
Abstract
Aim:
Paeoniflorin from Chinese herb Paeoniae Radix has been shown to ameliorate middle cerebral artery occlusion-induced ischemia in rats. The aim of this study was to investigate the mechanisms underlying the neuroprotective action of PF in cultured rat cortical neurons.
Methods:
Primary cultured cortical neurons of rats were subjected to oxygen-glucose deprivation and reoxygenation (OGD/R) insult. Cell survival was determined using MTT assay. HEK293 cells stably transfected with A1R (HEK293/A1R) were used for detailed analysis. Phosphorylation of the signaling proteins was evaluated by Western blot or immunoprecipitation. Receptor interactions were identified using co-immunoprecipitation and immunofluorescence staining.
Results:
Paeoniflorin (10 nmol/L to 1 μmol/L) increased the survival of neurons subjected to OGD/R. Furthermore, paeoniflorin increased the phosphorylation of Akt and ERK1/2 in these neurons. These effects were blocked by PI3K inhibitor wortmannin or MEK inhibitor U0126. Paeoniflorin also increased the phosphorylation of Akt and ERK1/2 in HEK293/A1R cells. Both A1R antagonist DPCPX and EGFR inhibitor AG1478 not only blocked paeoniflorin-induced phosphorylation of ERK1/2 and Akt in HEK293/A1R cells, but also paeoniflorin-increased survival of neurons subjected to OGD/R. In addition, paeoniflorin increased the phosphorylation of Src kinase and activation of MMP-2 in HEK293/A1R cells. Both Src inhibitor PP2 and MMP-2/MMP-9 inhibitor BiPs not only blocked paeoniflorin-induced phosphorylation of ERK1/2 (and Akt) in HEK293/A1R cells, but also paeoniflorin-increased survival of neurons subjected to OGD/R.
Conclusion:
Paeoniflorin promotes the survival of cultured cortical neurons by increasing Akt and ERK1/2 phosphorylation via A1R-mediated transactivation of EGFR.
Similar content being viewed by others


Introduction
Paeoniflorin (PF) is the main active component isolated from the herbal medicine Paeoniae Radix1. PF has many biological activities and has been shown to be anti-inflammatory2,3,4, analgesic5, and an antioxidant6,7. Furthermore, PF is a potential neuroprotective agent. An increasing number of studies have shown that natural compounds have great potential for preventing or treating cerebral ischemia and reperfusion injuries8. Our previous studies have demonstrated that PF pretreatment could decrease the infarct volumes of rats suffering from a middle cerebral artery occlusion (MCAO)9. Such neuroprotective effects might be mediated by adenosine A1 receptor (A1R) activation, because the A1R antagonist 8-cyclopentyl-1,3-dipropylxanthine (DPCPX) could suppress the protective effect of PF.
A1R is a subtype of adenosine receptors that are highly expressed in the brain10 and is coupled to the Gi protein. When ischemic injury occurs, extracellular adenosine increases and activates A1R, which works as a neuromodulator to inhibit excitotoxic neuronal injury11 . This induces presynaptic A1Rs to decrease the release of glutamate and relieves Ca2+ overloading at postsynaptic A1Rs12. A1R activation therefore prevents the damage to neurons caused by overstimulation with excitatory transmitters and promotes their survival after ischemic injury and reperfusion12. Oxygen-glucose deprivation followed by reoxygenation (OGD/R) mimics an ischemia and reperfusion injury and is often used as a model of ischemic injury in neurons and other cells.
The PI3K/Akt and MAPK/ERK signaling pathways are also known as the reperfusion and ischemia salvage kinase (RISK) pathways owing to their involvement in ameliorating ischemia and reperfusion injuries. The activation of RISK pathways is associated with an inhibition of apoptosis13,14,15. Several receptor tyrosine kinases (RTKs) activate both the Akt and ERK1/2 signaling pathways. Many G protein-coupled receptors (GPCRs) transactivate RTKs, resulting in Akt and ERK1/2 activation, which results in cell protection and survival16. Epidermal growth factor receptor (EGFR) is a typical RTK expressed in the membrane of many kinds of cells. EGFR plays a major role in cell growth, proliferation, survival, differentiation, and so on. Several phosphorylation sites in the C-terminal domain of EGFR, when activated, further stimulate the activation of downstream signaling pathways, including MAPK, Akt, and JNK17. The selective A1R agonist cyclopentyladenosine (CPA) promotes neuronal survival after an OGD/R injury, most likely through PI3K/Akt signaling induced by A1R-mediated transactivation of EGFR18. Consistent with its role in neuronal protection, it has also been found that A1R-mediated transactivation of EGFR protects cardiac cells against ischemia and reperfusion19.
The molecular mechanism of the anti-ischemic effect of PF remains unclear. We hypothesize that PF works as an agent that preconditions the brain against ischemic injury by activating the MAPK/ERK and PI3K/Akt signaling pathways. Preconditioning with PF could protect cells from subsequent injury and death through the activation of A1R. In this research, PF was applied to primary cultured cortical neurons and HEK293 cells stably transfected with A1R (HEK293/A1R) to explore its mechanism of neuroprotection.
