
Abstract
The cellular FLICE inhibitory protein (c-FLIP) is an endogenous inhibitor of the caspase-8 proapoptotic signaling pathway downstream of death receptors. Recent evidence indicates that the long form of c-FLIP (c-FLIPL) is required for proliferation and effector T-cell development. However, the role of c-FLIPL in triggering autoimmunity has not been carefully analyzed. We now report that c-FLIPL transgenic (Tg) mice develop splenomegaly, lymphadenopathy, multiorgan infiltration, high titers of auto-antibodies, and proliferative glomerulonephritis with immune complex deposition in a strain-dependent manner. The development of autoimmunity requires CD4+ T cells and may result from impaired thymic selection. At the molecular level, c-FLIPL overexpression inhibits the ζ chain-associated protein tyrosine kinase of 70 kDa (ZAP-70) activation, thus impairing the signaling pathway derived from ZAP-70 required for thymic selection. Therefore, we have identified c-FLIPL as a susceptibility factor under the influence of epistatic modifiers for the development of autoimmunity.
Similar content being viewed by others
Main
One of the fundamental properties of the immune system is to distinguish self from non-self, which is central to its ability to protect against pathogens and, at the same time, maintain non-responsiveness to self. This property is established during lymphocyte development within generative lymphoid organs (central tolerance) or in the secondary lymphoid organs (peripheral tolerance).1, 2 Much attention has been devoted to studies understanding the cellular and molecular basis of tolerance mechanisms, as well as the situations under which these mechanisms breakdown leading to autoimmune disease. The understanding of these complex biological processes at molecular levels has been facilitated by various transgenic (Tg) and knockout mouse strains, as well as by strains with spontaneous mutations that develop an autoimmune phenotype.
Signaling derived from the T-cell receptor (TCR) is required for thymocyte development and thymic selection. The selection of the T-cell repertoire, which is essential for establishing self/non-self discrimination of immune recognition, occurs in the thymus, and is based on the binding strength (affinity) of the TCR for peptide–MHC complexes that are presented by epithelial and bone marrow-derived antigen-presenting cells. Thymocytes that bind self-peptide–MHC complexes too strongly undergo apoptosis and fail to enter the mature T-cell pool (a process known as negative selection). The thymocytes that express low-affinity TCRs that are specific for self-peptide–MHC complexes, are the only ones to survive (in a process known as positive selection) and migrate out of the thymus.1, 2 Recent studies indicated that a number of TCR signaling molecules are involved in both positive and negative selection in the thymus.3 A defect in thymic selection results in autoimmunity.2, 3
Cellular FLICE inhibitory protein (c-FLIP) is an endogenous inhibitor of the Fas-associated death domain protein (FADD)-caspase-8 proapoptotic signaling pathway downstream of death receptors.4 The 55-kDa long form of c-FLIP (c-FLIPL) is structurally similar to caspase-8 and caspase-10, as it contains two death effector domains (DEDs) and a caspase-like domain. However, this caspase-like domain lacks residues that are important for the catalytic activity of caspase-8.5 Thus, c-FLIP can inhibit death receptor-mediated caspase-8 activation.4 Intriguingly, when expressed at a low concentration, c-FLIPL can act as an activator of pro-caspase-8, possibly by heterodimerizing with caspase-8.6 It appears that the expression levels of c-FLIPL may be crucial for its biological functions. Although mice deficient for both c-FLIPL and c-FLIPS in T cells show severe defects in thymocyte development,7, 8 c-FLIPL deficiency in T cells does not disturb thymocyte development. Rather, c-FLIPL may mediate T-cell activation and proliferation through an unidentified mechanism.7 c-FLIPL may, therefore, serve as a modulator between apoptotic and proliferative responses after TCR stimulation.9 However, it has not been shown that overexpression of c-FLIPL in the T-cell lineage triggers autoimmunity.
In this report, we describe that mice overexpressing c-FLIPL in T cells in a BALB/c background spontaneously develop an autoimmune lymphoproliferative syndrome showing features of systemic lupus erythematosus (SLE) in humans. In contrast, c-FLIPL Tg mice in the C57BL/6 (B6) background maintain tolerance to nuclear antigens, and are resistant to the development of lupus-like disease. The development of lupus-like disease in c-FLIPL Tg mice is because of altered thymic selection in the BALB/c background by interfering with the signaling pathway derived from the ζ chain-associated protein tyrosine kinase of 70 kDa (ZAP-70).
Results
c-FLIPL transgene product is specifically expressed in T cells but not in B cells and dendritic cells
c-FLIPL was expressed transgenically in the T-cell compartment as reported earlier.6 Briefly, FLAG-tagged mouse FLIPL cDNA was inserted into a target vector containing the β-globin promoter and a downstream human CD2 locus enhancer element. To confirm that the c-FLIPL transgene product is not expressed in other cell types, such as B cells and dendritic cells (DCs), c-FLIPL transgene expression in thymocytes, splenic T cells, B cells, and DCs of BALB/c and B6 backgrounds was detected using anti-FLAG Ab. As shown in Supplementary Figure 1, the c-FLIPL transgene product was detected only in thymocytes and T cells but not in B cells and DCs from BALB/c and B6 c-FLIPL Tg mice, neither in their non-Tg littermate controls (NLCs), and was comparable between BALB/c and B6 strains. Consistent with the earlier report,6 in addition to the p55 kDa form of c-FLIPL, an additional 43 kDa cleavage product of Tg c-FLIPL was found in both thymocytes and splenic T cells from BALB/c and B6 c-FLIPL Tg mice (Supplementary Figure S1).
BALB/c but not B6 c-FLIPL Tg mice spontaneously develop lupus-like disease
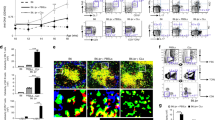
c-FLIPL Tg mice in a B6 background (B6 c-FLIPL Tg mice) do not develop any autoimmune disease.6 In accordance with these data, we did not observe any sign of autoimmunity in B6 c-FLIPL Tg mice until 10 months of age (data not shown). The absence of auto-antibodies (auto-Abs) or spontaneous autoimmune disease suggested that additional genetic or environmental factors might be necessary to trigger autoimmunity in these susceptible mice. To analyze the contribution of genetic background and modifier loci to the development of autoimmunity, we backcrossed these mice to a BALB/c background for 14 generations. Although new-born BALB/c c-FLIPL Tg mice seemed normal, these mice were smaller in size than NLCs when they grew up to 3–5 weeks of age (Figure 1a). Almost all BALB/c c-FLIPL Tg mice developed a generalized lymphadenopathy and splenomegaly starting from 3 weeks of age and being evident by 6–10 weeks of age, of which 20–30% BALB/c c-FLIPL Tg mice showed diarrhea (Figure 1a). In contrast, there was an age-dependent severe thymic atrophy in these mice (Figure 1b). The spleens from 7-week-old BALB/c c-FLIPL Tg mice showed that there was a significant increase in lymphoid cells, which formed aggregates in the white pulp and spread into the red pulp, filling in the cords and sinuses. PNA staining revealed a spontaneous germinal center (GC) formation in BALB/c c-FLIPL Tg mice but not in NLCs (Figure 1b).

