
Abstract
Several receptors that mediate apoptosis have been identified, such as Fas and tumor necrosis factor receptor I. Studies of the signal transduction pathways utilized by these receptors have played an important role in the understanding of apoptosis. Here we report the first ligand-receptor pair–the neuropeptide substance P and its receptor, neurokinin-1 receptor (NK1R)–that mediates an alternative, non-apoptotic form of programmed cell death. This pair is widely distributed in the central and peripheral nervous systems, and has been implicated in pain mediation and depression, among other effects. Here we demonstrate that substance P induces a non-apoptotic form of programmed cell death in hippocampal, striatal, and cortical neurons. This cell death requires gene expression, displays a non-apoptotic morphology, and is independent of caspase activation. The same form of cell death is induced by substance P in NK1R-transfected human embryonic kidney cells. These results argue that NK1R activates a death pathway different than apoptosis, and provide a signal transduction system by which to study an alternative, non-apoptotic cell death program.
Similar content being viewed by others


Introduction
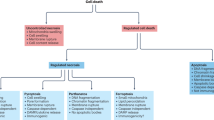
The original definition of apoptosis as a form of cell death distinct from necrosis was based on morphological criteria.1 Genetic studies in C. elegans led to the identification of key components of the apoptotic cell death pathway, including the protease Ced-3.2,3,4,5 However, it has become increasingly clear that the morphological descriptions of apoptosis and necrosis are not adequate to describe all cell deaths. For example, certain developmental cell deaths, such as ‘autophagic’ cell death6,7,8,9,10,11 and ‘cytoplasmic’ cell death,6,9,12,13,14,15 do not resemble apoptosis. Furthermore, neurodegenerative diseases, such as Huntington's disease and amyotrophic lateral sclerosis, demonstrate neuronal cell death that does not fulfil the criteria for apoptosis,16,17 but in some cases resembles a recently described form of cell death dubbed ‘paraptosis’.18 Ischemia-induced cell deaths may also display a non-apoptotic morphology, referred to as ‘oncosis’.19
The biochemical basis for these alternative morphological forms of cell death remains largely unknown. Understanding the mechanisms for these forms would have potentially important implications for the understanding of evolutionary aspects of cell death programs, developmental cell death, neurodegeneration, and cancer therapeutics, and for the design of novel therapeutic agents for diseases featuring these alternative forms of cell death.
Cell death has been divided into two main types: programmed cell death (pcd), in which the cell plays an active role in its own demise, and passive (necrotic) cell death. The pcd observed during development and tissue homeostasis has been classified morphologically into three main types: type 1, also known as nuclear or apoptotic; type 2 or autophagic; and type 3, a non-lysosomal vesiculate degeneration.9
Apoptosis is the best characterized type of pcd, in which the cells display membrane blebbing, flipping of phosphatidylserine (PS) in the plasma membrane,20 nuclear fragmentation, and activation of a family of cell-suicide cysteine proteases referred to as caspases (cysteinyl aspartic-specific proteases).5,21 The biochemical activation of apoptosis occurs through two general pathways: the intrinsic pathway, originating from mitochondrial release of cytochrome c and associated activation of caspase-9; and the extrinsic pathway, originating from the activation of cell surface death receptors such as Fas, and resulting in the activation of caspase-8.22
Much less is known about the biochemical mediators of type 2 and type 3 cell death. Type 2 (autophagic) cell death can be activated in some cases by Ras,23 while the molecular activation of type 3 cell death is unknown.
Interest in non-apoptotic forms of pcd has grown recently with the demonstration that the death of neurons in Huntington's disease is neither apoptotic nor necrotic.16 This work mirrors earlier work also showing non-apoptotic death in a transgenic mouse model of another neurodegenerative disease, amyotrophic lateral sclerosis.17 Similarly, neuronal cell death in other neurodegenerative diseases such as Alzheimer's disease and Parkinson's disease has not, to date, been shown to be mediated predominantly by apoptosis.
Substance P (SP) is an undecapeptide neurotransmitter belonging to the tachykinin family of peptides, members of which display some similarity in amino acid sequence in their carboxy-termini. SP is an alternatively spliced product of the preprotachykinin A gene (PPTA) that binds with nanomolar affinity to the neurokinin-1 receptor (NK1R). NK1R is a G-protein coupled receptor that is connected by various second messengers to a wide variety of effector mechanisms to modulate cellular function. Three apparently independent second messenger systems can be activated by G-proteins following ligand binding to NK1R: (1) Ca2+ mobilization from both intra- and extracellular sources via stimulation of phospholipase C; (2) arachidonic acid mobilization via phospholipase A2; and (3) cAMP accumulation via stimulation of adenylate cyclase.24 An alternative pathway, through interaction with β-arrestin, involves the activation of the MAPK pathway, leading to extracellular signal-regulated protein kinases 1 and 2 (ERKs) activation.25
The development of specific agonists and antagonists for NK1R has supported a role for NK1R in numerous biological processes, such as the transmission of pain in the spinal cord. In the CNS it also regulates cardiovascular and respiratory function, and is involved in activating the emetic reflex. NK1R also regulates several behavioral responses, and has recently been implicated in depression and schizophrenia. It appears to be involved in a wide variety of functions due to its ability to modulate the release of other neurotransmitters, such as excitatory amino acids.26
SP also seems to play an important role in pathological states in which neural cell death occurs, such as status epilepticus and ischemia. For example, SP null mice demonstrate resistance to excitotoxin-induced seizures, with an associated reduction in neuronal death.27 Similarly, treatment with an antagonist for NK1R inhibits seizures and reduces kainic acid-induced cell death in the CA1 region of the hippocampus.28 Furthermore, in a model of focal cerebral ischemia, administration of an NK1R antagonist reduced infarct volume and improved neurological function.29 Taken together, these observations suggest that NK1R may be a mediator of cell death in vivo.