Materials and methods
Materials
PF (purity, 99.42%20; structure shown in Figure 1) was isolated at the Shanghai Institute of Materia Medica. CPA, DPCPX, EGF, forskolin, rolipram, and anti-β-actin were purchased from Sigma-Aldrich (St Louis, MO, USA). AG1478, (2R)-[(4-biphenylylsulfonyl)amino]-N-hydroxy-3-phenylpropionamid (BiPs), wortmannin, and U0126 were purchased from Calbiochem (San Diego, CA, USA). PP2 and PP3 were purchased from Tocris (Bristol, UK). The following primary antibodies were purchased from Cell Signaling Technology: anti-phospho-ERK1/2 (Thr202/Tyr204), anti-ERK1/2 (Thr202/Tyr204), anti-phospho-Akt (Ser473), anti-Akt, anti-phospho-Src (Tyr416), and anti-phospho-EGFR (Tyr1068, Tyr845). Anti-EGFR (mouse IgG) was purchased from Upstate Biotechnology (Lake Placid, NY, USA). Anti-A1R was purchased from Affinity BioReagents (Golden, CO, USA). Normal immunoglobulin G (IgG), Protein A/G PLUS-Agarose immunoprecipitation reagent, anti-Src, and the horseradish peroxidase (HRP)-conjugated secondary antibodies used for Western blots were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Alexa Fluor 488-conjugated goat anti-mouse IgG and Alexa Fluor 555-conjugated goat anti-rabbit IgG were purchased from Invitrogen/Life Technologies (Grand Island, NY, USA). The homogeneous time-resolved fluorescence (HTRF) cAMP kit was purchased from Cisbio (Cisbio Bioassays, France). MTT was purchased from Bio Basic (Markham Ontario, Canada). DMEM, Neurobasal medium, B27 cell culture supplement, and G418 were purchased from Life Technologies (Grand Island, NY, USA).

Chemical structure of PF.
Primary culture of cortical neurons
All animal procedures were performed following a protocol approved by the Institutional Animal Care and Use Committee at the Shanghai Institute of Materia Medica, Chinese Academy of Sciences, permit number (2011-03-FLY-06). Cortical neurons were cultured from Sprague-Dawley rats at embryonic day 18 (E18) using a modified method that has been previously described18. Fetal brains were removed under sterile conditions. Isolated cerebral cortices were digested with 0.125% Trypsin, and cells were cultured on Poly-D-Lysine-coated plates in DMEM with 10% FBS. After 4 h, the neurons were placed in growth media (Neurobasal medium with B27 supplement) and cultured at 37 °C with 5% CO2 humidified air.
Oxygen-glucose deprivation/reoxygenation (OGD/R) and MTT assay
OGD was carried out on 7–8 DIV (days in vitro) cultures. Briefly, neurons were pretreated with PF in B27/Neurobasal media for 2 h. The cells were washed once and maintained in glucose-free DMEM without serum at 37 °C in a humidified, sealed chamber that had been flushed with 95% N2/5% CO2 for 5 min. At the end of the 4-h OGD period, the cultures were returned to growth media for another 24 h. In some assays, wortmannin (100 nmol/L), U0126 (1 μmol/L), AG1478 (1 μmol/L), DPCPX (1 μmol/L), PP2 (5 μmol/L) or BiPs (10 μmol/L) was added for 30 min before the PF pretreatment. The test compounds were always included in the OGD solution during the OGD/R period. MTT solution (0.5 mg/mL) was added to each well, and cells were maintained in growth media for an additional 4 h. After solubilization with DMSO, the absorbance of the formazan dye was measured using a microplate reader (NovoStar, BMG) at 595 nm. In each experiment, cultures exposed to OGD/R were compared with normoxic controls, which were maintained under standard incubation conditions.
HEK293/A1R cell culture
A derivative of the HEK293 cell line that stably expresses the human A1R (HEK293/A1R cells) was established previously and maintained in our laboratory18,21. Cells were cultured in DMEM supplemented with 10% FBS and 500 μg/mL G418, and cultured at 37 °C with 5% CO2 humidified air.
The plasmids of mutant A1Rs used for transient transfections were constructed previously in our laboratory22. After transfection, the cells were cultured in the same conditions as the HEK293/A1R cells.
Western blot analysis
The cells were lysed in RIPA buffer [20 mmol/L Tris (pH 7.4), 150 mmol/L NaCl, 1% NP-40, 0.5% sodium deoxycholate, and protease inhibitor cocktail] and centrifuged at 15 000×g for 10 min at 4 °C. The protein concentration was determined using a Bradford assay. The samples were electrophoresed on an SDS-PAGE gel and transferred onto a PVDF membrane. The membrane was blocked with 5% non-fat milk in Tris-buffered saline and 0.1% Tween-20 for 1 h and subsequently incubated overnight at 4 °C with primary antibody diluted in Tris-buffered saline with 5% BSA and 0.1% (v/v) Tween-20. After incubation with the appropriate HRP-conjugated secondary antibody for 1 h, the blots were developed by chemiluminescence and exposed to X-ray film. If necessary, the blot was stripped and reprobed with additional antibodies.
Co-immunoprecipitation (Co-IP) assay
HEK293/A1R cells were serum-starved overnight at 37 °C before treatment. The cell lysates were prepared by incubating the cells in lysis buffer on ice. Clarified lysates were immunoprecipitated with an anti-A1R antibody or an anti-EGFR antibody overnight at 4 °C and then incubated with Protein A/G-PLUS agarose beads. Equivalent amounts of protein were analyzed for each condition. Normal IgG was used as a negative control. The immune complexes were boiled in an SDS sample buffer and loaded on SDS-PAGE gels for immunoblot analysis.
