
Abstract
Subanesthetic doses of the noncompetitive N-methyl-D-aspartate (NMDA) antagonist ketamine exacerbate psychosis in schizophrenic patients, and ketamine has significant abuse liability. These observations indicate that a secondary effect of ketamine may be to increase dopamine concentrations. The present study was undertaken using positron emission tomography (PET) and the dopamine (D2) radiotracer 11C-raclopride to determine whether ketamine would decrease D2 receptor availability, indicative of an increase in dopamine concentrations. Two scans were performed in seven male control subjects before and after administration of ketamine (0.5 mg/kg, IV infused over 20 min). Ketamine significantly increased cortisol levels and decreased dopamine receptor availability in the striatum (specific binding), but not in the cerebellum (nonspecific binding). In addition, the cerebellar binding subtracted from the striatal binding (to account for changes in nonspecific binding) was significantly decreased after ketamine administration. These results provide in vivo evidence for the ability of ketamine to increase striatal dopamine concentrations, consistent with the role of the NMDA receptor in modulating dopamine function.
Similar content being viewed by others

Main
Glutamate is the primary excitatory neurotransmitter in the brain, and there is an increasing amount of evidence for the role of glutamine receptors in neuropsychiatric disease (e.g., Javitt and Zukin 1991; Coyle 1987; Choi 1988; Ellison 1995). To further elucidate the role of the glutamate receptor system in behavior and neurochemical function, particularly in human subjects, the development of a pharmacologic probe is extremely important. The importance of a pharmacologic probe for glutamatergic function is underscored by the lack of a suitable glutamate receptor radiotracer for in vivo imaging (e.g., Blin et al. 1991). This would permit the development of methods to measure glutamate modulation of other neurotransmitter systems that may provide greater insight into the role of glutamate in normal function and in disease.
The “dissociative anesthetics” such as phencyclidine (PCP) and its derivative ketamine have been administered as a pharmacologic challenge to evaluate the role of glutamate receptor antagonism. These agents bind to a site within the N-methyl-D-aspartate (NMDA) receptor complex, a glutamate receptor subtype (Corssen and Domino 1966; Zukin and Zukin 1976; Vincent et al. 1979). At anesthetic doses (1.0 mg/kg and higher), ketamine has multiple neurochemical effects including the inhibition of monoamine transport and acetylcholinesterase and the blockade of sigma receptors (e.g., Smith et al. 1981; Oye et al. 1991; Cohen et al. 1974). The administration of subanesthetic doses of ketamine (0.1–0.5 mg/kg) has demonstrated a relatively selective effect in binding to the NMDA site (Javitt and Zukin 1991; Thomson et al. 1985), as there is a several-fold difference in the affinities of monoamine transporter and sigma binding sites compared with the NMDA binding site (e.g., Smith et al. 1981; Oye et al. 1991).
Administration of anesthetic doses of PCP to patients and of subanesthetic doses of PCP to normal volunteers altered in behavior in a manner that has been compared with psychotic symptoms (e.g.,Javitt and Zukin 1991; Krystal et al. 1994). Both PCP and ketamine administration exacerbated symptomatology in schizophrenic patients (e.g., Lahti et al. 1995a). Given the hypothesis that hyperdopaminergic function underlies psychotic symptoms, the psychotic-like behaviors observed after PCP and ketamine administration may be attributable to secondary effects on the dopamine system. This is consistent with the preclinical, neurochemical observations that NMDA receptor antagonists such as ketamine alter the firing rate of mesocortical and mesolimbic dopamine neurons and increase concentrations of extracellular dopamine in the striatum and prefrontal cortex (e.b., French 1994; McGeer et al. 1977; Moghaddam et al 1990; Verma and Moghaddam 1996).