In the present work, we studied the ability of SP to induce cell death in primary cultures of hippocampal, cortical and striatal neurons. We found that NK1R activates a caspase-independent cell death program that requires new gene expression to occur. Morphologically, it does not resemble apoptosis, since there is no membrane blebbing, nuclear fragmentation or phosphatidylserine translocation. Expression of NK1R in non-neuronal cells converted them into SP-sensitive cells, reproducing the cell death morphology that had been observed in neurons. Ultrastructural analysis of the cells revealed vacuolation and some phagolysosomes, while the plasma and nuclear membranes remained intact. SP-induced death in this system was not prevented by caspase inhibitors, nor by over-expression of Bcl-xL.
These results support the view that apoptosis is not the sole cellular death program, and provide a ligand-receptor pair and associated signal transduction system by which to dissect an alternative, non-apoptotic cell death program.
Results
SP induces cell death in neurons in primary culture via the NK1 receptor
Studies of the effects of SP on neurons stemmed from our earlier finding that SP fits a biophysical profile generated to allow database searches for pro-apoptotic peptides.30,31 Indeed, it was shown that SP induces cytochrome c release in a cell-free system of apoptosis.31 However, the pro-apoptotic effect of SP required internalization, since extracellular SP was not toxic to fibroblasts even at millimolar concentrations. Surprisingly, then, it was noted that nanomolar concentrations of SP were neurotoxic in primary cultures of striatal, cortical, and hippocampal neurons, inducing a delayed cell death. All three types of neurons were sensitive to micromolar concentrations of SP 48 h after exposure (Figure 1A–G), and to nanomolar concentrations 7 days after exposure (Figure 1H). However, the dying cells did not show apoptotic bodies, and instead appeared to be vacuolated (Figure 1B,D,F).

Substance P induces death of neurons in primary culture. (A,B), cortical cells; (C,D), hippocampal cells; (E,F), striatal cells. (A,C,E) are control cultures; (B,D,F) are cells grown in the presence of 100 μM SP for 48 h. Scale bar, 50 μm. G, decreased viability at 48 h in the presence of the indicated concentrations of L-SP in cortical, hippocampal or striatal neurons, or D-SP in cortical cells. Bars represent standard deviations (n=3). (H), SP triggers neuronal cell death at sub-micromolar concentrations. Striatal cells were grown in the presence of the indicated concentrations of SP for 7 days (SP was added only once at the beginning of the experiment). Results from a representative experiment are shown. The experiment was repeated three times with similar results
The selective toxicity of SP observed with neuronal cells in comparison to fibroblasts could conceivably be explained by a receptor-dependent or receptor-independent mechanism (e.g., a difference in membrane composition and membrane potential between neurons and fibroblasts). Therefore, we assessed the stereospecificity of SP's effect by comparing the sensitivity of cortical neurons to L-SP and D-SP. Only L-SP induced cell death (Figure 1G).
The stereospecific neurotoxicity elicited by SP suggested interaction with a receptor. To determine whether NK1R is the mediator of the SP-induced neuronal cell death, two different antagonists with relative selectivity for NK1R were added to the neuronal cultures along with the SP. The L-tryptophan derivative L-732,138 is a competitive antagonist,32 and L-733,060 is a non-peptide antagonist.33 Both molecules completely inhibited SP-induced cell death in striatal cells, as shown in Figure 2A, arguing that SP-induced neurotoxicity is mediated by NK1R. Immunostaining of striatal, hippocampal, and cortical neurons showed that all three of these neuronal types express NK1R (data not shown).

SP-induced death of primary neurons requires NK1R activation, gene expression, is independent of caspases and nuclear fragmentation does not occur. (A) Striatal cells were grown in the presence or absence of 100 μM SP for 48 h. The antagonists L-733,060 (1 nM) or L-732,138 (5 nM) were added at the same time as SP. (B) Striatal cells were grown for 48 h in the presence or absence of 100 μM SP. At the same time, Actinomycin D (ActD), Cycloheximide (CHX), zVAD.fmk or BAF was added. Viability was quantified by Trypan blue exclusion, and represented in comparison to the control. Bars represent standard deviations (n=3). *P<0.0001 by ANOVA test. DAPI staining of striatal cells grown in the absence (C) or presence (D) of 100 μM SP for 48 h, or with 50 μM tamoxifen (Tmx) for 3 h (E). Scale bar, 50 μm
SP-induced death is neither apoptotic nor necrotic
In order to distinguish between an active cell death program and a passive cell death process, SP-induced neuronal cell death was evaluated in the presence of the transcription inhibitor Actinomycin D and the translation inhibitor Cycloheximide. Both prevented the SP toxicity in cortical and striatal neurons, arguing against its being a passive cellular process (Figure 2B and data not shown).
To determine whether caspases, the cysteine proteases that mediate apoptosis, are required for SP-induced neuronal cell death, the effects of the broad-spectrum caspase inhibitors zVAD.fmk and Boc-aspartyl.fmk (BAF) were evaluated. Neither had any effect on SP-induced neuronal cell death, indicating that this form of cell death is caspase-independent (Figure 2B).
In addition to caspase dependence, other hallmarks of apoptosis include apoptotic body formation, membrane bleb formation, nuclear fragmentation, and loss of the normal asymmetry of phosphatidylserine within the plasma membrane. These were evaluated in neurons undergoing SP-induced cell death. DAPI staining demonstrated no nuclear fragmentation, although some degree of chromatin condensation occurred. As a control, tamoxifen was used to induce apoptosis, and in that case nuclear fragmentation was readily detected (Figure 3). Annexin-V staining showed no evidence for phosphatidylserine flipping (data not shown).