Immunofluorescence staining
HEK293/A1R cells were fixed in 4% paraformaldehyde for 30 min at 4 °C. The cells were blocked for 1 h in blocking buffer (5% normal goat serum, 1% BSA, 0.1% Triton X-100, and 0.1% sodium azide in PBS), then incubated with anti-A1R and anti-EGFR antibodies diluted in blocking buffer at 4 °C overnight. The cells were washed three times with PBS and then incubated with Alexa Fluor 488-conjugated goat anti-rabbit IgG and Alexa Fluor 555-conjugated goat anti-mouse IgG for 1 h at room temperature in the dark. Images were acquired with a confocal microscope (Olympus, Japan). To determine the co-localization level of A1R and EGFR, Pearson's and Manders' coefficients were calculated using FV10-ASW2.1 software (Olympus) according to the previously reported analysis methods23. Briefly, Pearson's correlation coefficient describes the correlation of the intensity distribution between the channels, ranging from −1.0 to 1.0, where 0 indicates no significant correlation. The overlap coefficient according to Manders' indicates an actual overlap of the signals ranging from 0 to 1.0, while 0.5 means that 50% of both selected channels co-localize.
cAMP assay
Following the instructions of the HTRF cAMP test kit (Cisbio, #62AM4PEB), HEK293 cells and HEK293/A1R cells were harvested and resuspended in DMEM with rolipram (30 μmol/L). The cells were counted and pretreated with PF (100 nmol/L) or CPA (100 nmol/L) for 30 min and then forskolin for an additional 30 min. The cells were lysed and intracellular cAMP was detected using a competitive immunoassay. The cAMP concentration was calculated according to the fluorescence ratio (665 nm/620 nm).
Docking experiments
Homology modeling was used to determine the structure of A1R. The structure of A2AR (PDB code: 3EML24) was used as a template, and Modeler 9.1025 was used to build the structure. Lysozyme was removed and the missing residues in ICL2 were modeled by the loop-model module in Modeler. The default values were used for all other settings. Ten models were generated, and the best model was chosen by experience.
Docking experiments were performed using the Autodock 4.2 suite26. A docking box covering the entire extracellular domain and the orthodox binding site was used. A genetic algorithm was used for the conformation search, and a Lamarckian genetic algorithm was adopted for conformation evaluation. The 50 best energy-preferred conformations were noted.
Statistical analysis
Data were obtained from at least three independent experiments and presented as the mean±SEM. Statistical analysis of the results was carried out using one-way ANOVA followed by the Dunnett test, using Graphpad Prism software (La Jolla, CA, USA). P values <0.05 were considered significant.
Results
PF protects cortical neurons from OGD/R injury
PF (10 nmol/L, 100 nmol/L, or 1 μmol/L) was added to cortical neurons 2 h before OGD until the end of the reoxygenation period (24 h). As expected, compared with the normoxia group, exposure to OGD/R caused an impairment in the cortical neurons that appeared to damage their somas and neurites, as observed under a phase contrast microscope. Compared to the OGD/R neurons, neurons pretreated with PF were more likely to maintain their morphology (Figure 2A). PF improved neuronal viability as detected using an MTT assay. The neuroprotective effects of PF were significant at concentrations above 100 nmol/L (Figure 2B).

The neuroprotective effects of PF on primary cultured cortical neurons exposed to OGD/R injury. Primary cultured cortical neurons were treated with PF (10 nmol/L, 100 nmol/L, or 1 μmol/L) for 2 h and then exposed to OGD injury for 4 h, as previously described. The morphology of the neurons was imaged using phase contrast microscopy 24 h later (A-E) and neuronal viability was determined using the MTT assay (F). Data are presented as mean±SEM. cP<0.01 vs OGD/R. eP<0.05 vs normoxia. n=3. The scale bar represents 200 μm.
PF activates the Akt and ERK1/2 signaling pathways
Activation of the Akt and ERK1/2 signaling pathways may ameliorate injury in neurons exposed to ischemia and reperfusion. After 30 min of reoxygenation, Akt (Ser473) and ERK1/2 (Thr202/Tyr204) phosphorylation levels were evaluated using Western blot analysis. Neurons treated with PF (100 nmol/L) showed increased Akt and ERK1/2 phosphorylation levels compared to non-PF-treated OGD/R neurons (Figure 3A and 3B), suggesting that PF may protect neurons from ischemic injury by activating Akt and ERK1/2.

The neuroprotective effect of PF is mediated by the activation of both PI3K/Akt and MAPK/ERK in cortical neurons exposed to OGD/R injury. Cortical neurons were treated with PF (10 nmol/L, 100 nmol/L, or 1 μmol/L) for 2 h and then exposed to OGD for 4 h and reoxygenation for 30 min. The cell lysates were analyzed for Akt and ERK1/2 phosphorylation by Western blot. Neurons maintained at normoxia were used as a control (A and B). Data are presented as the mean±SEM. bP<0.05, cP<0.01 vs control, n=3. Cortical neurons were pretreated with wortmannin (100 nmol/L) or U0126 (1 μmol/L) for 30 min before PF treatment for 2 h. The neurons were then exposed to OGD/R injury as above. Neuronal survival was assessed using the MTT assay (C). Data are presented as the mean±SEM. eP<0.05, fP<0.01 vs OGD/R, n=8.
To identify the role of Akt and ERK1/2 in the neuroprotective effect of PF, inhibitors of PI3K and MEK were used to inhibit the activation of Akt and ERK1/2, respectively. Pretreatment with wortmannin (100 nmol/L) and U0126 (1 μmol/L) inhibited neuronal survival after OGD/R, an effect that was partially reversed by PF (100 nmol/L) (Figure 3C). Although pretreatment with wortmannin or U0126 slightly modified the effect of PF, it is likely that the activation of the PI3K/Akt and MAPK/ERK signaling pathways worked in concert for the total neuroprotective effect stimulated by PF. These results indicate that the PI3K/Akt and MAPK/ERK signaling pathways are simultaneously essential for the neuroprotective effect of PF. Next, PF (10 nmol/L, 100 nmol/L, or 1 μmol/L) was added to primary cultured neurons for a short period of 15 min. PF (100 nmol/L) induced both Akt and ERK1/2 phosphorylation (Figure 4A and 4B). Neurons were then treated with PF (100 nmol/L) for varying lengths of time. Significant phosphorylation of Akt and ERK1/2 was observed at 15 min and lasted for 1 h (Figure 4C and 4D). These results indicate that PF can ameliorate ischemic injury by activating survival signaling pathways, including the PI3K and ERK1/2 pathways.