Recently, dopamine concentrations and dopamine modulation has been measured in vivo using positron emission tomography (PET) and single photon emission computed tomography (SPECT) imaging modalities, radiotracers for the dopamine (D2) receptor and pharmacologic challenges that alter dopamine concentrations directly or indirectly through other neurotransmitter systems. Studies in non-human primates and human subjects have demonstrated that a pharmacologic increase in dopamine concentrations by administration of dopamine agonists and reuptake inhibitors such as d-amphetamine, GBR 12909, and methylphenidate decreased the striatal binding of D2 radiotracers (18F-N-methylspiroperidol. 123I-IBZM, 11C-raclopride), due to the increased competition between dopamine and for binding to the striatal D2 receptor (Dewey et al. 1991, 1993a; Innis et al. 1992; Volkow et al, 1994). Subsequent studies demonstrated that pharmacologic alterations of other systems known to modulate striatal dopamine (e.g., GABA, acetylcholine, serotonin) altered binding consistent with the preclinical data demonstrating that dopamine concentrations is modulated by these neurotransmitter systems (e.g., Dewey et al. 1992, 1993a, b, 1995; Smith et al. 1997). It is important to note these studies measured alterations in striatal D2 binding as the low densities of D2 receptors in extrastriatal regions cannot be detected with PET and the D2 radiotracers used in these studies. Given this limitation, it is of interest to assess cortical input to the striatum within the context of this PET experimental design. As the primary cortical input to the striatum is glutamatergic, this study was designed to develop a method to assess in vivo the functional integrity of cortical input to the striatum. The present study was undertaken to evaluate whether administration of subanesthetic doses of ketamine would decrease striatal 11C-raclopride binding in normal control subjects, consistent with an increase in dopamine concentrations.
MATERIALS AND METHODS
Subject Screening and Selection
Subjects underwent medical (physical and laboratory testing, toxicology screening, physical examination) and psychiatric evaluation and a magnetic resonance imaging (MRI) scan (Siemens 1.5T Magnatom Vision). Subjects were excluded based upon a history of or current significant medical, psychiatric, or neurologic disease, substance abuse, family history (first- or second-degree relatives) of psychiatric illness or substance abuse or use of prescription or over-the-counter medications with central nervous system (CNS) effects (e.g., antihistamines, cold medications) within the past month. Seven, right-handed male subjects were enrolled in the study, with a mean age of 31.0 ± 5.3 years (range 26–39 years). Subjects fasted after midnight and were given a standard breakfast 3 h before scanning. After a complete description of the study to the subjects, written informed consent was obtained. The experimental protocol and consent forms were approved by the Institutional Board of Research Associates at the New York University School of Medicine and the Human Studies Research Committee at Brookhaven National Laboratories.
PET Procedures
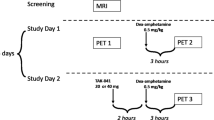
For each subject, two 11C-raclopride studies were performed on the same day. After performing a baseline 11C-raclopride scan, ketamine was administered (0.5 mg/kg, IV, infused over 20 min). Two pretreatment intervals were evaluated to determine when the maximal effects of ketamine on 11C-raclopride binding would be observed. In four subjects, the second 11C-raclopride scan was performed immediately after the ketamine infusion ended. In three subjects (subjects 3,4,5), 11C-raclopride was injected 10 min into the ketamine infusion, and the ketamine infusion continued for the initial 10 min of the 11C-raclopride study. As ketamine has a relatively short half-life in plasma, injecting the 11C-raclopride during the infusion could prolong potentially the pharmacologic effect. In the case of both time points, the timing of the second scan corresponded to the interval during which the maximal plasma concentration and the maximal behavioral effects of ketamine have been observed. The ketamine dose and administration method was consistent with that of the study reported by Krystal et al. (1994).
The 11C-raclopride studies were performed on the Computer Technology Inc. (CTI) 931-08/12 tomograph. Prior to scanning, catheters were placed in an antecubital vein for radiotracer injection and in a radial artery for blood sampling. The subject was positioned in the scanner using external lasers wet parallel to the cantho-meatal line and to the mid-line as marked on the head. Repositioning accuracy was insured by the use of the headholder and the positioning pins that secure the headholder to the cradle and the laser markers. The sensitivity of the machine was tested with a germanium phantom. After a blank scan, a 10-min transmission scan was obtained with a 68G source, to be used for attenuation correction of the emission scan.
11C-raclopride was synthesized by the reaction of carbon-11–labeled methyl iodide with nor-raclopride (Farde et al. 1986). 11C-raclopride was injected intravenously (5.09–13.41 mCi, Specific Activity 0.5–1.5 Ci/μmol). Scanning began immediately and lasted for 60 min. The scanning protocol involved 10 scans of 1 min, followed by 10 scans of 5 min. Continuous blood samples were obtained by an automated device (Ole Dich, Denmark) for the first 2 min after injection, at the peak of radioactivity in blood. For the remainder of the study, manual sampling was used. Selected plasma samples (1, 5, 10, 20, 30, 45, and 60 min) were analyzed for the presence of unchanged 11C-raclopride, as described previously (Volkow et al. 1993). Subject were scanned in a quiet, dimly lit room, with eyes open and ears unoccluded. The subjects were instructed that they would receive either ketamine or placebo before each study.