Expression of NK1R renders cells sensitive to SP-induced death 293T cells were transfected with the empty vector pcDNA3.1 (A,C,E) or with NK1R (B,D,F) and challenged by the addition of different concentrations of SP. (A,B), control cultures; (C,D), cells grown in the presence of 10 nM SP; (E,F), cells grown in the presence of 100 μM SP. Pictures were taken 24 h after SP addition. Scale bar, 50 μm
Thus SP-induced neuronal death required gene expression, was independent of caspase activation, and showed no evidence of membrane bleb formation, apoptotic body formation, nuclear fragmentation, or loss of phosphatidylserine asymmetry.
NK1R is a death receptor that mediates non-apoptotic programmed cell death
As noted above, antagonists of NK1R blocked SP-induced neuronal cell death, indicating that NK1R mediates this type of cell death. It was conceivable that this cell death may require both NK1R and some neuron-specific factor or factors. In order to determine whether NK1R-mediated cell death requires neuron-specific factors, we expressed NK1R in 293T human embryonic kidney cells, which do not express endogenous NK1R. Whereas 293T cells are resistant to SP even at millimolar concentrations,31 the expression of NK1R in 293T cells rendered them sensitive to SP at concentrations of 0.1 nM or greater, displaying the same morphology up to 100 μM (Figures 4A–F and 10A). It is noteworthy that 0.1 nM is similar to the affinity of the NK1R for SP.24

Lack of nuclear fragmentation in SP-induced death in non-neuronal cells. 293T cells transfected with either the empty vector pcDNA3.1 (A,D,G) or NK1R (B,C,E,F,H,I) were immunostained to detect NK1R expression and stained with Hoechst to reveal the nuclear morphology. Note that the rounded cells correspond to the NK1R-expressing ones. 10 μM SP was added for 24 h to cells shown in panels C, F and I. Scale bar, 50 μm
The morphology of NK1R-mediated cell death in 293T cells did not present features typical of apoptosis, but instead resembled the process of NK1R-mediated cell death observed in primary neurons. The cells rounded up but demonstrated no bleb formation, apoptotic body formation, cell shrinkage or nuclear fragmentation (Figure 5A–I). Instead, electron microscopic analysis revealed swelling and vacuolation (the observed vacuolation of the 293T cells was unlikely to be due to a necrotic process, since the vacuolation preceded membrane permeability to Trypan blue by over 24 h) (Figure 6). Furthermore, Annexin-V staining was again negative in NK1R-mediated death of 293T cells (Figure 7E,F), whereas controls transfected with a Bax expression construct demonstrated clear bleb formation and Annexin-V staining (Figure 7G,H). Similarly, TUNEL (TdT-mediated dUTP Nick-End Labeling) staining to detect DNA fragmentation was negative for NK1R-mediated cell death (Figure 8A,B), while control cells expressing Bax developed a positive signal (Figure 8C,D).

Ultrastructure of NK1R-mediated cell death. Electron micrographs of 293T cells 24 h after treatment with 10 μM SP. (A) Control cells exhibited normal nuclear and cytoplasmic ultrastructure and the usual complement of organelles. (B) Less affected SP-treated cells were somewhat rounded and swollen, had minor changes in nuclear outline and chromatin organization, and contained peroxisomes, phagolysosomes, and numerous membrane-bound vacuoles, a few occasionally within the nuclear matrix. (C) More severely affected SP-treated cells showed disorganized nuclei, loss of normal subcellular organelles, the appearance of lysosomal and multivesicular bodies, and large cytoplasmic vacuoles. No cells appeared to have pyknotic or fragmented nuclei, membrane blebbing, or other hallmarks of apoptosis

Retention of phosphatidylserine asymmetry (lack of phosphatidylserine ‘flipping’) and absence of DNA fragmentation in SP-induced cell death. Cells transfected with NK1R in the absence (A–C) or presence of 10 μM SP for 24 h (D,E) were stained with FITC-Annexin-V (B,E), then fixed and immunostained to detect NK1R (C,F), demonstrating that the cells expressing NK1R were negative for FITC-Annexin-V staining. As a control for apoptosis, cells were transfected with Bax (G,H) and stained with FITC-Annexin-V (H). Scale bar, 50 μm. Note that the few cells that are Annexin-V positive in (E) are not those expressing NK1R (compare F). (I) TUNEL staining of 293T cells transfected with NK1R and treated for 24 h with 10 μM SP. (J) Propidium iodide (PI) staining of the same field as in I, to show the total number of cells. (K) TUNEL staining of cells transfected with Bax as a positive control for apoptosis. (L) Propidium iodide staining of the same field as in K

NK1R-mediated cell death is independent of caspase activation. (A) 293T cells transfected with either the empty vector pcDNA3.1 (white bars) or NK1R construct (black bars) were incubated for 48 h in the presence of the caspase inhibitors indicated. (B) 293T cells transfected with caspase-8 cDNA were incubated during 24 h in the presence of the caspase inhibitors indicated, to demonstrate the functionality of the compounds. (C) SP did not induce hydrolysis of specific substrates for either ‘initiator’ caspases-1 (YVAD), -2 (VDVAD), -8 (IETD), -9 (LEHD), or ‘executioner’ caspases-3, -7 (DEVD) and -6 (VEID). (D) Activation of procaspase-3 by the serine protease granzyme B. The rates of substrate hydrolysis are expressed in absolute units μM.s−1

NK1R-mediated cell death is not prevented by the inhibitor of apoptosis Bcl-XL. Co-transfection of 293T cells with the indicated plasmids, followed by growth in the absence (white bars) or presence (black bars) of 10 μM SP for 48 h
Since SP had been shown to display toxicity toward bacteria and mitochondria,31 it was still possible that the mediation of SP-induced death by NK1R was due to internalization of SP and subsequent mitochondrial toxicity, rather than resulting from the triggering of a signal cascade mediated by NK1R. Therefore, neurokinin A and SP6–11, which are NK1R agonists that do not exhibit mitochondrial toxicity, were evaluated. We observed that these molecules were able to induce cell death as well as SP (data not shown), suggesting that NK1R mediates SP-induced neuronal cell death by a mechanism independent of direct mitochondrial damage.