The effect of PF on the activation of Akt and ERK1/2 in primary cultured cortical neurons. Primary cultured neurons were starved of the B27 supplement overnight and then incubated with PF (10 nmol/L, 100 nmol/L, or 1 μmol/L) for 15 min. The phosphorylation of Akt and ERK1/2 was detected by Western blot (A and B). A time course (5, 15, 30, 60, and 120 min) of the effect of PF (100 nmol/L) on Akt and ERK1/2 phosphorylation in cortical neurons (C and D). Data are presented as the mean±SEM. bP<0.05, cP<0.01 vs control. n=3.
Akt and ERK1/2 activation by PF is dependent on A1R
In has been reported that pretreatment with the A1R-selective antagonist DPCPX can block PF's neuroprotective effects in the MCAO rat model of ischemic stroke9. Molecule dockings were therefore conducted on PF, CPA (an A1R agonist), and DPCPX (Supplementary Figure S1). The molecular docking revealed that both DPCPX and CPA bind to A1R at the orthodox binding site, which is located within the transmembrane helical bundles. PF, conversely, docks at an extracellular site. Out of the 50 best energy-preferred conformations, 32 were docked to an extracellular site, while 18 were docked to a site above the orthodox site. These results suggest that PF binds to A1R differently from selective agonists.
To further investigate the detailed molecular mechanism of PF neuroprotection, A1R activation by PF in stably transfected HEK293/A1R cells was examined. PF-induced phosphorylation of Akt and ERK1/2 was apparent at 5 min (Figure 5A and 5B) and lasted for approximately 1 h (Figure 5C and 5D), as in the cortical neurons. These results also confirmed that the selective A1R agonist CPA could stimulate Akt and ERK1/2 phosphorylation in HEK293/A1R cells and suggest that the signaling pathways induced by PF might also be dependent on A1R.

The effect of PF on the activation of Akt and ERK1/2 in HEK293/A1R cells. HEK293/A1R cells were serum-starved overnight and then treated with PF (10 nmol/L, 100 nmol/L, or 1 μmol/L) and CPA (100 nmol/L) for 5 min. The phosphorylation of Akt and ERK1/2 was detected by Western blot (A and B). Serum-starved HEK293/A1R cells were treated with PF (100 nmol/L) for 5, 15, 30, or 60 min. The cell lysates were collected and analyzed for phosphorylation of Akt and ERK1/2 by Western blot (C and D). Data are presented as the mean±SEM. bP<0.05, cP<0.01 vs control. n=3.
To determine the role of A1R in Akt and ERK1/2 phosphorylation in response to PF, several experiments were designed. First, cAMP levels were evaluated, because A1R activates Gi protein signaling, which in turn induces cAMP downregulation. PF could depress the increase in cAMP levels induced by forskolin (1 μmol/L) in HEK293/A1R cells but not in control HEK293 cells. These results indicate that PF can target and activate A1R (Figure 6A). PF did not induce Akt and ERK1/2 phosphorylation in control HEK293 cells without A1R (Figure 6B). Furthermore, the A1R antagonist DPCPX could inhibit the Akt and ERK1/2 phosphorylation induced by PF, in agreement with results in MCAO rats27 (Figure 6C and 6D). Next, several A1R mutants were transiently transfected into HEK293 cells. The mutants include L65T (Leu-65 to Thr-65), I69S (Ile-69 to Ser-69), I69T (Ile-69 to Thr-69), I69V (Ile-69 to Val-69), and F185L (Phe-185 to Leu-185)22. The mutated residues were chosen based on their possible involvement in PF binding to A1R, particularly residues with hydrophobic amino acids. Several mutants resulted in a reduced phosphorylation of Akt and ERK1/2 compared to wild-type A1R (Figure 6E and 6F). These results suggest that A1R is essential for PF-induced Akt and ERK1/2 phosphorylation.

The activation of Akt and ERK1/2 by PF is mediated by A1R. HEK293/A1R cells and HEK293 cells were pretreated with PF (100 nmol/L) or CPA (100 nmol/L) for 30 min and then incubated with forskolin (1 μmol/L) for 30 min. Cyclic AMP was measured using a cAMP kit. The cAMP levels in forskolin-treated HEK293/A1R and HEK293 cells were normalized to 100%. All other values are shown relative to forskolin-treated cells, for each respective cell type (A). Data are presented as the mean±SEM. bP<0.05 vs forskolin-treated HEK293 cells. fP<0.01 vs forskolin-treated HEK293/A1R cells. n=3. The phosphorylation of Akt and ERK1/2 induced by a 5-min treatment with PF (100 nmol/L), CPA (100 nmol/L), and EGF (5 ng/mL) in HEK293 cells was detected by Western blot (B). HEK293/A1R cells were pretreated with DPCPX (1 μmol/L) for 30 min in the presence or absence of 100 nmol/L PF (C and D). Data are presented as the mean±SEM. bP<0.05, cP<0.01 vs control. eP<0.05 vs PF. n=3. HEK293 cells were transiently transfected with A1R (wild-type or L65T, I69S, I69T, or I69V) or an empty vector, as a negative control. Akt and ERK1/2 phosphorylation induced by 5 min of PF treatment was analyzed by Western blot (E and F). Data are presented as the mean±SEM. bP<0.05, cP<0.01 vs control. n=3.
EGFR is involved in Akt and ERK1/2 phosphorylation in response to PF
It has been demonstrated that the A1R agonist CPA can activate Akt and/or ERK1/2 in a manner that is dependent on EGFR transactivation. This Akt and ERK1/2 signaling protected neurons and cardiac cells against injury from ischemia and reperfusion in vitro18,19.