BALB/c c-FLIPL Tg mice develop lupus-like disease. (a) Splenomegaly and lymphadenopathy in BALB/c c-FLIPL Tg mice. BALB/c c-FLIPL Tg mice were smaller than NLCs in size and showed splenomegaly and lymphadenopathy at 6–10 weeks of age. Pictures are representative of five mice analyzed for each group. (b) A disorganized structure of the spleen in BALB/c c-FLIPL Tg mice. Expansions of the white pulp were observed in the spleen of a BALB/c c-FLIPL Tg mouse with a spontaneous GC formation. Sections of spleens from 6-week-old BALB/c c-FLIPL Tg mice and NLCs were stained with HE, or immunostained with biotinylated PNA, followed by ABC-AP for GC (blue) and FITC-labeled anti-B220, followed by HRP-conjugated anti-FITC for B cells (brown). (c) Multiorgan infiltration. Lymphocyte infiltrations were observed in the liver, heart, stomach, large intestine, lung, and salivary gland of BALB/c c-FLIPL Tg mice at 7 weeks of age. Original magnification, × 100. (d) Histopathological analysis of kidneys. Sections of kidneys from 6-week-old BALB/c c-FLIPL Tg mice and NLCs were stained with PAS and observed under microscopy. Representative histopathology of c-FLIPL-transgenic glomeruli-exhibiting proliferative glomerulonephritis. Original magnification, × 400. (e) Immunohistochemical analysis of kidneys. Frozen sections of kidneys from the same animals as in D were stained with FITC-conjugated anti-mouse IgG Ab or FITC-conjugated anti-mouse CD3 Ab. Sections were observed under fluorescence microscopy. Original magnification, × 400. (f) Spontaneous auto-Ab production in BALB/c but not B6 c-FLIPL Tg mice. Sera were obtained from 6- to 10-week-old BALB/c or B6 c-FLIPL Tg mice and NLCs. Anti-ssDNA, anti-dsDNA, and anti-ANA Abs were measured by ELISA
Histological analysis showed evident lymphocytic infiltration in the portal tracts of the liver, heart valves, peri-bronchiolar spaces of the lung, and the mucosa and submucosa of the stomach and the large and small intestines. Salivary glands from BALB/c c-FLIPL Tg mice also showed dense lymphocyte infiltration (Figure 1c). Kidney sections from BALB/c c-FLIPL Tg mice showed a diffuse proliferative glomerulonephritis in BALB/c c-FLIPL Tg mice, characterized by enlarged glomeruli, loss of Bowman's capsule space, thickening of the mesangium, and hypercellularity. Immunofluorescence staining showed IgG and C3 deposition in glomeruli (Figure 1d-e). Serological analysis showed that BALB/c c-FLIPL Tg mice had markedly elevated levels of antinuclear, anti-ssDNA, and anti-dsDNA Abs, whereas B6 c-FLIPL Tg mice were negative for these auto-Abs (Figure 1f). Taken together, our results suggest that overexpressing c-FLIPL in T cells leads to the development of systemic lupus-like disease, which was contingent on the BALB/c background. The susceptible c-FLIPL Tg background requires the host background to trigger autoimmunity.
c-FLIPL overexpression leads to impaired thymic selection in a BALB/c background
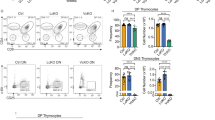
The difference observed in susceptibility to disease development in BALB/c c-FLIPL Tg mice as compared with B6 c-FLIPL Tg mice may result from the differences in T-cell development in these two strains. Development of autoimmunity could be because of the breakdown of self-tolerance in the thymus and/or changes in the activation thresholds of peripheral lymphocytes. It has been documented that thymocyte development is not altered in B6 c-FLIPL Tg mice.6 To analyze whether a defect in T-cell development might be responsible for disease development in BALB/c mice overexpressing c-FLIPL in T cells, we stained thymocytes from BALB/c c-FLIPL Tg and NLCs (1 week of age) with fluorescence-conjugated Abs against CD4 and CD8 in combination with CD25 and CD44 to define thymocyte development. BALB/c c-FLIPL Tg thymocytes developed successfully from the double-negative (DN) to the double-positive (DP) stage (data not shown). Although total thymocyte numbers in BALB/c c-FLIPL Tg mice were normal at 1 week of age, there were significant reductions in the percentages and absolute numbers of CD4 single-positive (SP) thymocyte populations (Figure 2a; Table 1), suggesting that their differentiation from DP to CD4 SP thymocytes was compromised. At 7 weeks of age, the total thymocyte numbers of BALB/c c-FLIPL Tg mice were reduced to about 20% of that of NLCs. The reduced thymocyte numbers were the result of smaller DP (31% of control) and CD4+ SP (65% of control) populations (Figure 2a; Table 1). The involuted thymus in older BALB/c c-FLIPL Tg mice may be secondary to stress. In sharp contrast, B6 c-FLIPL Tg mice showed normal distributions of DP and SP thymocytes (Supplementary Figure S2a) and no thymic atrophy (data not shown). These data suggest that overexpression of c-FLIPL impairs thymocyte development in a strain-dependent manner.

Thymic selection is impaired in BALB/c c-FLIPL Tg mice. (a) CD4+ SP thymocytes are selectively reduced in BALB/c c-FLIPL Tg mice. The freshly isolated thymocytes from c-FLIPL Tg mice and their NLCs at 1 and 7 weeks of age were stained with Abs against CD4 and CD8. (b) BALB/c c-FLIPL Tg thymocytes failed to upregulate CD69 and CD5. Thymocytes from BALB/c c-FLIPL Tg mice and NLCs at 1 week of age were stained with Abs against CD4, CD8, CD69, and CD5. The expression of CD5 and CD69 in DP thymocytes from BALB/c c-FLIPL Tg mice and NLCs was determined. (c) Overexpression of c-FLIPL impairs both positive and negative selection in the thymus. DO11.10.c-FLIPL Tg mice and DO11.10 mice (at 2 weeks of age) were injected i.p. with 30 or 300 μg OVAp323−339 peptide every day for 3 days. Thymocytes from these mice were stained with KJ1-26 and anti-CD8, and analyzed by flow cytometry. (d) VSAg-mediated deletion is impaired in BALB/c but not B6 c-FLIPL Tg mice. Thymocytes from B6 and BALB/c c-FLIPL Tg and their NLCs at 1 week of age were stained with Abs against CD4, CD8, Vβ3, Vβ5, Vβ11, and Vβ12, and analyzed by flow cytometry
To obtain further insight into the mechanism underlying altered thymic development, we analyzed whether the reduction in the numbers of CD4 SP thymocytes was caused by a defect in positive selection. During positive selection, CD69 expression is induced, and CD5 is upregulated.10 As shown in Figure 2b, BALB/c c-FLIPL Tg thymocytes failed to upregulate the expression of CD69 and CD5, suggesting that thymic positive selection is impaired in the presence of c-FLIPL overexpression. To further facilitate the examination of positive selection, we established a TCR Tg system to specifically study the effect of the overexpression of c-FLIPL on the development of these cells. This was accomplished by breeding the I-Ad-restricted DO11.10 TCR transgene, which is specific for a peptide from chicken ovalbumin (OVA) (amino acids 323–339; OVAp323−339)11, 12 onto BALB/c c-FLIPL Tg mice. Positive selection of the CD4 lineage was analyzed with the use of KJ1–26 anti-idiotypic mAb, which recognizes DO11.10 TCR. The analysis indicated that there was a significant decrease in the percentage of CD4 SP thymocytes in DO11.10.c-FLIPL Tg mice (Figure 2c, upper panel); further showing that positive selection of the CD4 lineage was impaired.