As noted above, the caspase inhibitors zVAD.fmk and BAF did not inhibit NK1R-mediated cell death in neurons. We found a similar lack of effect in 293T cells (Figure 9A), while the apoptosis induced by over-expression of caspase-8 was completely abolished, indicating that the inhibitors were effective (Figure 9B). Accordingly, no hydrolysis of substrates for either ‘initiator’ caspases-1, -2, -8, -9 and 10, or ‘executioner’ caspases-3, -6 and -7 was detected (Figure 9C). It has been demonstrated that procaspase-3 is present in 293T cells34 and that it is activated by the serine protease granzyme B.35 To ensure that the lack of DEVDase activation was not simply due to the lack of activable pro-caspase-3, we added granzyme B to each cytosolic extract of 293T cells transfected with NK1R, treated (or not) with SP. The addition of granzyme B activated the procaspase-3, as shown by a sevenfold increase of DEVDase activity, whereas no significant difference was observed in response to SP stimulation (Figure 9D). These results indicate that the lack of DEVD cleavage during SP-induced cell death was not due to the presence of a non-functional procaspase-3. The activation of other endopeptidases was also tested, such as calpains (requiring micromolar or millimolar concentrations of calcium) and the chymotrypsin-like activity of the proteosome, and no significant effect on the rates of substrate hydrolysis was detected. We found further that Bcl-xL expression did not inhibit NK1R-mediated cell death (P=0.486 by ANOVA test) (Figure 10).
Discussion
It is becoming increasingly clear that non-apoptotic forms of programmed cell death exist,9,18,36 yet, in marked contrast to apoptotic pcd, very little is known about the biochemical mechanisms involved in non-apoptotic pcd. The importance of defining these mechanisms has been highlighted by recent descriptions of non-apoptotic cell death in neurodegenerative conditions such as Huntington's disease16 and amyotrophic lateral sclerosis,17 coupled with the finding that alternative programs of cell death are not affected by apoptosis inhibitors.18 Thus, elucidation of the biochemical mechanisms of non-apoptotic forms of pcd may contribute to the development of therapies that modulate the neurodegenerative process.
In the present work we have described the first ligand-receptor pair that mediates a non-apoptotic form of pcd. Since the discovery of Fas-FasL–a ligand-receptor pair that mediates apoptotic cell death–facilitated the identification of apoptotic pathways such as the Fas-FADD-caspase-8 pathway,37 the identification of the pathway by which NK1R mediates non-apoptotic pcd should facilitate the definition of biochemical factors involved in non-apoptotic pcd.
The requirement for gene expression indicates that NK1R-mediated cell death is programmatic; however, the lack of nuclear fragmentation, bleb formation, apoptotic body formation, phosphatidylserine flipping, and caspase dependence, as well as the lack of effect of Bcl-xL, all argue against apoptosis as the form of pcd mediated by NK1R. The presence of vacuoles also argues against apoptotic pcd, and instead suggests either type 2 (autophagic) or type 3 (non-lysosomal vesiculate) pcd (or a previously undescribed form of pcd). The origin of the vacuoles in type 2 pcd is lysosomal, whereas in type 3 pcd the origin of the vacuoles is the endoplasmic reticulum and mitochondria. Further studies will be required to determine whether NK1R-mediated cell death is type 2 or type 3 pcd or a novel type of pcd.
It has been shown previously that SP induces glutamate release in the hippocampus38 and enhances glutamate responses.39 Thus, the question arises as to whether the SP-induced neuronal cell death we observed was due to glutamate toxicity. This is unlikely for two reasons: first, it requires approximately 10 days for primary neuronal cultures to become sensitive to receptor-mediated glutamate toxicity, and we observed the SP toxicity during the first 7 days of culture. Second, we were able to reproduce the same morphological features of neuronal cell death in 293T cells that had been transfected with a NK1R expression construct, despite the fact that 293T cells do not express glutamate receptors and are not sensitive to receptor-mediated glutamate toxicity. We also excluded the possibility that SP acts as a direct mitochondrial toxic peptide, since other NK1R agonists such as NKA or SP6–11 (which lack direct mitochondrial toxicity) killed as well as SP, while the enantiomer D-SP (which does exhibit mitochondrial toxicity but is not an NK1R agonist) had no effect.
A member of the NK1R-mediated kinase activation pathway–(Ras)–has been implicated previously in caspase-independent pcd.23 Although further work will be required to characterize the signal pathway involved in NK1R-mediated non-apoptotic pcd, preliminary data suggest that a MAPK pathway is involved (data not shown).
The kinetics of neuronal cell death induced by SP and mediated by NK1R are relatively slow, with cell death occurring within approximately 2–7 days, depending on the concentration of SP. Since the half-life of SP in solution is only a few minutes and we administered the SP only once at the beginning of each experiment, the long delay in cell death appears to be due to the cellular response rather than any prolonged presence of the SP. This offers a system to study gene expression in neurons that are morphologically normal (during the first days after exposure to SP) yet are committed to pcd.
