly. Our results provide a novel target for clinical therapies of immune-mediated hepatic injury and acute liver failure. They also provide new ideas for the future development of a CD24 therapeutic antibody and its clinical application in therapies for acute hepatic injury.
References
dos Santos DC, Neves PC, Azeredo EL, Pelajo-Machado M, Martinho JM, Pacheco-Moreira LF et al. Activated lymphocytes and high liver expression of IFN-gamma are associated with fulminant hepatic failure in patients. Liver Int 2012; 32: 147–157.
Beland K, Marceau G, Labardy A, Bourbonnais S, Alvarez F . Depletion of B cells induces remission of autoimmune hepatitis in mice through reduced antigen presentation and help to T cells. Hepatology 2015; 62: 1511–1523.
Krawitt EL . Autoimmune hepatitis. N Engl J Med 1996; 334: 897–903.
Liberal R, Vergani D, Mieli-Vergani G . Update on autoimmune hepatitis. J Clin Transl Hepatol 2015; 3: 42–52.
Manns MP, Lohse AW, Vergani D . Autoimmune hepatitis—update 2015. J Hepatol 2015; 62 (1 Suppl): S100–S111.
Robinson MW, Harmon C, O'Farrelly C . Liver immunology and its role in inflammation and homeostasis. Cell Mol Immunol 2016; 13: 267–276.
Shuai Z, Leung MW, He X, Zhang W, Yang G, Leung PS et al. Adaptive immunity in the liver. Cell Mol Immunol 2016; 13: 354–368.
Takeda K, Hayakawa Y, Van Kaer L, Matsuda H, Yagita H, Okumura K . Critical contribution of liver natural killer T cells to a murine model of hepatitis. Proc Natl Acad Sci USA 2000; 97: 5498–5503.
Gao B, Jeong WI, Tian Z . Liver: an organ with predominant innate immunity. Hepatology 2008; 47: 729–736.
Baier JL, Mattner J . Mechanisms of autoimmune liver disease. Discov Med 2014; 18: 255–263.
Mizuhara H, O'Neill E, Seki N, Ogawa T, Kusunoki C, Otsuka K et al. T cell activation-associated hepatic injury: mediation by tumor necrosis factors and protection by interleukin 6. J Exp Med 1994; 179: 1529–1537.
Iwamoto S, Kido M, Aoki N, Nishiura H, Maruoka R, Ikeda A et al. IFN-gamma is reciprocally involved in the concurrent development of organ-specific autoimmunity in the liver and stomach. Autoimmunity 2012; 45: 186–198.
Zhao Y, Zhang Y, Liu YM, Liu Y, Feng X, Liao HY et al. Identification of T cell epitopes on soluble liver antigen in Chinese patients with auto-immune hepatitis. Liver Int 2011; 31: 721–729.
Nakamoto N, Kanai T . Role of toll-like receptors in immune activation and tolerance in the liver. Front Immunol 2014; 5: 221.
Kowalewska PM, Patrick AL, Fox-Robichaud AE . Innate immunity of the liver microcirculation. Cell Tissue Res 2011; 343: 85–96.
Tiegs G, Hentschel J, Wendel AA . T cell-dependent experimental liver injury in mice inducible by concanavalin A. J Clin Invest 1992; 90: 196–203.
Heymann F, Hamesch K, Weiskirchen R, Tacke F . The concanavalin A model of acute hepatitis in mice. Lab Anim 2015; 49 (suppl 1): 12–20.
Tan Y, Zhao M, Xiang B, Chang C, Lu Q . CD24: from a hematopoietic differentiation antigen to a genetic risk factor for multiple autoimmune diseases. Clin Rev Allergy Immunol 2016; 50: 70–83.
Zuo D, Yu X, Guo C, Wang H, Qian J, Yi H et al. Scavenger receptor A restrains T-cell activation and protects against concanavalin A-induced hepatic injury. Hepatology 2013; 57: 228–238.
Chen Y, Peng H, Chen Y, Wei H, Sun R, Tian Z . CD49a promotes T-cell-mediated hepatitis by driving T helper 1 cytokine and interleukin-17 production. Immunology 2014; 141: 388–400.
Liu Y, Chen GY, Zheng P . CD24-Siglec G/10 discriminates danger- from pathogen-associated molecular patterns. Trends Immunol 2009; 30: 557–561.