Negative selection of thymocytes is a major mechanism of central tolerance. It is possible that the autoimmune lupus-like disorder observed in c-FLIPL Tg mice is because of the breakdown of central tolerance. To substantiate the role of c-FLIPL in thymic negative selection, we systemically administrated OVAp323−339 peptide into OVA-specific TCR Tg DO11.10 and DO11.10.c-FLIPL Tg mice. Injection of OVAp323−339 peptide into DO11.10 mice reduces both the percentage and the absolute number of immature Tg thymocytes.13 We found that administration of OVAp323−339 peptide did induce a deletion of DP thymocytes in NLCs in a dose-dependent manner, but this deletion was significantly impaired in BALB/c c-FLIPL Tg mice (Figure 2c, middle and lower panels), suggesting that thymic negative selection is also impaired in BALB/c c-FLIPL Tg mice.
To confirm the effect of c-FLIPL overexpression in thymic selection in a strain-dependent manner, we first measured the expression of CD5 and CD69 in DP thymocytes from B6 c-FLIPL Tg mice and their NLCs. There was no difference in the CD5 and CD69 expression between B6 c-FLIPL Tg and their NLC DP thymocytes (Supplementary Figure S2b). To further verify c-FLIPL overexpression in thymic selection in a B6 strain, we bred B6 c-FLIPL Tg mice onto OT-II mice in which T cells express a TCR specific for peptide 323–339 of OVA in the context of I-Ab.12 We then treated OT-II and OT-II.c-FLIPL Tg mice with OVAp323−339 peptide or with PBS, and monitored the positive and negative selection of thymocytes in these mice. As shown in Supplementary Figure 2c, overexpressing c-FLIPL in OT-II T cells did not result in any significant impairment in both positive and negative selection. Our data indicate that c-FLIPL overexpression may affect the threshold of TCR avidity for the self-peptide/MHC ligands required for each selection event under BALB/c but not B6 background, leading to aberrant thymic selection in the BALB/c strain.
The model of negative selection described above depends on the activation of exogenous TCR ligand in the system. To determine whether negative selection induced by endogenous antigens is also affected by the c-FLIPL transgene, we analyzed the repertoire of T cells reactive to endogenous mouse mammary tumor virus (MMTV) products presented on an I-E molecule. BALB/c mice have the integration of MMTVs (Mtv types 6, 8, and 9) in the genome and normally delete the viral super-antigen (VSAg)-reactive TCR Vβ3-, 5-, 11-, and 12-positive T cells, whereas B6 mice fail to delete them because of the lack of MHC class II I-E molecules.14, 15 As shown in Figure 2d, TCR Vβ3-, 5-, 11-, and 12-positive cells were significantly deleted in thymi from NLCs, but this deletion was inhibited in BALB/c and not in B6 c-FLIPL Tg mice. Taken together, our data suggest that thymic negative selection is impaired in BALB/c mice overexpressing c-FLIPL. It is to be noted that DO11.10.c-FLIPL Tg mice developed a lupus-like syndrome at high incidence (Supplementary Figure S3), suggesting that disease development in these mice is possibly because of the expression of endogenous TCRs.
BALB/c c-FLIPL Tg T and B cells show activated phenotypes
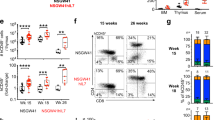
It has been shown that c-FLIPL Tg mice in a B6 background do not show an activated phenotype in their T and B cells.6 Having examined thymocyte development/selection in BALB/c c-FLIPL Tg mice, we wanted to determine the phenotype of peripheral T cells in BALB/c c-FLIPL Tg mice. Although the spleens and lymph nodes were enlarged in BALB/c c-FLIPL Tg mice, T cells were significantly reduced in the spleen and lymph nodes of BALB/c c-FLIPL Tg mice compared with those of NLCs at 7 weeks of age (Figure 3a and b; Table 1). The ratio of T to B cells was abnormally increased in the spleens and lymph nodes, suggesting that enlarged secondary lymphoid tissues are not caused by expanded T-cell compartments of BALB/c c-FLIPL Tg mice. A significant fraction of CD4+ and CD8+ T cells from spleens and lymph nodes of BALB/c c-FLIPL Tg mice at 7 weeks of age expressed high levels of CD25, CD69, and CD44, but low levels of CD62L (Figure 3a and b), suggesting that they had recently been stimulated by antigens and were activated effector T cells. Therefore, splenomegaly and lymphoadenopathy observed in BALB/c c-FLIPL Tg mice are likely to be secondary to aberrant effector T-cell activation. In support of this notion, the percentages and numbers of B cells, granulocytes, macrophages, and DCs were significantly increased in BALB/c c-FLIPL Tg mice compared with those in their NLCs (Table 1). We did not detect any B220+CD3+CD4−CD8− DN T cells that are typically present in lpr mice16 (data not shown). In addition, the expression of B7-1, B7-2, CD69, I-Ad, and Fas was increased in BALB/c c-FLIPL Tg B cells, suggesting that BALB/c c-FLIPL Tg B cells are also activated in vivo (Figure 3c). In contrast, B6 c-FLIPL Tg mice showed reduced CD8+ T cells in their spleens and lymph nodes (Supplementary Figure S4a), consistent with the earlier report.6 In addition, we also found that expression of activation markers such as CD25, CD62L, CD69, and Fas was comparable between B6 c-FLIPL Tg T cells and their NLC T cells (Supplementary Figure S4b). Therefore, development of autoimmunity requires a susceptible genetic background.

T and B cells of BALB/c c-FLIPL Tg mice show an activated phenotype. (a and b) The spleen and lymph node cells from c-FLIPL Tg mice and their NLCs at 7 weeks of age were stained with Abs against CD4, CD8, CD25, CD44, CD62 L, CD69, and Fas. (c) c-FLIPL Tg and spleen cells of NLC (7-week-old) were stained with Abs against B220 and CD3, or against B220 with I-Ad, B7-1, B7-2, Fas, or CD69. The percentages of CD4 and CD8 in spleen and lymph node cells and of B cells in the spleen, as well as the expression of activation markers in T cells and B cells, were detected by flow cytometry
To test whether CD4+ T cells are autoreactive in BALB/c c-FLIPL Tg mice, CD4+ T cells isolated from either NLC or c-FLIPL Tg mice at 2 weeks of age (before disease onset) were adoptively transferred in conjunction with either c-FLIPL Tg or NLC B cells into Rag-1−/− hosts (BALB/c background). Seven weeks later, Rag-1−/− mice were monitored for anti-dsDNA Ab production and lupus-like syndrome. Rag-1−/− mice reconstituted with BALB/c c-FLIPL Tg CD4+ T cells and BALB/c c-FLIPL Tg or NLC B cells showed an increase in anti-dsDNA Ab production and lymphocyte infiltration in multiorgans (Figure 4a and b). However, transfer of NLC CD4+ T cells in conjunction with either c-FLIPL Tg or NLC B cells failed to result in high anti-dsDNA Ab titers and remained disease free. FACS analysis clearly identified B-cell populations in Rag-1−/− hosts, suggesting that transfer of B cells, together with CD4+ T cells, could survive long enough in Rag-1−/− mice.17, 18 It is to be noted that B-cell populations were increased in Rag-1−/− recipients receiving c-FLIPL Tg CD4+ T cells compared with those receiving CD4+ T cells from NLCs (Figure 4c).