As noted above, recent work has highlighted neurodegenerative cell death, demonstrating that it fits the criteria for neither apoptosis nor necrosis.16,17 Interestingly, the cell death morphologies described in both of these cases–a transgenic model of amyotrophic lateral sclerosis17 and Huntington's disease16–featured vacuoles in the dying neurons, suggesting the possibility that a non-apoptotic form of pcd may be involved. Furthermore, it is possible that some of the cell death previously described as necrotic, based on morphology, may be due to an alternative, non-apoptotic program of cell death.
It is also noteworthy that, for certain pathological states such as status epilepticus and neuronal ischemia, a burst of SP release has been reported to occur, prior to neuronal cell death.29,38,40 For instance, SP is over-expressed in hippocampal neurons during status epilepticus, and intrahippocampal administration of SP triggers self-sustained status epilepticus, generating a pattern of acute hippocampal damage resembling that known to occur in human epilepsy.38 Furthermore, in some neurodegenerative diseases, a reduction in SP-immunostaining neurons has been demonstrated in specific brain regions: in the inferior temporal cortex and hippocampus in Alzheimer's disease, and in the pars compacta and pars reticulata of the substantia nigra in both Huntington's and Parkinson's diseases.40 Unfortunately, there is little information published on the pattern of expression of NK1R in these diseases, so that it is currently unclear whether the loss of SP-immunostaining cells in these diseases involves NK1R.
In addition to playing a role in cell death, SP has been proposed to function as a neurotrophic factor, so the loss of SP-immunopositive neurons could cause a loss of trophic support rather than simply reflecting neurodegeneration.40 However, the cells that die are the cells that contain SP, rather than the cells that lack it. Compatible with our results, a previous report suggested a neurotoxic role for SP in hippocampal cells, since it was observed that the addition of SP abrogated a neurotrophic activity displayed by the amyloid β1-40 peptide.41 Others have also observed a killing activity of SP on proliferating non-neuronal cells.42 The apparent discrepancy in proposed roles for SP may be explicable by differences in cell type, age and SP concentrations used in different studies. Also, other forms of the NK1R have been identified, and although their specific functions remain unknown,24 it is possible that the different responses to SP observed in the other systems are mediated by different receptor complexes.
In summary, SP-NK1R represents the first ligand-receptor pair described that mediates non-apoptotic pcd. These results support the view that apoptosis is not the sole cell death program, and provide a signal transduction system by which to study an alternative, non-apoptotic cell death program. Whether NK1R-mediated neuronal cell death plays a role in neurodegenerative disease states remains to be determined; however, it is increasingly clear that apoptosis, at least as classically described, does not represent the major mode of cell death in these diseases. Thus, defining the pathways involved in non-apoptotic forms of pcd may lead to novel therapies for these diseases.
Materials and Methods
Neuronal primary culture
Primary striatal, cortical and hippocampal cultures were prepared from 17-day-old Sprague–Dawley rat embryos (B&K, Fremont, CA, USA). The tissue was dissected, minced and trypsinized for 5 min using 0.25% trypsin (Cell Grow). After the addition of 10% horse serum to inhibit the trypsin, the cell suspension was triturated 15–20 times with a 10 ml syringe and centrifuged for 5 min at 800×g. The pellet was resuspended in MEM–PAK (UCSF Cell Culture facility, San Francisco, CA, USA), supplemented with 2.02 gm glucose, 2 mM of GlutaMax (Life Technologies, Grand Island, NY, USA) and penicillin/streptomycin (100 U/ml). The suspension was filtered through a 70 μm cell strainer and the final culture medium contained 5% horse serum (Life Technologies). 3–4×105 cells per cm2 were seeded onto either poly-D-lysine precoated 8-well chamber slides (Becton-Dickson Labware, Bedford, MA, USA) or 96-well plates pre-coated with 50 μg/ml of poly-D-lysine (Sigma, St. Louis, MO, USA) in water. After 30 min incubation, unattached cells were removed together with the medium and replaced with glucose enriched MEM-PAK plus 5% horse serum (Life Technologies). The cultures were incubated at 37°C in 95% air 5% carbon dioxide with 95% humidity. Cultures were used for experiments between day 1 and day 7 when glial contaminants were at a minimum.
SP (SIGMA) 0.1 nM–100 μM was added 24–48 h after seeding, in the presence of 2.5% horse serum. When the cultures were maintained for more than 5 days, 10 μM cytosine arabinoside was added for 24 h, beginning 3 days after plating the cells. The following compounds were added at the same time as SP in the indicated experiments: 5 nM L-732,138; 1 nM L-733,060; 1 μg/ml Actinomycin D or 1 μg/ml Cycloheximide. All of these compounds were obtained from SIGMA. zVAD.fmk or BAF was added at 50 μM and obtained from Enzyme System Products, Livermore, CA, USA. 4-Hydroxytamoxifen from SIGMA was added at 50 μM.
Viability was quantified by Trypan blue staining of the total cell population.