After treatment of cortical neurons and HEK293/A1R cells with PF for 5 min, EGFR was phosphorylated at Tyr1068, as detected by Western blot (Figure 7A and 7B). It is notable that PF failed to activate Akt and ERK1/2 through EGFR in HEK293 cells but did so only in the HEK293/A1R cells (Figure 6B and 7B).

EGFR transactivation by PF activates Akt and ERK1/2. Cortical neurons were treated with PF (10 nmol/L, 100 nmol/L, or 1 μmol/L) for 15 min and EGFR Tyr1068 phosphorylation was detected by Western blot (A). HEK293/A1R cells were treated with PF (100 nmol/L), CPA (100 nmol/L), or EGF (5 ng/mL) for 5 min and the cell lysates were probed for the phosphorylation of EGFR Tyr1068 (B). HEK293/A1R cells were pretreated with AG1478 (1 μmol/L) for 30 min. The phosphorylation of Akt and ERK1/2 after a 5-min treatment with PF was detected by Western blot (C and D). Data are presented as the mean±SEM. bP<0.05, cP<0.01 vs control. eP<0.05, fP<0.01 vs PF. n=3.
AG1478 (1 μmol/L), a selective inhibitor of the EGFR tyrosine kinase, suppressed Akt and ERK1/2 phosphorylation in response to PF (Figure 7C and 7D). This suggests that EGFR phosphorylation was involved in the activation of the Akt and ERK1/2 signaling pathways as an upstream event. These results imply that the PF-induced transactivation of EGFR was mediated by A1R and resulted in the phosphorylation of Akt and ERK1/2. The phosphorylation of EGFR at Tyr1068 was always associated with MAPK/ERK and PI3K/Akt activation, suggesting that PF could activate EGFR.
PF strengthened the interaction between A1R and EGFR
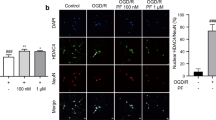
Since PF activation of the Akt and ERK1/2 signaling pathways required A1R and EGFR activation, the interaction between these two receptors after PF treatment was examined next. In an experiment with OGD/R neurons, pretreatment with DPCPX or AG1478 suppressed the neuroprotective effects of PF (Figure 8A). In HEK293/A1R cells, a co-IP assay also indicated that PF could strengthen the interaction between A1R and EGFR (Figure 8B). PF (100 nmol/L) treatment of HEK293/A1R cells for 5 min enhanced the co-localization of A1R and EGFR on the cell surface, as observed using confocal microscopy (Figure 8C). The increase in their co-localization coefficients indicates that this interaction facilitates downstream signal transduction (Figure 8D and 8E).

A1R and EGFR are essential for the effects of PF. Cortical neurons were pretreated with DPCPX (1 μmol/L) or AG1478 (1 μmol/L) for 30 min and then treated with PF for 2 h. Afterwards, the neurons were exposed to OGD/R and survival was evaluated using the MTT assay (A). Data are presented as the mean±SEM. cP<0.01 vs OGD/R. fP<0.01 vs OGD/R+PF, n=8. HEK293/A1R cells were treated with PF (100 nmol/L) and EGF (5 ng/mL) for 5 min. The cell lysates were immunoprecipitated with an anti-A1R antibody and immunoblotted with antibodies against A1R and EGFR (B). HEK293/A1R cells were treated with PF (10 nmol/L, 100 nmol/L, or 1 μmol/L) for 5 min. Confocal microscopy showed that A1R and EGFR co-localized at the cell membrane, as revealed by immunofluorescence staining. The scale bar represents 50 μm (C). The extent of co-localization was evaluated by correlation coefficients (D and E). Data are presented as the mean±SEM. bP<0.05, cP<0.01 vs control. n=30.
Both Src and MMP-2 are involved in signaling pathways induced by PF
Two patterns of GPCR transactivation of EGFR have been reported. One mechanism is the direct activation of the EGFR tyrosine kinase at the C terminus (in particular, at Tyr845 and Tyr1101) by Src kinase, and the other mechanism is the cleavage of endogenous ligands of EGFR, including heparin-binding EGF-like growth factor (HB-EGF) and transforming growth factor (TGF)-α, by matrix metalloproteases (MMPs)28.
In our study, Src tyrosine kinase was phosphorylated at Tyr416 in response to PF treatment in HEK293/A1R cells. The pretreatment of cells with PP2 (5 μmol/L), a selective inhibitor of Src, but not PP3 (an analogue of PP2 that serves as negative control) inhibited the phosphorylation of Src kinase (Figure 9A). Furthermore, EGFR was also phosphorylated at Tyr845 by the PF treatment (Figure 9B). The pretreatment with PP2 also lowered the phosphorylation levels of Akt and ERK1/2 (Figure 9C and 9D), which indicates that Src is involved in the signaling pathways induced by PF. These data suggest that A1R activates Src, which then activates EGFR, resulting in the observed downstream signaling.

Src is involved in the activation of both Akt and ERK1/2 by PF. HEK293/A1R cells were pretreated with PP2 (5 μmol/L) or PP3 (5 μmol/L) for 30 min and then treated with PF for 5 min. The cell lysates were analyzed for phosphorylation of Src at Tyr416 (A). HEK293/A1R cells were treated with PF and EGF for 5 min. Phosphorylation of EGFR Tyr845 was detected using immunoprecipitation with an anti-EGFR antibody (B). HEK293/A1R cells were pretreated with PP2 for 30 min and then treated with PF for 5 min. The phosphorylation of Akt and ERK1/2 was detected by Western blot (C and D). Data are presented as the mean±SEM. bP<0.05 vs control. eP<0.05, fP<0.01 vs PF. n=3.