Production of auto-Abs and autoimmune syndrome in BALB/c c-FLIPL Tg mice requires CD4+ T cells. CD4+ T cells and B cells from BALB/c c-FLIPL Tg mice and NLCs at 2 weeks of age were injected into Rag-1−/− recipients (n=5) with different combinations as indicated. (a) The mice were killed 7 weeks later, and serum anti-dsDNA Abs were determined by ELISA. (b) The sections of heart, kidney, and liver were stained with H&E and observed under microscope. Original magnification, × 100. (c) The presence of T and B cells in spleens of Rag-1−/− recipients was determined by flow cytometry
Overexpression of c-FLIPL inhibits T-cell proliferation by promoting apoptosis, and leads to heightened Th2 cytokine production
The development of lupus-like syndrome in BALB/c c-FLIPL Tg mice could be because of an aberrant autoreactive T-cell proliferation in the periphery. It was earlier shown that overexpression of c-FLIPL in T cells in the B6 background may result in an increase in T-cell proliferation6, 19 (Supplementary Figure S5a and b). To verify the roles of c-FLIPL in T-cell proliferation, splenic T cells from BALB/c c-FLIPL Tg mice and NLCs at 2 weeks of age before disease onset were stimulated for 72 h with different concentrations of plate-bound anti-CD3 or anti-CD3 plus anti-CD28. Interestingly, the proliferation rate of BALB/c c-FLIPL Tg splenic T cells in response to TCR or TCR/CD28 stimulation was significantly lower than that in NLC splenic T cells (Figure 5a). Regardless of the hypo-responsiveness to TCR stimulation because of the overexpression of c-FLIPL, c-FLIPL Tg CD4+ T cells were twice as proliferative as NLC CD4+ T cells in the physiological state, as shown with in vivo 5-bromo-2-deoxyuridine (BrdU) incorporation (Figure 5b). Overexpressing c-FLIPL in the T-cell lineage did not affect CD4 SP thymocyte proliferation, indicating that the proliferating T cells may be because of homeostasis of peripheral T cells.

c-FLIPL overexpression inhibits T-cell proliferation and leads to a Th2-biased cytokine production. (a) Splenic CD4+ T cells from BALB/c c-FLIPL Tg mice and their NLCs (2-week-old) were stimulated with different concentrations of anti-CD3, or anti-CD3 (1 μg/ml) plus anti-CD28 (1 μg/ml), for 72 h and proliferation of T cells was measured by [3H]thymidine incorporation. (b) BALB/c c-FLIPL Tg mice and NLCs at 2 weeks of age were administered with BrdU for 3 days, and percentages of BrdU-stained cells among CD4+ SP thymocytes and CD4+ T cells are shown. (c) Splenic CD4+ T cells from BALB/c c-FLIPL Tg mice and their NLCs (2-weeks-old) were labeled with CFSE, and cultured in the plate pre-coated with anti-CD3 for 72 h. The cells were washed, and stained with PE-annexin V. (d) T cells from BALB/c c-FLIPL Tg mice and their NLCs were stimulated with anti-CD3, anti-CD3 plus anti-CD28 as in A. Supernatants were collected at 48 h after stimulation and the production of IL-2, IL-4, IFN-γ, and IL-17 was detected by mouse cytokine 20-plex assay
We found that BALB/c c-FLIPL Tg CD4+ T cells are hypo-proliferative in vitro but not in vivo, and CD4+ T cells are dramatically reduced in BALB/c c-FLIPL Tg mice. These observations raised a possibility that BALB/c c-FLIPL Tg CD4+ T cells are highly susceptible to apoptosis. To assess proliferation and apoptosis simultaneously, we labeled CD4+ T cells of BALB/c c-FLIPL Tg and NLC mice with carboxyfluorescein succinimidyl ester (CFSE), and stimulated these T cells with anti-CD3. The apoptosis in proliferating or non-proliferating T cells was determined by staining them with PE-conjugated annexin V. As shown in Figure 5c, a significant portion of BALB/c c-FLIPL Tg CD4+ T cells that did not enter cell cycle underwent apoptosis. Consistent with [3H]thymidine incorporation results, BALB/c c-FLIPL Tg CD4+ T cells proliferated poorly in response to CD3 stimulation (Figure 5c). Therefore, the reduced peripheral T cells may be at least in part because of cell cycle arrest and increased sensitivity to apoptosis in BALB/c c-FLIPL Tg mice. In sharp contrast, B6 c-FLIPL Tg T cells highly proliferated in response to lower doses of anti-CD3 stimulation, although they showed a similar sensitivity to apoptosis (Supplementary Figure S5b), in agreement with earlier reports.6, 19
One of the principal responses of activated T cells is the production of cytokines. To test whether activated T cells from c-FLIPL Tg mice were able to secrete effector cytokines, we stimulated CD4+ T cells from c-FLIPL Tg mice and NLCs with anti-CD3 or with anti-CD3 plus anti-CD28 for 72 h as described before, and the production of Th1, Th2, and Th17, as well as that of proinflammatory cytokines and chemokines, was measured by Cytokine 20-Plex in the supernatants collected. As shown in Figure 5d and Supplementary Figure S5c, overexpression of c-FLIPL in T cells selectively led to a significant increase in the production of Th2 cytokines, especially IL-4, detected on TCR or TCR/CD28 stimulation, consistent with earlier reports.20, 21 In support of this observation, serum IgG1 and IgE production was significantly elevated in BALB/c c-FLIPL Tg mice (data not shown).
Overexpressing c-FLIPL in T cells in a BALB/c background impairs TCR-induced ZAP-70 activation and downstream signaling pathways in thymocytes
Several studies reveal that a point mutation at ZAP-70 (ZAP-70W163C) or at the linker for activation of T cells (LAT) (LATY136F) gene leads to the development of autoimmune arthritis or systemic autoimmunity as a result of altered thymic selection.22, 23 To test whether overexpression of c-FLIPL interferes with proximal TCR signaling, we measured activation of Lck, ZAP-70, TCR-ζ, and LAT in thymocytes from BALB/c and B6 c-FLIPL Tg and NLC in response to TCR stimulation. Intriguingly, we found that although Lck Y394 phosphorylation was comparable between BALB/c c-FLIPL Tg and NLC thymocytes, phosphorylation of ZAP-70, TCR-ζ, and LAT was significantly reduced (Figure 6a). However, we did not observe a similar reduction in the phosphorylation of ZAP-70, TCR-ζ, and LAT in B6 c-FLIPL Tg thymocytes after TCR stimulation (Supplementary Figure 6a). These observations suggest that overexpression of c-FLIPL may selectively impair the formation of TCR signalosome in a BALB/c background, which connects events on the plasma membrane to distal signaling cascades.24 To test this idea, we assessed the assembly of TCR signalosome in thymocytes from BALB/c c-FLIPL Tg mice and their NLCs. As expected, TCR-induced signaling complex formation consisting of ZAP-70, TCR-ζ, LAT, and SH2-domain-containing leukocyte protein of 76 kDa (SLP-76) was defective in thymocytes overexpressing c-FLIPL in BALB/c mice (Figure 6b). These data suggest that c-FLIPL overexpression may constrain ZAP-70 in the cytosol, thus inhibiting the translocation of ZAP-70 from the cytosol to the plasma membrane. To verify this possibility, we isolated cytosol and membrane fractions from BALB/c c-FLIPL Tg mice and NLCs before and after TCR stimulation by subcellular fractionation.25 The amounts of ZAP-70, LAT, SLP-76, and phospholipase-gamma1 (PLC-γ1) were significantly reduced in the membrane fraction from BALB/c c-FLIPL Tg thymocytes compared with that from NLC thymocytes after TCR stimulation (Figure 6c). Therefore, our data support the notion that overexpressing c-FLIPL in T cells may impair the ZAP-70-mediated signaling pathway, thus attenuating TCR signaling responsible for thymic selection.