Induction of death in transfected human embryonic kidney 293T cells
The full-length (1.2 kb) NK1R cDNA was cloned by RT–PCR from adult rat hippocampal RNA into the XbaI/EcoRI sites of the vector pcDNA3.1− (Invitrogen), using the following oligonucleotides: forward 5′-GGTCTATTGCCCCAAAATGGATAACG-3′; reverse 5′-CTGAATTCAAGATGGCTAGAGGCATGCC-3′. The mutants were created using the QuickChange strategy (Stratagene, La Jolla, CA, USA), using the following sequences for the oligonucleotides: 5′-GTGGACCTGGCCTTCGCTCAGGCCTGCATGGCTGCATTC-3′ for NK1RE78Q mutation and 5′-GGAAATGAAATCCACCCGATACCTCTAGGCATATTCAAGGCATGC-3′ for NK1RΔ342 truncation. Human embryonic kidney 293T cells were grown in high glucose DMEM (Life Technologies, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (SIGMA) and penicillin/streptomycin 100 U/ml (Life Technologies, Grand Island, NY, USA). The cultures were incubated at 37°C in 95% air 5% carbon dioxide with 95% humidity. Transient transfection was performed using Superfect (Qiagen) as described by the manufacturer. Briefly, 2×105 cells/well were seeded into 35 mm wells 16 h prior to transfection. Superfect : DNA complexes were added at a 5 μl : 2 μg ratio for 3 h; then the medium was replaced and after 24 h SP was added. After 24–48 h addition of SP cell death was quantified as follows: the media containing floating cells were collected and centrifuged 10 min at 2000×g, the pellet was resuspended in PBS/Trypan blue and the cells were counted using a hematocytometer. When other compounds were used, they were added at the same time as SP:zVAD.fmk or BAF was added at 50 μM and obtained from Enzyme System Products, Livermore, CA, USA.
Nuclear staining
Either neuronal primary cultures or human embryonic kidney 293T cells were fixed with 4% paraformaldehyde and stained with 0.5 μg/ml DAPI (Molecular Probes) or Hoechst (SIGMA).
NK1R immunostaining
Cells were fixed 10 min with 4% paraformaldehyde; permeabilized 10 min with 2% Triton X-100/phosphate-buffered saline (PBS); washed with PBS, blocked 30 min with 4% goat serum/4%BSA/PBS; washed with PBS; incubated 16 h at 4°C with a 1 : 500 dilution of anti-NK1R (SIGMA); washed with PBS; incubated 30 min with 1 : 10,000 anti-rabbit coupled to FITC or Texas Red (Molecular Probes) and washed with PBS.
Electron microscopy
Human embryonic kidney 293T cells were grown to 50% confluency on poly-L-lysine-coated Thermanox™ coverslips (13 mm, round, VWR-Scientific Products) and transfected with NK1R expression construct as described above. Pairs of duplicate coverslips were treated with 10 μM SP or vehicle alone for 24 h, and then fixed with 2.0% paraformaldehyde/0.2% glutaraldehyde in 150 mM sodium cacodylate buffer, pH 7.4 (CB) at RT. Fixation was continued at 4°C until the cells were washed well and postfixed in 2% osmium tetroxide (OsO4) (Electron Microscopy Sciences). They were reacted with 0.5% tannic acid (Sigma) followed by 1%NaSO4, and stained with 5% uranyl acetate in 50% ethanol during dehydration. Cells were flat embedded in Epon 812 between a second Thermanox™ coverslip, and three or four samples from each sandwich were re-embedded in Epon blocks. En face thin sections were counterstained with uranyl acetate and lead citrate, and examined at 80 kV in a FEI-Philips Tecnai 12 (FEI-Philips, Hillsborough, OR, USA). Digital CCD images (Gatan Bioscan, Livermore, CA, USA) were acquired in Digital Micrograph 1.0 (FEI-Philips) and trimmed in Adobe Photoshop 5.0.
Annexin-V staining
Cells were stained according to the manufacturer's instructions (Clontech). Briefly, cells were washed with binding buffer and incubated with FITC-Annexin-V or EGFP-Annexin-V for 5–10 min and fixed with 4% paraformaldehyde.
TUNEL staining
Cells were fixed 10 min with 4% paraformaldehyde and stained according to the manufacturer's instructions (Promega).
Protease activation during SP-induced cell death
Preparation of cell-free extract for kinetic studies: 293T cells seeded at the density of 2 million cells per 10 cm dish were transfected with NK1R cDNA using Lipofectamine 2000 (Gibco) according the instructions provided by the manufacturer. After 24 h 10 nM SP was added to half of the culture dishes. Cells were washed 24 h later with ice-cold PBS and collected with a cell scraper. The cells pelleted at 700×g for 4 min were resuspended in cell lysis buffer at pH 7.4 containing 1 mM dithiothreitol at a ratio of 1 : 1 (v/v). After 30 min on ice the swelled cells were disrupted with 20 strokes using a 27 gauge needle and further spun at 16 000×g for 30 min. The pellet was discarded and the supernatant which represents the cytosolic fraction was aliquoted and stored at −80°C or used in the kinetic studies. The protein concentration was determined using the Bradford assay (Pierce).
Kinetic studies: the proteolytic activities were followed fluorometrically by monitoring AFC or AMC release from 100 μM of the corresponding substrates (Enzyme Systems Products): Caspase-1 like, Ac-YVAD-AFC; caspase-2, Ac-VDVAD-AFC; caspases-3 like, Ac-DEVD-AFC; caspase-6, Ac-VEID-AFC; caspase-8, Ac-IETD-AFC; caspase-9, Ac-LEHD-AFC. Calpain activity was monitored with 100 μM Suc-LY-AFC (succinyl-Leu-Tyr-7-amino-4-methylcoumarin, Enzyme Systems Products) at 5 μM and 5 mM CaCl2. The chymotrypsin activity of the proteosome was followed with 100 μM Suc-LLVY-AFC (succinyl-Leu-Leu-Val-Tyr-7-amino-4-methylcoumarin, Enzyme Systems Products), and cathepsins were assayed using 20 μM Z-FR-AMC (Benzyloxycarbonyl-Phe-Arg-7-amino-4-methylcoumarin, Bachem). A 10 μl aliquot of extract in the presence or absence of SP was added to 90 μl of substrate at the appropriate concentration and pH. The reaction was continuously monitored during 30 min and 37°C in a fluorescence microplate reader (Molecular Devices). The instrument was set at excitation and emission wavelengths of 405 and 510 for AFC releasing group, and 370 and 460 nm, for AMC releasing group, respectively. The steady-state hydrolysis rates were obtained from the linear part of the curves.