Friederichs J, Zeller Y, Hafezi-Moghadam A, Grone HJ, Ley K, Altevogt P . The CD24/P-selectin binding pathway initiates lung arrest of human A125 adenocarcinoma cells. Cancer Res 2000; 60: 6714–6722.
Harris HE, Raucci A . Alarmin(g) news about danger: workshop on innate danger signals and HMGB1. EMBO Rep 2006; 7: 774–778.
Liu Y, Zheng P . CD24: a genetic checkpoint in T cell homeostasis and autoimmune diseases. Trends Immunol 2007; 28: 315–320.
Gonzalez SJ, Rojas JI, Redal MA, Patrucco L, Correale J, Argibay PF et al. CD24 as a genetic modifier of disease progression in multiple sclerosis in Argentinean patients. J Neurol Sci 2011; 307: 18–21.
Chen M, Zhang L, Ren Y, Zhang K, Yang Y, Fang Y et al. Defective function of CD24(+)CD38(+) regulatory B cells in ankylosing spondylitis. DNA Cell Biol 2016; 35: 88–95.
Zhou Q, Rammohan K, Lin S, Robinson N, Li O, Liu X et al. CD24 is a genetic modifier for risk and progression of multiple sclerosis. Proc Natl Acad Sci USA 2003; 100: 15041–15046.
Wang L, Lin S, Rammohan KW, Liu Z, Liu JQ, Liu RH et al. A dinucleotide deletion in CD24 confers protection against autoimmune diseases. PLoS Genet 2007; 3: e49.
Bai XF, Liu JQ, Liu X, Guo Y, Cox K, Wen J et al. The heat-stable antigen determines pathogenicity of self-reactive T cells in experimental autoimmune encephalomyelitis. J Clin Invest 2000; 105: 1227–1232.
Liu JQ, Carl Jr JW, Joshi PS, RayChaudhury A, Pu XA, Shi FD et al. CD24 on the resident cells of the central nervous system enhances experimental autoimmune encephalomyelitis. J Immunol 2007; 178: 6227–6235.
Froh M, Konno A, Thurman RG . Isolation of liver Kupffer cells. Curr Protoc Toxicol 2003; Chapter 14: Unit14.4.
Jung K, Kang M, Park C, Hyun Choi Y, Jeon Y, Park SH et al. Protective role of V-set and immunoglobulin domain-containing 4 expressed on kupffer cells during immune-mediated liver injury by inducing tolerance of liver T- and natural killer T-cells. Hepatology 2012; 56: 1838–1848.
Kwon HJ, Won YS, Park O, Feng D, Gao B . Opposing effects of prednisolone treatment on T/NKT cell- and hepatotoxin-mediated hepatitis in mice. Hepatology 2014; 59: 1094–1106.
Higashimoto M, Sakai Y, Takamura M, Usui S, Nasti A, Yoshida K et al. Adipose tissue derived stromal stem cell therapy in murine ConA-derived hepatitis is dependent on myeloid-lineage and CD4+ T-cell suppression. Eur J Immunol 2013; 43: 2956–2968.
Wang H, Feng D, Park O, Yin S, Gao B . Invariant NKT cell activation induces neutrophil accumulation and hepatitis: opposite regulation by IL-4 and IFN-gamma. Hepatology 2013; 58: 1474–1485.
Kaneko Y, Harada M, Kawano T, Yamashita M, Shibata Y, Gejyo F et al. Augmentation of Valpha14 NKT cell-mediated cytotoxicity by interleukin 4 in an autocrine mechanism resulting in the development of concanavalin A-induced hepatitis. J Exp Med 2000; 191: 105–114.
Grivennikov SI, Tumanov AV, Liepinsh DJ, Kruglov AA, Marakusha BI, Shakhov AN et al. Distinct and nonredundant in vivo functions of TNF produced by t cells and macrophages/neutrophils: protective and deleterious effects. Immunity 2005; 22: 93–104.
Gao B, Wang H, Lafdil F, Feng D . STAT proteins - key regulators of anti-viral responses, inflammation, and tumorigenesis in the liver. J Hepatol 2012; 57: 430–441.
Li O, Zheng P, Liu Y . CD24 expression on T cells is required for optimal T cell proliferation in lymphopenic host. J Exp Med 2004; 200: 1083–1089.