Plasma samples were obtained for analyses of the dopamine metabolite homovanillic acid (HVA), prolactin, and cortisol. The analytical methods and their coefficients of variation have been reported previously (Hollander et al. 1992). Plasma samples were obtained before and immediately after each of the two 11C-raclopride scans. The initial plasma sample (before the first 11C-raclopride study) was obtained approximately 30 min after placement of the arterial and venous catheters.
Data Analysis
Regions of interest (ROIs) were identified for the striatum (specific binding) and cerebellum (nonspecific binding). The ROIs for the striatum were drawn on four slices beginning at the head of the caudate and encompassing the caudate and putamen on the other three slices. Receptor availability was measured by a graphical analysis method designed for reversible systems (Logan et al. 1990). The date will be reported in terms of the distribution volume (DV), which is a linear function of the free receptor concentration. This method has been described in detail in earlier publications (e.g., Logan et al. 1990, 1995). The DV parameter is less sensitive to noise than the individual kinetic parameters, which often have large standard errors associated with their determination. In addition, the DV as derived is flow independent, since the flow terms appear in the numerator and denominator and thus, cancel out. The insensitivity of the DV analysis to changes in cerebral blood flow has been addressed experimentally in the analysis of two reversible tracers for two different neurotransmitter systems (11C-flumazenil, 11C-raclopride; Logan et al. 1995; Holthoff et al. 1991). To take into account changes in nonspecific binding, the data will be presented as the cerebellar DV (average of the two studies for each subject) subtracted from the striatal DV. As the magnitude of change of the cerebellar DV for each subject is within the range of the test-retest variability for the cerebellar DV (Volkow et al. 1993), the two cerebellar DV values were averaged to reduce the potential noise contributed by the cerebellar DV. In subtracting the cerebellum from the striatum, the striatal data is weighted more and the differences in receptor availability observed are due to a greater extent to changes in striatal binding. The use of the subtraction approach is supported by the observation that the striatum and cerebellum have comparable amounts of nonspecific binding (Farde et al. 1989) and that the administration of ketamine did not alter the cerebellar DV in a systematic manner (either increase or decrease) in excess of the test-retest variability for the cerebellum (Volkow et al. 1993). In considering the effect of ketamine on rCBF that might contribute to the changes in nonspecific binding, ketamine administration has been shown to alter rCBF primarily in limbic areas, but not in the striatum and cerebellum, the regions of interest for the 11C-raclopride studies (Lahti et al. 1995b). Thus, regionally selective changes in rCBF in the striatum and cerebellum are not likely to represent a major confounding factor.
Statistical Analysis
Repeated measures analysis of variance (ANOVA) was used to investigate the effects of ketamine on plasma levels of HVA, cortisol, prolactin, the rate of metabolism of 11C-raclopride and DV measurements for 11C-raclopride. If a significant repeated measures effect was obtained, post-hoc ANOVAs were performed to compare the individual time points.
RESULTS
As the subjects were being scanned during the maximal behavioral effects of the drug, they were questioned about their response to the drug after scanning. The subjects consistently reported feelings of anxiety, a sensation that they were floating and a general feeling of derealization that could not be easily described. All subjects reported that the maximal effect of the drug occurred within the first half hour of scanning.
The levels of cortisol, prolactin, and HVA in plasma are shown in Table 1. For the HVA and prolactin measures, the main effects of condition (placebo/drug) and time (pre/post-scan) and the condition by time interaction were not-significant (HVA: f[1,6] = 1.70, p > .1, f[1,6] = 0.19, p < .1 and f[1,6] = 0.00, p < .1, respectively: prolactin: (f[1,6] = 5.47, p > .05, f[1,6] = 3.35, p > .1 and f[1,6] = 1.95, p > .1, respectively). For cortisol, there was a significant effect of drug, but a nonsignicant effect of time and a drug by time interaction (f[1,6] = 8.96, p < .05, f[1,6] = 5.61, p > .05 and f[1,6] = 0.00, p < .1, respectively). To follow-up the significant effect of condition for cortisol, the sum of the baseline measures was compared with the two post-ketamine measures and a significant effect was obtained for the pre-scan 2 measure, but not for the post-scan 2 measure (f[1,6] = 12.43, p < .05, f[1,6] = 4.25, p < .1). Table 2 shows the percent of unchanged 11C-raclopride in arterial plasma over time for the two scans. Ketamine administration did not significantly alter the rate of metabolism of 11C-raclopride as the repeated measures ANOVA demonstrated a significant main effect of time, as expected, but not a significant effect of condition or a time by condition interaction (f[5,25] = 39.16, p < .001, f[1,6] = 0.29, p < .1 and f[1,5] = 0.24, p < .1, respectively).