Abbreviations
- zVAD.fmk:
-
Benzyloxycarbonyl-Val-Ala-DL-Asp-fluoromethylketone
- BAF:
-
Boc-Asp(Ome)CH2F
- Rp-8-Br-cAMPS:
-
Adenosine 3′,5′-cyclic Monophosphorothioate, 8-Bromo-,Rp-Isomer
- SP:
-
Substance P
- NK1R:
-
Neurokinin-1 receptor
- pcd:
-
programmed cell death
- PS:
-
phosphatidylserine
- PPTA:
-
preprotachykinin A gene
- TUNEL:
-
TdT-mediated dUTP nick-end labeling
- AFC:
-
7-amino-4-trifluoromethylcoumaride
- AMC:
-
7-amino-4-methylcoumarin
References
Kerr JF, Wyllie AH, Currie AR . 1972 Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics Br. J. Cancer 26: 239–257
Ellis HM, Horvitz HR . 1986 Genetic control of programmed cell death in the nematode C. elegans Cell 44: 817–829
Hengartner MO, Horvitz HR . 1994 Activation of C. elegans cell death protein CED-9 by an amino-acid substitution in a domain conserved in Bcl-2 see comments Nature 369: 318–320
Hengartner MO, Horvitz HR . 1994 C. elegans cell survival gene ced-9 encodes a functional homolog of the mammalian proto-oncogene bcl-2 Cell 76: 665–676
Yuan J, Shaham S, Ledoux S, Ellis HM, Horvitz HR . 1993 The C. elegans cell death gene ced-3 encodes a protein similar to mammalian interleukin-1 beta-converting enzyme Cell 75: 641–652
Schweichel JU . 1972 [Elektron microscopic studies on the degradation of the apical ridge during the development of limbs in rat embryos] Z. Anat. Entwicklungsgesch. 136: 192–203
Schweichel JU, Merker HJ . 1973 The morphology of various types of cell death in prenatal tissues Teratology 7: 253–266
Schwartz LM . 1991 The role of cell death genes during development BioEssays 13: 389–395
Clarke PG . 1990 Developmental cell death: morphological diversity and multiple mechanisms Anat. Embryol. 181: 195–213
Lockshin RA, Williams CM . 1964 Programmed cell death. II. Endocrine potentiation of the breakdown of the intersegmental muscles of silkmoths J. Insect Physiol. 10: 643–649
Xue L, Fletcher GC, Tolkovsky AM . 1999 Autophagy is activated by apoptotic signalling in sympathetic neurons: an alternative mechanism of death execution Mol. Cell. Neurosci. 14: 180–198
Pilar G, Landmesser L . 1976 Ultrastructural differences during embryonic cell death in normal and peripherally deprived ciliary ganglia J. Cell. Biol. 68: 339–356
Oppenheim RW . 1991 Cell death during development of the nervous system Annu. Rev. Neurosci. 14: 453–501
Oppenheim RW . 1985 Naturally occurring cell death during neural development Trends Neurosci. 17: 487–493
Cunningham TJ . 1982 Naturally occurring neuron death and its regulation by developing neural pathways Int. Rev. Cytol. 74: 163–186
Turmaine M, Raza A, Mahal A, Mangiarini L, Bates GP, Davies SW . 2000 Nonapoptotic neurodegeneration in a transgenic mouse model of Huntington's disease Proc. Natl. Acad. Sci. USA 97: 8093–8097
Dal Canto MC, Gurney ME . 1994 Development of central nervous system pathology in a murine transgenic model of human amyotrophic lateral sclerosis Am. J. Pathol. 145: 1271–1279
Sperandio S, de Belle I, Bredesen DE . 2000 An alternative, non-apoptotic form of programmed cell death Proc. Natl. Acad. Sci. USA 97: 14376–14381
Majno G, Joris I . 1995 Apoptosis, oncosis, and necrosis. An overview of cell death [see comments] Am. J. Pathol. 146: 3–15
Fadok VA, Voelker DR, Campbell PA, Cohen JJ, Bratton DL, Henson PM . 1992 Exposure of phosphatidylserine on the surface of apoptotic lymphocytes triggers specific recognition and removal by macrophages J. Immunol. 148: 2207–2216
Thornberry NA, Lazebnik Y . 1998 Caspases: enemies within Science 281: 1312–1316
Salvesen GS, Dixit VM . 1997 Caspases: intracellular signaling by proteolysis Cell 91: 443–446
Chi S, Kitanaka C, Noguchi K, Mochizuki T, Nagashima Y, Shirozou M, Fujita H, Yoshida M, Chen W, Asai A, Himeno M, Yokoyama S, Kuchino Y . 1999 Oncogenic Ras triggers cell suicide through the activation of a caspase-independent cell death program in human cancer cells Oncogene 18: 2281–2290
Quartara L, Maggi CA . 1997 The tachykinin NK1 receptor. Part I : ligands and mechanisms of cellular activation Neuropeptides 31: 537–563
Defea K, Schmidlin F, Dery O, Grady EF, Bunnett NW . 2000 Mechanisms of initiation and termination of signalling by neuropeptide receptors: a comparison with the proteinase-activated receptors Biochem. Soc. Trans. 28: 419–426
Quartara L, Maggi CA . 1998 The tachykinin NK1 receptor. Part II: Distribution and pathophysiological roles Neuropeptides 32: 1–49
Liu H, Cao Y, Basbaum AI, Mazarati AM, Sankar R, Wasterlain CG . 1999 Resistance to excitotoxin-induced seizures and neuronal death in mice lacking the preprotachykinin A gene Proc. Natl. Acad. Sci. USA 96: 12096–12101