c-FLIPL overexpression impairs the signaling pathway derived from ZAP-70 BALB/c c-FLIPL Tg thymocytes. (a) c-FLIPL overexpression results in defective phosphorylation of ZAP-70, TCR-ζ, and LAT in thymocytes of BALB/c c-FLIPL Tg mice. Thymocytes from BALB/c c-FLIPL Tg mice and their NLCs (2 weeks of age) were stimulated with anti-CD3, and lysed. The cell lysates were immunoblotted with phospho-Abs against ZAP-70 (Y493), and LAT (Y191), or immunoprecipitated with anti-Lck or anti-TCR-ζ, and blotted with anti-phospho-src-PTK (Y416), which recognizes Lck Y394 or anti-phospho-tyrosine mAb (4G10). (b) c-FLIPL overexpression impairs the formation of TCR signalsome. Thymocytes from BALB/c c-FLIPL Tg mice and their NLCs (2 weeks of age) were stimulated with anti-CD3, and lysed in 0.5% NP40 lysis buffer. The cell lysates were immunoprecipitated with anti-ZAP-70, and blotted with anti-TCR-ζ, anti-LAT, and anti-SLP-76. (c) Overexpression of c-FLIPL in T cells inhibits the recruitment of ZAP-70, LAT, SLP-76, and PLC-γ1 to the plasma membrane. Thymocytes from BALB/c c-FLIPL Tg mice and their NLCs (2 weeks of age) were stimulated with anti-CD3, and the membrane and cytosol fractions were isolated. The expression of TCR-ζ, ZAP-70, LAT, SLP-76, and PLC-γ1 in the membrane and cytosol fractions was detected by immunoblotting
The signaling events downstream of the TCR have a critical role in positive and negative selection. To further determine the cause of the defect in positive and negative selection of BALB/c c-FLIPL Tg thymocytes, we assessed their responses to TCR stimulation. In particular, we examined mitogen-activated protein kinase (MAPK)- and NF-κB-pathways, as these TCR responses have an essential role in thymic selection26, 27, 28, 29 We found a significant reduction in the phosphorylation of excellular signal-regulated kinase (ERK) and IκBα on TCR stimulation, whereas phosphorylation of c-Jun-NH2-terminal kinase (JNK) and p38 MAPK was comparable between BALB/c c-FLIPL Tg and NLC thymocytes in response to TCR stimulation (Figure 7). These data suggest that c-FLIPL overexpression impairs TCR-induced activation of the ZAP-70-mediated signaling pathway, which subsequently results in an altered activation of ERK and NF-κB responsible for thymic selection, thus eliciting lupus-like disease in BALB/c mice. In contrast, activation of MAPKs including ERK, JNK, and p38, as well as NF-κB, was comparable in the presence or absence of the c-FLIPL transgene in a B6 background (Supplementary Figure S6b).

TCR-induced activation of ERK and NF-κB is defective in BALB/c c-FLIPL Tg thymocytes. Thymocytes from BALB/c c-FLIPL Tg mice and their NLCs (2 weeks of age) were stimulated with anti-CD3, and lysed. The cell lysates were blotted with phospho-Abs against ERK (Thr282/Tyr284), JNK (T183/Y185), p38 MAPK (T180/Y182), and IκBα (S32). The membranes were stripped and reprobed with anti-actin as loading controls
Discussion
In this study, we first report that BALB/c c-FLIPL Tg mice spontaneously develop lupus-like disease starting from 3 to 4 weeks of age, characterized by splenomegaly, lymphadenopathy, multiorgan infiltration including liver, lung, gastrointestine, heart, and salivary gland, proliferative glomerulonephritis with immune complex deposition, and high titers of auto-Abs (Figure 1). These features are clinically similar to human SLE patients. BALB/c c-FLIPL Tg T and B cells show activated phenotypes (Figure 3), suggesting that they are hyper-activated in vivo. The development of the disease requires CD4+ T cells, and may be because of impaired thymic selection. Thus, the BALB/c c-FLIPL Tg strain provides evidence of a key role for c-FLIPL signals in the maintenance of immunological self-tolerance. Our data are consistent with reports that the elevated c-FLIP expression was observed in human T cells in patients with SLE,30, 31 although the exact role of c-FLIP in SLE development in human remains to be elucidated.
The development of autoimmunity may result from a breakdown of central or peripheral tolerance. Autoreactive T cells are deleted in the thymus by negative selection. Recently, the defect in positive selection was reported to cause autoimmunity.32 Moreover, a point mutation at the ZAP-70 (ZAP-70W163C) or LAT (LATY136F) gene alters both positive and negative selection in the thymus, eliciting either autoimmune arthritis or systemic autoimmunity.22, 23 In keeping with this, targeted disruption of Gads, as well as the deletion of the Gads binding site in SLP-76, impairs both positive and negative selection.33 Moreover, in the Lcp2twp strain, an ENU-induced mutation in a splice donor site reduces the amount of full-length mRNA encoding SLP-76 and promotes autoimmunity.3 The failure to upregulate CD69 and CD5 in DP thymocytes of BALB/c c-FLIPL Tg mice suggests a defect in positive selection. Indeed, a similar reduction of CD4 SP thymocytes was also seen in DO11.10.c-FLIPL Tg mice (Figure 2c, upper panel), supporting a defect in positive selection in these mice. Using two model systems, VSAg-mediated deletion and systemic administration of the OVA peptide to DO11.10.c-FLIPL Tg mice, we showed that thymic negative selection is also impaired in BALB/c mice overexpressing c-FLIPL (Figure 2c, middle and lower panels, and Figure 2d). Taken together, our data indicate that c-FLIPL overexpression in T cells in a BALB/c background leads to altered thymic selection, resulting in positive selection of otherwise negatively selecting self-reactive T cells in the thymus.
Mechanistically, overexpressing c-FLIPL in T cells may impair the ZAP-70-mediated signaling pathway, thus attenuating the TCR signaling responsible for thymic selection. Consistent with the altered thymic selection in BALB/c c-FLIPL Tg mice, activation of ERK and NF-κB was impaired in c-FLIPL Tg thymocytes after TCR stimulation (Figure 7). The attenuated activation of ERK and NF-κB would impair both positive and negative selections of T cells in BALB/c c-FLIPL Tg mice.33, 34 As c-FLIPL may heterodimerize with caspase-8 on TCR stimulation, and caspase-8 may mediate NF-κB activation in a CARMA-1-dependent manner,35, 36 it is presumed that TCR-induced c-FLIPL-caspase-8 heterodimer formation may interfere with the role of caspase-8 in NF-κB activation. However, c-FLIPL-mediated inhibition of NF-κB activation in the thymus remains to be further determined, as deficiency for caspase-8, CARMA-1, Bcl-10, and MALT-1 does not perturb thymic selection.37, 38
It has been documented that c-FLIPL inhibits FasL-induced cell death.39 However, c-FLIPL has also been suggested to be crucial for T-cell proliferation and differentiation 6, 7, 8, 21 As c-FLIPL is an endogenous inhibitor of caspase-8, it was expected that mice overexpressing c-FLIPL in T cells would lead to the development of lupus-like disease as seen in lpr mice. Unexpectedly, mice overexpressing c-FLIPL in T cells in a B6 background do not succumb to lupus-like syndrome.6, 21, 39 In contrast to this, retrovirus-mediated overexpression of c-FLIPL in T and B cells blocks Fas-induced apoptosis of activated T and B cells, which leads to the production of auto-Abs and to the development of autoimmune diseases;40 however, the contribution of c-FLIPL-overexpressing T cells versus B cells in disease development is not defined. We found that c-FLIPL Tg mice in the BALB/c background spontaneously develop lupus-like disease (Figure 1). The development of the disease is mediated by CD4+ T cells from BALB/c c-FLIPL Tg mice because adoptive transfer of CD4+ T cells from BALB/c c-FLIPL Tg mice but not from NLCs, together with B cells from either BALB/c c-FLIPL Tg or NLC mice into Rag-1−/− recipients causes the disease (Figure 4a and b). Our data also suggest that c-FLIPL overexpression does not inhibit T-cell apoptosis in vitro, but rather results in cell cycle arrest and aberrant apoptosis, thus suppressing CD3-triggered T-cell proliferation (Figure 5a and c). The reduced export of CD4+ SP thymocytes from BALB/c c-FLIPL Tg mice to the periphery may result in heightened homeostatic proliferation in the periphery as shown by increased BrdU incorporation in splenic CD4+ T cells in these mice (Figure 5b).