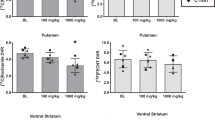
The DVs were decreased significantly in the striatum, but not in the cerebellum (f[1,6] = 11.86, p < .05, f[1,6] = 0.03, p > .1, respectively). A significant decrease in the subtracted DVs was observed. The cerebellar DVs subtracted from the striatal DVs (striatum − cerebellum) for the baseline and ketamine pretreatment condition were decreased significantly by ketamine administration (f[1,6] = 11.86, p < .05). The values for the subtracted DVs for the individual subjects are shown in Table 3 and Figure 1 . For the subtracted DVs, the magnitude of decrease varied across subjects. A separate set of analyses were performed to determine whether the magnitude of response differed as a function of timing of ketamine infusion (the subjects who received the second 11C-raclopride injection at the end of the ketamine infusion versus those who received the second 11C-raclopride injection in the middle of the ketamine infusion). No significant group differences were observed in the change in 11C-raclopride binding, cortisol, prolactin, or HVA. Finally, significant correlations were not obtained between the change in 11C-raclopride binding and the change in cortisol, prolactin, or HVA.

The effect of ketamine administration on 11C-raclopride binding. Ketamine administration resulted in a decrease in 11C-raclopride binding consistent with an increase in dopamine activity.
DISCUSSION
In the present study, ketamine administration decreased 11C-raclopride binding, consistent with an increase in dopamine concentrations. An alternative explanation is that the results obtained could be due to a shift of D2 receptors from a low to a high affinity state for dopamine. In addition, either a presynaptic mechanism (for example, increased dopamine release or decreased reuptake) or a postsynaptic mechanism (for example, increased affinity of receptors for synaptic dopamine) could account for the present findings.
For the subjects as a group, the change in 11C-raclopride binding was greater in magnitude than the test-retest variability of 11C-raclopride binding in studies performed on 2 consecutive days at the same time of day (Volkow et al. 1995). Whereas all of the subjects showed a decrease in binding, the magnitude of decrease varied across subjects. The variability was not accounted for by differences between subjects in age, baseline 11C-raclopride binding, behavioral response to ketamine infusion, baseline levels, or the magnitude of change in levels of cortisol, prolactin, or HVA. The differential timing of the 11C-raclopride injection relative to the ketamine infusion did not result in a systematic difference in response within the two groups of subjects. As the maximal effects of ketamine on behavior occurred during the PET scan, it was possible only to interview the subject after the PET scan. The behavioral effects of ketamine in these subjects were consistent with the effects reported in other studies designed specifically to measure behavioral effects. Whether the variability observed is inherent to the subjects in this study remains to be evaluated in a larger sample of subjects.
An increase in cortisol was observed after ketamine administration. Cortisol levels are also increased by pharmacologic challenges that increase dopamine concentrations directly and indirectly, such as d-amphetamine, methylphenidate, and fenfluramine (e.g., Smith et al. 1994, 1997; McBride et al. 1990). However, the neuroendocrine effects in the present study could also occur by a direct blockade of hypothalamic NMDA receptors (Gribkoff 1991). The lack of correlation between the change in 11C-raclopride binding and the change in the neuroendocrine measures was also observed in a previous 11C-raclopride study using fenfluramine challenge to measure serotonin modulation of dopamine concentrations (Smith et al. 1997). The lack of correlation may be due to the differential time course of the effects of ketamine on neuroendocrine function and dopamine concentrations.