Zachrisson O, Lindefors N, Brene S . 1998 A tachykinin NK1 receptor antagonist, CP-122,721-1, attenuates kainic acid-induced seizure activity Brain Res. Mol. Brain. Res. 60: 291–295
Yu Z, Cheng G, Huang X, Li K, Cao X . 1997 Neurokinin-1 receptor antagonist SR140333: a novel type of drug to treat cerebral ischemia Neuroreport 8: 2117–2119
Ellerby HM, Arap W, Ellerby LM, Kain R, Andrusiak R, Rio GD, Krajewski S, Lombardo CR, Rao R, Ruoslahti E, Bredesen DE, Pasqualini R . 1999 Anti-cancer activity of targeted pro-apoptotic peptides Nat. Med. 5: 1032–1038
del Rio G, Castro-Obregon S, Rao R, Ellerby HM, Bredesen DE . 2001 APAP, a sequence-pattern recognition approach identifies substance P as a potential apoptotic peptide FEBS Lett. 494: 213–219
MacLeod AM, Merchant KJ, Cascieri MA, Sadowski S, Ber E, Swain CJ, Baker R . 1993 N-acyl-L-tryptophan benzyl esters: potent substance P receptor antagonists J. Med. Chem. 36: 2044–2045
Seabrook GR, Shepheard SL, Williamson DJ, Tyrer P, Rigby M, Cascieri MA, Harrison T, Hargreaves RJ, Hill RG . 1996 L-733,060, a novel tachykinin NK1 receptor antagonist; effects in [Ca2+]i mobilisation, cardiovascular and dural extravasation assays Eur. J. Pharmacol. 317: 129–135
Stennicke HR, Jurgensmeier HM, Shin H, Deveraux Q, Wolf BB, Yang X, Zhou Q, Ellerby HM, Ellerby LM, Bredesen D, Green DR, Reed JC, Froelich CJ, Salvesen GS . 1998 Pro-caspase-3 is a major physiologic target of caspase-8 J. Biol. Chem. 273: 27084–27090
Atkinson EA, Barry M, Darmon AJ, Shostak I, Turner PC, Moyer RW, Bleackley RC . 1998 Cytotoxic T lymphocyte-assisted suicide. Caspase 3 activation is primarily the result of the direct action of granzyme B J. Biol. Chem. 273: 21261–21266
Leist M, Jaattela M . 2001 Four deaths and a funeral: from caspases to alternative mechanisms Nat. Rev. Mol. Cell. Biol. 2: 589–598
Muzio M, Chinnaiyan AM, Kischkel FC, O'Rourke K, Shevchenko A, Ni J, Scaffidi C, Bretz JD, Zhang M, Gentz R, Mann M, Krammer PH, Peter ME, Dixit VM . 1996 FLICE, a novel FADD-homologous ICE/CED-3-like protease, is recruited to the CD95 (Fas/APO-1) death-inducing signaling complex Cell 85: 817–827
Liu H, Mazarati AM, Katsumori H, Sankar R, Wasterlain CG . 1999 Substance P is expressed in hippocampal principal neurons during status epilepticus and plays a critical role in the maintenance of status epilepticus Proc. Natl. Acad. Sci. USA 96: 5286–5291
Urban L, Thompson SW, Dray A . 1994 Modulation of spinal excitability: co-operation between neurokinin and excitatory amino acid neurotransmitters Trends Neurosci. 17: 432–438
Raffa RB . 1998 Possible role(s) of neurokinins in CNS development and neurodegenerative or other disorders Neurosci. Behav. Rev. 22: 789–813
Yankner BA, Caceres A, Duffy LK . 1990 Nerve growth factor potentiates the neurotoxicity of beta amyloid Proc. Natl. Acad. Sci. USA 87: 9020–9023
DeFea KA, Vaughn ZD, O'Bryan EM, Nishijima D, Dery O, Bunnett NW . 2000 The proliferative and antiapoptotic effects of substance P are facilitated by formation of a beta-arrestin-dependent scaffolding complex Proc. Natl. Acad. Sci. USA 97: 11086–11091
Acknowledgements
We thank Drs. Paul Goldsmith and Anna Logvinova for the electron microscopic analysis, and Drs. H Michael Ellerby and Sabina Sperandio for thoughtful discussion. Susana Castro-Obregón is a Pew Latin American Fellow. Harald Frankowski was supported by grant number 1Ge-53240 from the Swiss National Science Foundation. Supported by NIH grants NS33376 and AG12282 to DE Bredesen.
Author information
Authors and Affiliations
Corresponding author
Additional information
Edited by C J Thiele
Rights and permissions
About this article
Cite this article
Castro-Obregón, S., del Rio, G., Chen, S. et al. A ligand-receptor pair that triggers a non-apoptotic form of programmed cell death. Cell Death Differ 9, 807–817 (2002). https://doi.org/10.1038/sj.cdd.4401035
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.cdd.4401035
Keywords
This article is cited by
-
Substance P failed to reverse dextran sulfate sodium-induced murine colitis mediated by mitochondrial dysfunction: implications in ulcerative colitis
3 Biotech (2021)
-
An RNA interference screen identifies new avenues for nephroprotection
Cell Death & Differentiation (2016)
-
Antagonist of the neurokinin-1 receptor curbs neuroinflammation in ex vivo and in vitro models of Lyme neuroborreliosis
Journal of Neuroinflammation (2015)
-
Involvement of substance P and the NK-1 receptor in human pathology
Amino Acids (2014)
-
Singapore grouper iridovirus, a large DNA virus, induces nonapoptotic cell death by a cell type dependent fashion and evokes ERK signaling
Apoptosis (2011)