The restrictive function of c-FLIPL further explains the requirement for genetic modifiers for the emergence of autoimmunity in mice overexpressing this molecule in T cells. Autoimmunity appears when multiple immunological pathways are perturbed. In general, single gene knockouts or transgenes that result in autoimmunity have pleiotropic effects on T- and B-cell function, as well as in antigen presentation. Specific genetic backgrounds may provide the additional co-activating components necessary for the development of autoimmune disease. This notion is supported by the fact that FcγRIIB−/− mice develop lupus-like autoimmune diseases in B6 but not in a BALB/c background,41 suggesting that spontaneous autoimmune disease in FcγRIIB−/− mice results from strain-specific epistasis. Our data indeed support the notion that overexpression of c-FLIPL in T cells is not sufficient to trigger autoimmunity, and the development of a lupus-like autoimmune phenotype requires the trigger from a host genetic background. We believe that these two factors together affect the threshold for thymic selection that results in aberrant positive and negative selection in the thymus in BALB/c but not in B6 background. Indeed, thymic selection is intact in OT-II.c-FLIPL Tg mice (Supplementary Figure S2). Moreover, the c-FLIPL transgene does not perturb proximal TCR signaling in the B6 background as seen in BALB/c c-FLIPL Tg mice (Supplementary Figure S6a). Taken together, these data suggest that c-FLIPL overexpression in T cells may alter the threshold for T-cell activation in the BALB/c but not in the B6 strain required for thymic selection. The nature of these genetic factors that influence c-FLIPL function is currently under investigation.
In conclusion, BALB/c c-FLIPL Tg mice spontaneously develop an autoimmune disorder showing many similarities to human SLE, providing a novel model of autoimmune disease in a strain-specific epistasis. The development of the disease is because of altered thymic selection. c-FLIPL is thus a susceptibility factor in the development of systemic autoimmune disease, contributing in a combinatorial manner through the presence of interacting genetic factors to the emergence of disease. Further study of BALB/c c-FLIPL Tg mice will improve our understanding of the complex network of signals that serve to maintain self-tolerance and will improve our insight into the genetic basis of autoimmune disease.
Materials and Methods
Mice
c-FLIPL was expressed transgenically in the T-cell compartment as reported earlier.6, 39 Briefly, FLAG-tagged mouse c-FLIPL cDNA was inserted into a target vector containing the β-globin promoter and a downstream human CD2 locus enhancer element. B6 c-FLIPL Tg mice were backcrossed onto a BALB/c background for 14 generations. DO11.10 mice were purchased from the Jackson Laboratory (Bar Harbor, ME, USA). BALB/c c-FLIPL Tg mice were crossed onto DO11.10 mice to generate DO11.10.c-FLIPL Tg mice. B6 c-FLIPL Tg mice were crossed onto OT-II mice, which were purchased from the Jackson Laboratory, to generate OT-II.c-FLIPL Tg mice. All animals were used in accordance with the National Institutes of Health guidelines. The experiments described in this study were reviewed and approved by the University of Chicago Institutional Animal Care and Use Committee. Mice were housed at the University of Chicago Animal Care facility. The expression of the c-FLIPL transgene in thymocytes, splenic T cells, B cells, and DCs was detected by western blotting with anti-FLAG and anti-c-FLIPL (Dave-2).
Histopathology
Tissue specimens for histopathology were fixed in 4% paraformaldehyde (Fisher Scientific, Pittsburgh, PA, USA) or were snap frozen. Tissue sections (5 μm) were stained with hematoxylin and eosin (H&E) and evaluated by light microscopy. Immunohistochemical analysis on frozen kidney sections was performed on acetone-fixed sections. In brief, sections were blocked with 10% goat serum and, in some cases, with 0.1% Triton X-100, followed by fluorescein-conjugated goat anti-mouse IgG (Jackson Immunotech, West Grove, PA, USA) for 1 hr at room temperature. After washing, immunofluorescence analysis was performed using a Nikon Eclipse E600 microscope (Nikon, Meville, NY, USA).
Purification of T cells, B cells, and DCs
CD4+ T cells were purified from the spleen by negative selection (>90% CD4+) (Miltenyi Biotec, Auburn, CA, USA). B cells were isolated (purity>95% as determined by FACS analysis of B220 cell surface expression) using a B-cell isolation kit using negative selection (Miltenyi Biotec.). Contaminating T cells were <1%, as determined by CD3 staining. DCs were isolated from spleen by positive selection (>90% DCs as determined by FACS analysis of CD11c) (Miltenyi Biotec.).
Cell activation and lysis
Thymocytes were maintained on ice in RPMI-1640 supplemented with 20 mM Hepes until stimulation. Quiescent thymocytes (4 × 107/ml) were stimulated with hamster anti-mouse CD3 mAb (1 μg/ml) on ice for 30 min, and washed in prewarmed medium, and crosslinked with anti-hamster IgG (10 μg/ml) for various times. The cells were lysed in 1% Triton X-100 lysis or in 0.5% NP-40 lysis buffer as indicated.
Adoptive transfer
CD4+ T cells (5 × 106) and B cells (107) from BALB/c c-FLIPL Tg mice and NLCs at 2 weeks of age were i.v. injected into Rag-1−/− recipients (n=5) with different combinations as indicated. The mice were killed 7 weeks later, and serum anti-dsDNA Abs were determined by ELISA. Different organs were collected for histopathological analysis.
Detection of auto-Abs by ELISA
Anti-nuclear Abs (ANA) were detected by ELISA. Auto-Abs to double-stranded DNA (dsDNA), ssDNA, and nuclei were determined by ELISA as described earlier.42
Flow cytometry
The following Abs (Pharmingen, San Diego, CA, USA) were used: anti-CD4 (clone GK1.5), anti-CD8 (53-6.7), KJ1-26, anti-Vβ3, anti-Vβ5, anti-Vβ11, anti-Vβ12, anti-B7-1 (16-10A1), anti-B7-2(GL1), and anti-B-220 (RA3-6B2) for lineage markers; anti-CD69 (H1.2F3), anti-CD44 (IM7), anti-CD62L (MEL-14), anti-Fas (Jo2), anti-CD5 (53-7.3), and anti-CD25 (PC81) for activation markers. Acquisition of samples was performed on a FACScan flow cytometer, and data were analyzed using FlowJo software V.4.4.3 (Tree Star Inc., Ashland, OR, USA).
In vivo administration of OVAp323−339
DO11.10 and DO11.10.c-FLIPL Tg or OT-II and OT-II.c-FLIPL Tg mice (2 weeks of age) were injected intraperitoneally with PBS or OVAp323−339 once a day at doses of 30 μg or 300 μg/mouse. Mice were killed after 72 h, and their thymi were removed for analysis.