Liu Y, Jones B, Aruffo A, Sullivan KM, Linsley PS, Janeway CA Jr . Heat-stable antigen is a costimulatory molecule for CD4 T cell growth. J Exp Med 1992; 175: 437–445.
Loganathan D, Osborne SE, Glick GD, Goldstein IJ . Synthesis of high-affinity, hydrophobic monosaccharide derivatives and study of their interaction with concanavalin A, the pea, the lentil, and fava bean lectins. Arch Biochem Biophys 1992; 299: 268–274.
Bai XF, Li O, Zhou Q, Zhang H, Joshi PS, Zheng X et al. CD24 controls expansion and persistence of autoreactive T cells in the central nervous system during experimental autoimmune encephalomyelitis. J Exp Med 2004; 200: 447–458.
Wang YC, Zhu L, McHugh R, Sell KW, Selvaraj P . Expression of heat-stable antigen on tumor cells provides co-stimulation for tumor-specific T cell proliferation and cytotoxicity in mice. Eur J Immunol 1995; 25: 1163–1167.
Zhang X, Liu JQ, Shi Y, Reid HH, Boyd RL, Khattabi M et al. CD24 on thymic APCs regulates negative selection of myelin antigen-specific T lymphocytes. Eur J Immunol 2012; 42: 924–935.
Thaxton JE, Liu B, Zheng P, Liu Y, Li Z . Deletion of CD24 impairs development of heat shock protein gp96-driven autoimmune disease through expansion of myeloid-derived suppressor cells. J Immunol 2014; 192: 5679–5686.
Sammar M, Gulbins E, Hilbert K, Lang F, Altevogt P . Mouse CD24 as a signaling molecule for integrin-mediated cell binding: functional and physical association with src-kinases. Biochem Biophys Res Commun 1997; 234: 330–334.
Suzuki T, Kiyokawa N, Taguchi T, Sekino T, Katagiri YU, Fujimoto J . CD24 induces apoptosis in human B cells via the glycolipid-enriched membrane domains/rafts-mediated signaling system. J Immunol 2001; 166: 5567–5577.
Hernandez-Vargas H, Ouzounova M, Le Calvez-Kelm F, Lambert MP, McKay-Chopin S, Tavtigian SV et al. Methylome analysis reveals Jak-STAT pathway deregulation in putative breast cancer stem cells. Epigenetics 2011; 6: 428–439.
Berga-Bolanos R, Drews-Elger K, Aramburu J, Lopez-Rodriguez C . NFAT5 regulates T lymphocyte homeostasis and CD24-dependent T cell expansion under pathologic hypernatremia. J Immunol 2010; 185: 6624–6635.
Acknowledgements
This work was supported by grants from the Army Technology Research Program of China (BWS12J051), the National Natural Science Foundation of China (31570873), and the Shanghai Committee of Science and Technology (2015QA1404700). We thank Professor Guanhong Song and Prof. Xuetao Cao for their critical review of the manuscript.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information for this article can be found on the Cellular & Molecular Immunology website
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/4.0/
About this article

Cite this article
Zheng, C., Yin, S., Yang, Y. et al. CD24 aggravates acute liver injury in autoimmune hepatitis by promoting IFN-γ production by CD4+ T cells. Cell Mol Immunol 15, 260–271 (2018). https://doi.org/10.1038/cmi.2016.57
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/cmi.2016.57
Keywords
This article is cited by
-
CD73 mediates the therapeutic effects of endometrial regenerative cells in concanavalin A-induced hepatitis by regulating CD4+ T cells
Stem Cell Research & Therapy (2023)
-
Concanavalin A as a promising lectin-based anti-cancer agent: the molecular mechanisms and therapeutic potential
Cell Communication and Signaling (2022)
-
Mesenchymal stem cells alleviate experimental immune-mediated liver injury via chitinase 3-like protein 1-mediated T cell suppression
Cell Death & Disease (2021)
-
Functional heterogeneity of CD4+ T cells in liver inflammation
Seminars in Immunopathology (2021)
-
Interferon gamma inhibits the differentiation of mouse adult liver and bone marrow hematopoietic stem cells by inhibiting the activation of notch signaling
Stem Cell Research & Therapy (2019)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}