To investigate the other potential mechanism of EGFR transactivation, MMP activation was evaluated by Western blot. BiPs, an inhibitor of MMP-2/MMP-9, inhibited the phosphorylation of ERK1/2 in response to PF in HEK293/A1R cells (Figure 10A). MMP-2/MMP-9 cleaves pro-HB-EGF into free HB-EGF that could activate the EGFR at the cell membrane. PF increased the generation of active MMP-2 and BiPs inhibited this effect (Figure 10B). MMP-9 did not change after PF treatment (data not show). These data indicate that PF increased the amount of HB-EGF and induced EGFR autophosphorylation.

MMP-2 is involved in the activation of ERK1/2. HEK293/A1R cells were pretreated with BiPs (10 μmol/L) for 30 min. The cells were then treated with PF (100 nmol/L) for 5 min, and ERK1/2 phosphorylation (A), pro-MMP-2, and active MMP-2 (B) were detected by Western blot. Data are presented as the mean±SEM. bP<0.05 vs control. eP<0.05, fP<0.01 vs PF. n=3. Cortical neurons were pretreated with PP2 (5 μmol/L) and BiPs (10 μmol/L) for 30 min prior to the treatment with PF for 2 h and then exposed to OGD/R injury as described above. Neuronal viability was analyzed using the MTT assay (C). Data are presented as the mean±SEM. cP<0.01 vs OGD/R. fP<0.01 vs OGD/R+PF. n=8.
Because both Src and MMP-2 were involved in the activation of the Akt and/or ERK signaling pathways by PF, they might also be involved in the neuroprotective effect of PF. PP2 (5 μmol/L) and BiPs (10 μmol/L) pretreatment reversed the protective effect of PF on neuronal survival following an ischemic OGD/R injury (Figure 10C). These results suggest that Src and MMP-2 are both involved in the neuroprotective effect of PF via the activation of EGFR.
Discussion
While stroke is becoming a worldwide financial burden and health challenge, the cellular and molecular mechanisms of brain damage remain unclear. No neuroprotective agents exist to protect against an ischemic cerebrovascular injury29. PF was isolated from one herbal medicine that has been demonstrated to be neuroprotective in the MCAO rat model of ischemic stroke, although the underlying mechanism is unknown.
In our studies, PF preconditioning exerted a neuroprotective effect against ischemia and reoxygenation via the activation of the Akt and ERK1/2 survival signaling pathways. It has been known that the phosphorylation of Akt and ERK1/2 inhibits apoptosis and can rescue cells from injury following ischemia and reperfusion15. In this study, the treatment of cortical neurons with PF induced Akt and ERK1/2 phosphorylation during the reoxygenation period and promoted their survival. PI3K/Akt pathway activation is found in many studies and plays a key role against ischemia/reperfusion injuries30,31. Activation of the ERK cascade has also been shown to promote the restoration of learning and memory after cerebral ischemia8.
The activation of Akt and ERK1/2 by PF was mediated by the transactivation of EGFR through A1R. Akt and ERK activation is likely downstream of the EGFR activation stimulated by PF; thus, they play a common role in the neuroprotective effect exerted by PF against OGD/R injury. PF could not induce Akt or ERK1/2 phosphorylation in the absence of A1R, which demonstrates that PF activates these two pathways via transactivation. This might explain why inhibiting A1R or EGFR reduced neuronal survival compared with PF treatment alone.
Although PF is not a typical A1R agonist, several studies have shown that PF targets the A1R9,27,32,33. In our study, PF was found to activate A1R and reduce cAMP levels. Some A1R mutants are thought to be related to the ligand-receptor binding22 that would affect the signaling induced by PF especially L65T and I69S in which hydrophobic residues are replaced by hydrophilic ones located in the second transmembrane region of A1R. It will be interesting to explore the relationship between the structure and function of PF binding to A1R.
To further understand the binding behavior of PF, the molecular docking of PF was compared to that of CPA and DPCPX (Supplementary Figure S1). GPCRs have been known to have allosteric binding sites within the extracellular loops, and the docking results suggested that PF would bind to either an allosteric site in an extracellular loop or a site in the extracellular region. These binding site differences may explain the difference in the biological activities of PF and DPCPX.
It has been shown that A1R transactivation of EGFR exerts protective effects against ischemia and reperfusion18,19. PF acted at the surface of HEK293/A1R cells to promote an interaction between A1R and EGFR, which increased signal transduction from the two receptors. The phosphorylation of EGFR at Tyr1068 and Tyr845 induced by PF is thought to be involved in MAPK and Src activation, respectively. Both Src tyrosine kinase and MMP-2 were involved in the transactivation of EGFR after PF treatment. Src seems to play a greater role than MMP-2 in HEK293/A1R cells, because inhibition of MMP-2 with BiPs only inhibited ERK1/2 phosphorylation and not Akt phosphorylation. However, the inhibition of Src or MMP in neurons abolished the neuroprotective effects significantly, indicating that differences in the mechanisms of PF could exist in different cell types.
The literature of cerebral ischemia indicates that MMPs play multiple roles34. MMP-9 exhibits high levels of activity during ischemia, especially in the acute phase, while MMP-2 has been found to be elevated in the delayed phase and showed contrary performance which remains to disclosure35. Here, we found that PF could activate MMP-2 and subsequently EGFR in HEK293/A1R cells, an effect that might occur via sheared HB-EGF release. However, it is only inferred that an increase in MMP-2 could contribute to the neuroprotective effect of PF, as neurons are different from HEK293/A1R cells.