Assays for cell proliferation and apoptosis
T cells were stimulated with anti-CD3 or anti-CD3 and anti-CD28 for the indicated times. Proliferation was assayed by measuring [3H]thymidine incorporation. Alternatively, purified CD4+ T cells were labeled with CFSE (Molecular Probes; Eugene, OR, USA) at 5 μM in PBS for 10 min at 37 °C, washed with RPMI-containing 10% FCS, and incubated with plate-bound anti-CD3 for 72 h. The cells were then washed, stained with PE-labeled annexin V, and analyzed by flow cytometry. For in vivo BrdU labeling, mice were i.p. injected with 1.0 mg BrdU (Sigma-Aldrich, St. Louis, MO, USA) twice every 12 h until flow cytometric analysis.
Cytokine assay by luminex
Mouse cytokine 20-plex assay kits (Invitrogen, Carlsbad, CA, USA) were used to measure the concentrations of cytokines in the supernatants collected from T-cell cultures according to the manufacturer's instruction. Concentrations of analyte in samples were determined by interpolation from 20 individual standard curves, using the five-parameter curve fitting provided in Bio-Plex Manager software version 3.0 (Bio-Rad, Hercules, CA, USA).
Subcellular fractionation
Thymocytes (108) were resuspended and lysed by brief sonication in ice-cold 10 mM Tris, pH 7.4, 10 mM KCl, 1.5 mM MgCl2 and 2 mM EGTA hypotonic buffer containing the above-described mixture of protease and phosphatase inhibitors (buffer A). Lysates were adjusted to 150 mM NaCl, centrifuged to remove nuclei and debris, and particulate membrane-containing (P100) and soluble cytoplasm-containing (S100) fractions were separated by differential centrifugation for 30 min at 100 000 g.43 Membrane fractions were washed with ice-cold buffer A, and solubilized by sonication in buffer A supplemented with 150 mM NaCl and 1% Triton X-100.
Immunoprecipitations of cellular proteins
Precleared postnuclear fractions obtained from 4 × 107 cells were normalized for protein concentration levels and immunoprecipitated (3 h at 4 °C) with the specific polyclonal Abs or control isotype-matched preimmune Ig precoupled to 30 μl of protein A-Sepharose CL-4B (GE Healthcares Systems, Piscataway, NJ, USA). This was followed by four washes of the precipitates with ice-cold lysis buffer.
Western blot analysis
The thymocyte lysates from NLCs and BALB/c c-FLIPL Tg mice were blotted with phospho-Abs against Lck (Y394), ZAP-70 (Y493), LAT (Y191), PLC-γ1 (Y783), ERK (T202/Y204), JNK (T183/Y185), p38 MAPK (T180/Y182), or IκB(S32) (Cell Signaling Technology, Inc., Beverly, MA, USA), and anti-actin (Sigma-Aldrich). Precipitated proteins or the membrane and cytosol fractions were solubilized in 2 × Laemmli sample buffer-containing 2-ME, 20 mM EDTA, and 2 mM Na3VO4, resolved by SDS-PAGE under reducing conditions, transferred to immobilon (Millipore, Bedford, MA, USA) or nitrocellulose (Schleicher & Schuell, Keene, NH, USA) membranes, and immunoblotted with the indicated Abs.
Conflict of interest
The authors declare no conflict of interest.
Abbreviations
- Ab:
-
antibody
- B6:
-
C57BL/6
- BrdU:
-
5-bromo-2-deoxyuridine
- c-FLIP:
-
cellular FLICE-inhibitory protein
- CFSE:
-
carboxyfluorescein succinimidyl ester
- DC:
-
dendritic cells
- DN:
-
double negative
- DP:
-
double positive
- ERK:
-
excellular signal-regulated kinase
- JNK:
-
c-Jun-NH2-terminal kinase
- LAT:
-
the Linker for activation of T cells
- MAPK:
-
mitogen-activated protein kinase
- MMTV:
-
mouse mammary tumor virus
- NLC:
-
non-Tg littermate controls
- OVA:
-
ovalbumin
- PLC-γ1:
-
phospholipase C-gamma-1
- SLE:
-
systemic lupus erythematosus
- SLP-76:
-
SH2-domain-containing leukocyte protein of 76 kDa
- SP:
-
single positive
- Tg:
-
transgenic
- TCR:
-
T-cell receptor
- VSAg:
-
viral super-antigen
- ZAP-70:
-
ζ chain-associated protein tyrosine kinase of 70 kDa
References
Hogquist KA, Baldwin TA, Jameson SC . Central tolerance: learning self- control in the thymus. Nat Rev Immunol 2005; 5: 772–782.
Palmer E, Naeher D . Affinity threshold for thymic selection through a T-cell receptor-co-receptor zipper. Nat Rev Immunol 2009; 9: 207–213.
Liston A, Enders A, Siggs OM . Unravelling the association of partial T-cell immunodeficiency and immune dysregulation. Nat Rev Immunol 2008; 8: 545–558.
Micheau O, Lens S, Gaide O, Alevizopoulos K, Tschopp J . NF-κB signals induce the expression of c-FLIP. Mol Cell Biol 2001; 21: 5299–5305.
Budd RC, Yeh WC, Tschopp J . cFLIP regulation of lymphocyte activation and development. Nat Rev Immunol 2006; 6: 196–204.
Lens SM, Kataoka T, Fortner KA, Tinel A, Ferrero I, MacDonald RH et al. The caspase 8 inhibitor c-FLIPL modulates T-cell receptor-induced proliferation but not activation-induced cell death of lymphocytes. Mol Cell Biol 2002; 22: 5419–5433.
Zhang N, He YW . An essential role for c-FLIP in the efficient development of mature T lymphocytes. J Exp Med 2005; 202: 395–404.
Chau H, Wong V, Chen NJ, Huang HL, Lin WJ, Mirtsos C et al. Cellular FLICE-inhibitory protein is required for T cell survival and cycling. J Exp Med 2005; 202: 405–413.
Zhang J, Xu X, Liu Y . Activation-induced cell death in T cells and autoimmunity. Cell Mol Immunol 2004; 1: 186–191.
Azzam HS, Grinberg A, Lui K, Shen H, Shores EW, Love PE . CD5 expression is developmentally regulated by T cell receptor (TCR) signals and TCR avidity. J Exp Med 1998; 188: 2301–2311.
Murphy KM, Heimberger AB, Loh DY . Induction by antigen of intrathymic apoptosis of CD4+CD8+TCRlo thymocytes in vivo. Science 1990; 250: 1720–1723.
Robertson JM, Jensen PE, Evavold BD . DO11.10 and OT-II T cells recognize a C-terminal ovalbumin 323-339 epitope. J Immunol 2000; 164: 4706–4712.
Bouillet P, Purton JF, Godfrey DI, Zhang LC, Coultas L, Puthalakath H et al. BH3-only Bcl-2 family member Bim is required for apoptosis of autoreactive thymocytes. Nature 2002; 415: 922–926.
Herman A, Kappler JW, Marrack P, Pullen AM . Superantigens: mechanism of T-cell stimulation and role in immune responses. Annu Rev Immunol 1991; 9: 745–772.
Gao JX, Zhang H, Bai XF, Wen J, Zheng X, Liu J et al. Perinatal blockade of B7-1 and B7-2 inhibits clonal deletion of highly pathogenic autoreactive T cells. J Exp Med 2002; 195: 959–971.