The phenomenon of GPCR transactivation of RTKs is universal and may represent a novel target for drug development36. Several studies have found that GPCRs transactivate RTKs that exert neurotrophic effects, such as the insulin growth factor (IGF)-1 receptor, EGFR, and TrkA/B37,38,39. PF has been studied for decades and has many effects in vivo and in vitro, several of which are independent of A1R3,20. We explored the mechanism of action of PF in HEK293/A1R cells and cortical neurons because both A1R and EGFR are expressed in these two cell types. PF functioned as a neurotrophic factor to protect cortical neurons from OGD/R injury. These results explain the anti-ischemic effect of PF in MCAO rats, because the cortex suffers from ischemia and reperfusion in that model. In addition to the other effects of PF, the neurotrophic-like effects of PF could rescue neurons during ischemic stroke and other cerebrovascular diseases.
In conclusion, PF preconditioned neurons to enhance their survival after ischemic injury. This effect was dependent on Akt and ERK1/2 phosphorylation, which was induced by A1R-mediated transactivation of EGFR. Src and MMP-2 were also involved in the transactivation of EGFR by A1R.
Author contribution
Lin-yin FENG conceived and designed the experiments; Yang YE synthesized the natural product; Min ZHONG performed the experiments and analyzed the data; Wan-ling SONG and Ye-chun XU conducted the computational docking analysis; Min ZHONG and Lin-yin FENG wrote the manuscript.
References
Ikeda N, Fukuda T, Jyo H, Shimada Y, Murakami N . Quality evaluation on Paeoniae radix. I. Quantitative analysis of monoterpene glycosides constituents of Paeoniae radix by means of high performance liquid chromatography. Comparative characterization of the external figures, processing method and the cultivated areas. Yakugaku Zasshi 1996; 116: 138–47.
Zhang LL, Wei W, Wang NP, Wang QT, Chen JY, Chen Y, et al. Paeoniflorin suppresses inflammatory mediator production and regulates G protein-coupled signaling in fibroblast-like synoviocytes of collagen induced arthritic rats. Inflamm Res 2008; 57: 388–95.
Kim ID, Ha BJ . Paeoniflorin protects RAW 264.7 macrophages from LPS-induced cytotoxicity and genotoxicity. Toxicol In Vitro 2009; 23: 1014–9.
Chen T, Guo ZP, Jiao XY, Jia RZ, Zhang YH, Li JY, et al. Peoniflorin suppresses tumor necrosis factor-alpha induced chemokine production in human dermal microvascular endothelial cells by blocking nuclear factor-kappaB and ERK pathway. Arch Dermatol Res 2011; 303: 351–60.
Zhang XJ, Li Z, Leung WM, Liu L, Xu HX, Bian ZX . The analgesic effect of paeoniflorin on neonatal maternal separation-induced visceral hyperalgesia in rats. J Pain 2008; 9: 497–505.
Li CR, Zhou Z, Zhu D, Sun YN, Dai JM, Wang SQ . Protective effect of paeoniflorin on irradiation-induced cell damage involved in modulation of reactive oxygen species and the mitogen-activated protein kinases. Int J Biochem Cell Biol 2007; 39: 426–38.
Chen T, Guo ZP, Jiao XY, Zhang YH, Li JY, Liu HJ . Protective effects of peoniflorin against hydrogen peroxide-induced oxidative stress in human umbilical vein endothelial cells. Can J Physiol Pharmacol 2011; 89: 445–53.
WU PF, ZHANG Z, WANG F, CHEN JG . Natural compounds from traditional medicinal herbs in the treatment of cerebral ischemia/reperfusion injury. Acta Pharmacol Sin 2010; 31: 1523–31.
Liu DZ, Xie KQ, Ji XQ, Ye Y, Jiang CL, Zhu XZ . Neuroprotective effect of paeoniflorin on cerebral ischemic rat by activating adenosine A1receptor in a manner different from its classical agonists. Br J Pharmacol 2005; 146: 604–11.
Dixon AK, Gubitz AK, Sirinathsinghji DJ, Richardson PJ, Freeman TC . Tissue distribution of adenosine receptor mRNAs in the rat. Br J Pharmacol 1996; 118: 1461–8.
Chu S, Xiong W, Zhang D, Soylu H, Sun C, Albensi BC, et al. Regulation of adenosine levels during cerebral ischemia. Acta Pharmacol Sin 2013; 34: 60–6.
Wardas J . Neuroprotective role of adenosine in the CNS. Pol J Pharmacol 2002; 54: 313–26.
Hausenloy DJ, Yellon DM . New directions for protecting the heart against ischaemia-reperfusion injury: targeting the reperfusion injury salvage kinase (RISK)-pathway. Cardiovasc Res 2004; 61: 448–60.
Hausenloy DJ, Tsang A, Yellon DM . The reperfusion injury salvage kinase pathway: a common target for both ischemic preconditioning and postconditioning. Trends Cardiovasc Med 2005; 15: 69–75.
Liu XQ, Sheng R, Qin ZH . The neuroprotective mechanism of brain ischemic preconditioning. Acta Pharmacol Sin 2009; 30: 1071–80.
Shah BH, Catt KJ . GPCR-mediated transactivation of RTKs in the CNS: mechanisms and consequences. Trends Neurosci 2004; 27: 48–53.
Oda K, Matsuoka Y, Funahashi A, Kitano H . A comprehensive pathway map of epidermal growth factor receptor signaling. Mol Syst Biol 2005; 1: 2005.0010.
Xie KQ, Zhang LM, Cao Y, Zhu J, Feng LY . Adenosine A1 receptor-mediated transactivation of the EGF receptor produces a neuroprotective effect on cortical neurons in vitro. Acta Pharmacol Sin 2009; 30: 889–98.
Williams-Pritchard G, Knight M, Hoe LS, Headrick JP, Peart JN . Essential role of EGFR in cardioprotection and signaling responses to A1 adenosine receptors and ischemic preconditioning. Am J Physiol Heart Circ Physiol 2011; 300: H2161–8.