Theofilopoulos AN, Dixon FJ . Murine models of systemic lupus erythmatoses. Adv Immunol 1985; 37: 269–390.
Toyama H, Okada S, Hatano M, Takahashi Y, Takeda N, Ichii H et al. Memory B cells without somatic hypermutation are generated from Bcl6-deficient B cells. Immunity 2002; 17: 329–339.
Cannons JL, Yu LJ, Hill B, Mijares LA, Dombroski D, Nichols KE et al. SAP regulates T(H)2 differentiation and PKC-theta-mediated activation of NF-kappaB1. Immunity 2004; 21: 693–706.
Zhang J, Bárdos T, Shao Q, Tschopp J, Mikecz K, Glant TT et al. IL-4 potentiates activated T cell apoptosis via an IL-2-dependent mechanism. J Immunol 2003; 170: 3495–3503.
Wu W, Rinaldi L, Fortner KA, Russell JQ, Tschopp J, Irvin C et al. Cellular FLIP long form-transgenic mice manifest a Th2 cytokine bias and enhanced allergic airway inflammation. J Immunol 2004; 172: 4724–4732.
Tseveleki V, Bauer J, Taoufik E, Ruan C, Leondiadis L, Haralambous S et al. Cellular FLIP (long isoform) overexpression in T cells drives Th2 effector responses and promotes immunoregulation in experimental autoimmune encephalomyelitis. J Immunol 2004; 173: 6619–6626.
Sommers CL, Park CS, Lee J, Feng C, Fuller CL, Grinberg A et al. A LAT mutation that inhibits T cell development yet induces lymphoproliferation. Science 2002; 296: 2040–2043.
Sakaguchi N, Takahashi T, Hata H, Nomura T, Tagami T, Yamazaki S et al. Altered thymic T-cell selection due to a mutation of the ZAP-70 gene causes autoimmune arthritis in mice. Nature 2003; 426: 454–460.
Koretzky GA, Abtahian F, Silverman MA . SLP76 and SLP65: complex regulation of signalling in lymphocytes and beyond. Nat Rev Immunol 2006; 6: 67–78.
Salojin KV, Zhang J, Meagher C, Delovitch TL . ZAP-70 is essential for the T cell antigen receptor-induced plasma membrane targeting of SOS and Vav in T cells. J Biol Chem 2000; 275: 5966–5975.
Mora AL, Stanley S, Armistead W, Chan AC, Boothby M . Inefficient ZAP-70 phosphorylation and decreased thymic selection in vivo result from inhibition of NF-kappaB/Rel. J Immunol 2001; 167: 5628–5635.
McNeil LK, Starr TK, Hogquist KA . A requirement for sustained ERK signaling during thymocyte positive selection in vivo. Proc Natl Acad Sci USA 2005; 102: 13574–13579.
Mariathasan S, Ho SSN, Zakarian A, Ohashi PS . Degree of ERK activation influences both positive and negative thymocyte selection. Eur J Immunol 2000; 30: 1060–1068.
Sugawara T, Moriguchi T, Nishida E, Takahama Y . Differential roles of ERK and p38 MAP kinase pathways in positive and negative selection of T lymphocytes. Immunity 1998; 9: 565–574.
Hutcheson J, Scatizzi JC, Siddiqui AM, Haines III GK, Wu T, Li QZ et al. Combined deficiency of proapoptotic regulators Bim and Fas results in the early onset of systemic autoimmunity. Immunity 2008; 28: 206–217.
Xu L, Zhang L, Yi Y, Kang HK, Datta SK . Human lupus T cells resist inactivation and escape death by upregulating COX-2. Nat Med 2004; 10: 411–415.
Kretz-Rommel A, Rubin RL . Disruption of positive selection of thymocytes causes autoimmunity. Nat Med 2000; 6: 298–305.
Werlen G, Hausmann B, Naeher D, Palmer E . Signaling life and death in the thymus: timing is everything. Science 2003; 299: 1859–1863.
Starr TK, Jameson SC, Hogquist KA . Positive and negative selection of T cells. Annu Rev Immunol 2003; 21: 139–176.
Su H, Bidere N, Zheng L, Cubre A, Sakai K, Dale J et al. Requirement for caspase-8 in NF-kappaB activation by antigen receptor. Science 2005; 307: 1465–1468.
Misra RS, Russell JQ, Koenig A, Hinshaw-Makepeace JA, Wen R, Wang D et al. Caspase-8 and c-FLIPL associate in lipid rafts with NF-kappaB adaptors during T cell activation. J Biol Chem 2007; 282: 19365–19374.
Salmena L, Lemmers B, Hakem A, Matysiak-Zablocki E, Murakami K, Au PYB et al. Essential role for caspase 8 in T-cell homeostasis and T-cell-mediated immunity. Genes Dev 2003; 17: 883–895.
Thome M . CARMA1, BCL-10 and MALT1 in lymphocyte development and activation. Nat Rev Immunol 2004; 4: 348–359.
Kataoka T, Budd RC, Holler N, Thome M, Martinon F, Irmler M et al. The caspase-8 inhibitor FLIP promotes activation of NF-kB and Erk signaling pathways. Curr Biol 2000; 10: 640–648.
van Parijs L, Refaeli Y, Abbas AK, Baltimore D . Autoimmunity as a consequence of retrovirus-mediated expression of c-FLIP in lymphocytes. Immunity 1999; 11: 763–770.
Bolland S, Ravetch JV . Spontaneous autoimmune disease in FcγRIIB-deficient mice results from strain-specific epistasis. Immunity 2000; 13: 277–285.
Layer K, Lin G, Nencioni A, Hu W, Schmucker A, Antov AN et al. Autoimmunity as the consequence of a spontaneous mutation in Rasgrp1. Immunity 2003; 19: 243–255.
Salojin K, Zhang J, Cameron M, Gill B, Arreaza G, Ochi A et al. Impaired plasma membrane targeting of Grb2-murine son of sevenless (mSOS) complex and differential activation of the Fyn-T cell receptor (TCR)-zeta-Cbl pathway mediates T cell hyporesponsiveness in autoimmune nonobese diabetic mice. J Exp Med 1997; 186: 887–897.
Acknowledgements
We thank Drs. Marcus Peter and Yang-Xin Fu for critical reading of the paper and discussion. This work is in part supported by a novel research grant from the Lupus Research Institute (to JZ) and a start-up fund from the University of Chicago. Additional support was provided by grants from the National Institutes of Health (NIH) (R01 AR049775 to JZ and DK055357 to RJQ) and from the American Heart Association (09GRNT2010084 to JZ). JZ was supported by a NIH independent scientist award (K02 AR 049047), and is an American Lung Association Career Investigator.
Author information
Authors and Affiliations
Corresponding author
Additional information
Edited by JP Medema
Supplementary Information accompanies the paper on Cell Death and Differentiation website (http://www.nature.com/cdd)
Rights and permissions
About this article
Cite this article
Qiao, G., Li, Z., Minto, A. et al. Altered thymic selection by overexpressing cellular FLICE inhibitory protein in T cells causes lupus-like syndrome in a BALB/c but not C57BL/6 strain. Cell Death Differ 17, 522–533 (2010). https://doi.org/10.1038/cdd.2009.143
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/cdd.2009.143
Keywords
This article is cited by
-
Effects of the C57BL/6 strain background on tauopathy progression in the rTg4510 mouse model
Molecular Neurodegeneration (2014)
-
Constitutive expression of murine c-FLIPR causes autoimmunity in aged mice
Cell Death & Disease (2014)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}