Jin L, Zhang LM, Xie KQ, Ye Y, Feng L . Paeoniflorin suppresses the expression of intercellular adhesion molecule-1 (ICAM-1) in endotoxin-treated human monocytic cells. Br J Pharmacol 2011; 164: 694–703.
Cao Y, Sun WC, Jin L, Xie KQ, Zhu XZ . Activation of adenosine A1 receptor modulates dopamine D1 receptor activity in stably cotransfected human embryonic kidney 293 cells. Eur J Pharmacol 2006; 548: 29–35.
Xie KQ, Cao Y, Zhu XZ . Role of the second transmembrane domain of rat adenosine A1 receptor in ligand-receptor interaction. Biochem Pharmacol 2006; 71: 865–71.
Zinchuk V, Zinchuk O, Okada T . Quantitative colocalization analysis of multicolor confocal immunofluorescence microscopy images: pushing pixels to explore biological phenomena. Acta Histochem Cytochem 2007; 40: 101–11.
Jaakola VP, Griffith MT, Hanson MA, Cherezov V, Chien EY, Lane JR, et al. The 2.6 angstrom crystal structure of a human A2A adenosine receptor bound to an antagonist. Science 2008; 322: 1211–7.
Eswar N, Webb B, Marti-Renom MA, Madhusudhan MS, Eramian D, Shen MY, et al. Comparative protein structure modeling using Modeller. Curr Protoc Bioinformatics 2006; Chapter 5: Unit 5.6.
Morris GM, Huey R, Lindstrom W, Sanner MF, Belew RK, Goodsell DS, et al. AutoDock4 and AutoDockTools4: automated docking with selective receptor flexibility. J Comput Chem 2009; 30: 2785–91.
Liu HQ, Zhang WY, Luo XT, Ye Y, Zhu XZ . Paeoniflorin attenuates neuroinflammation and dopaminergic neurodegeneration in the MPTP model of Parkinson's disease by activation of adenosine A1 receptor. Br J Pharmacol 2006; 148: 314–25.
Wetzker R, Böhmer FD . Transactivation joins multiple tracks to the ERK/MAPK cascade. Nat Rev Mol Cell Biol 2003; 4: 651–7.
Shuaib A, Hussain MS . The past and future of neuroprotection in cerebral ischaemic stroke. Eur Neurol 2008; 59: 4–14.
Zhang MQ, Zheng YL, Chen H, Tu JF, Shen Y, Guo JP, et al. Sodium tanshinone IIA sulfonate protects rat myocardium against ischemia-reperfusion injury via activation of PI3K/Akt/FOXO3A/Bim pathway. Acta Pharmacol Sin 2013; 34: 1386–96.
Zhao MM, Yang JY, Wang XB, Tang CS, Du JB, Jin HF . The PI3K/Akt pathway mediates the protection of SO2 preconditioning against myocardial ischemia/reperfusion injury in rats. Acta Pharmacol Sin 2013; 34: 501–6.
Tabata K, Matsumoto K, Murakami Y, Watanabe H . Ameliorative effects of paeoniflorin, a major constituent of peony root, on adenosine A1 receptor-mediated impairment of passive avoidance performance and long-term potentiation in the hippocampus. Biol Pharm Bull 2001; 24: 496–500.
Cheng JT, Wang CJ, Hsu FL . Paeoniflorin reverses guanethidine-induced hypotension via activation of central adenosine A1 receptors in Wistar rats. Clin Exp Pharmacol Physiol 1999; 26: 815–6.
Cunningham LA, Wetzel M, Rosenberg GA . Multiple roles for MMPs and TIMPs in cerebral ischemia. Glia 2005; 50: 329–39.
Rosell A, Lo EH . Multiphasic roles for matrix metalloproteinases after stroke. Curr Opin Pharmacol 2008; 8: 82–9.
Lappano R, Maggiolini M . G protein-coupled receptors: novel targets for drug discovery in cancer. Nat Rev Drug Discov 2011; 10: 47–60.
Lowes VL, Ip NY, Wong YH . Integration of signals from receptor tyrosine kinases and G protein-coupled receptors. Neurosignals 2002; 11: 5–19.
Lee FS, Rajagopal R, Chao MV . Distinctive features of Trk neurotrophin receptor transactivation by G protein-coupled receptors. Cytokine Growth Factor Rev 2002; 13: 11–7.
Chao MV, Rajagopal R, Lee FS . Neurotrophin signalling in health and disease. Clin Sci (Lond) 2006; 110: 167–73.
Acknowledgements
This work was supported by grants from National Natural Science Foundation of China (81123004) and the Scientific Innovation Project of the Chinese Academy of Sciences (XDA 01040304).
Author information
Authors and Affiliations
Corresponding author
Additional information
Supplementary Figure is available on the Acta Pharmacologica Sinica's website.
Supplementary information
Supplementary Information, Figure S1
Binding sites of PF, DPCPX, and CPA. (JPG 816 kb)
Rights and permissions
About this article

Cite this article
Zhong, M., Song, Wl., Xu, Yc. et al. Paeoniflorin ameliorates ischemic neuronal damage in vitro via adenosine A1 receptor-mediated transactivation of epidermal growth factor receptor. Acta Pharmacol Sin 36, 298–310 (2015). https://doi.org/10.1038/aps.2014.154
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/aps.2014.154
Keywords
This article is cited by
-
Transactivation of receptor tyrosine kinases by purinergic P2Y and adenosine receptors
Purinergic Signalling (2023)
-
Paeoniflorin ameliorates ischemic injury in rat brain via inhibiting cytochrome c/caspase3/HDAC4 pathway
Acta Pharmacologica Sinica (2022)
-
Caffeine and Its Neuroprotective Role in Ischemic Events: A Mechanism Dependent on Adenosine Receptors
Cellular and Molecular Neurobiology (2022)

{kind=link}