The results of the present study are consistent with the findings of in vivo microdialysis studies that ketamine administration increased extracellular dopamine concentrations in the prefrontal cortex and the striatum, to a greater extent in prefrontal cortex (e.g., Verma and Moghaddam 1996). With regard to the mechanism underlying this effect, there is evidence that glutamate modulation of striatal dopamine occurs both at the level of the dopaminergic cell bodies of the ventral tegmental area (VTA) and substantia nigra (SN) and the level of the glutamatergic afferents of the prefrontal cortex that modulate concentrations in the VTA, SN, or in the striatum by providing excitatory input to the GABA interneurons (Monaghan and Cotman 1985; Carter 1982; Sesack and Pickel 1992). In evaluating these different mechanisms, a recent in vivo microdialysis study demonstrated that glutamatergic projections from the prefrontal cortex to the VTA were critical in controlling striatal dopamine release (Karreman and Moghaddam 1996). These microdialysis results, when considered with the effects of PCP or ketamine administration on cerebral metabolism and blood flow described below, support the hypothesis that the alterations in striatal dopamine concentrations observed in the present study may be a secondary consequence of altered cortical function. Radiotracers are currently in development to measure extrastriatal D1 and D2 receptors that may be sensitive to alterations in dopamine concentrations (e.g., 18F-desmethoxyfallypride, 11C-NNC 01-0112; Mukherjee et al. 1995; Halldin et al. 1994). When a suitable radiotracer is developed, it will be possible to measure glutamate modulation of dopamine in cortical and limbic areas in addition to the striatum.
The effects of acute ketamine or PCP administration differ from that of amphetamine administration (as reviewed by Javitt and Zukin 1991; Ellison 1995; Krystal et al. 1994); however, both drugs mimic different aspects of symptomatology in schizophrenia (negative versus positive symptoms). Both classes of drugs and have different regional effects on cerebral metabolism that resemble the metabolic alterations observed in schizophrenia. PCP administration to rats and ketamine administration to schizophrenic patients increased metabolism in sensory cortices, motor cortex, thalamus, and limbic areas (e.g., Gao et al. 1993; Weissman et al. 1987; Lahti et al. 1995b). D-amphetamine administration to schizophrenic patients decreased metabolism in frontal and temporal cortical areas (e.g., Wolkin et al. 1987). It is possible that deficits in the dopamine and glutamate systems may relate to different aspects of symptomatology and metabolic alterations in schizophrenia. The variability in symptomatology observed between patients may be related to different degrees of glutamate-dopamine imbalance.
In contrast, there are similarities between chronic use of PCP and ketamine and the stimulant agents (e.g., cocaine, amphetamine), and both classes of drugs are known for their abuse liability (for a review, see Siegel 1978). There are similarities in the pattern of abuse of both classes of agents in that “binge” intake patterns are observed, followed by a rebound depressive phase and in some individuals, persistent psychotic symptoms (Burns and Lerner 1981; Ellison 1995). As the release of dopamine is the primary neurochemical effect of cocaine and amphetamine and a secondary effect of PCP and ketamine, the addictive properties and associated symptoms of chronic use of these agents may be related to a common dopaminergic mechanism (e.g., Nomikos et al. 1990).
In conclusion, the administration of a subanesthetic dose of ketamine decreased 11C-raclopride binding, consistent with an increase in dopamine concentrations. These findings are consistent with the ability of NMDA receptor blockade to alter dopamine concentrations demonstrated by neurophysiologic and neurochemical studies. This experimental paradigm for measuring glutamate modulation of dopamine concentrations in vivo may be used to elucidate etiologic and therapeutic mechanisms in the psychotic and addictive disorders.
References
Blin J, Denis A, Yamaguchi T, Crouzel C, MacKenzie E, Baron J . (1991): PET studies of [18F]-methyl-MK-801, a potential NMDA receptor complex radioligand. Neurosci Lett 121: 183–186
Burns R, Lerner S . (1981): The effects of phencyclidine in man: A review. In Domino E (ed), PCP (Phencyclidine): Historical and Current Perspectives. Ann Arbor, MI, NPP Books, pp 449–469
Carter C . (1982): Topographic distribution of possible glutamatergic pathways from the frontal cortex to the striatum and substantia nigra in rats. Neuropsychopharmacology 21: 379–383
Choi D . (1988): Glutamate neurotoxicity and diseases of the nervous system. Neuron 1: 623–634
Cohen M, Chan S, Bhargava H, Trevor A . (1974): Inhibition of mammalian brain acetylcholinesterase by ketamine. Biochem Pharmacol 23: 1647–1652
Corssen G, Domino E . (1966): Dissociative anesthetics: Further pharmacologic studies and first clinical experience with the phencyclidine derivative Cl-581. Anesth Analg 45: 29–40
Coyle J . (1987): Kainate acid: Insights into excitatory mechanisms causing selective neuron degeneration. Ciba Found Symp 126: 186–203
Dewey SL, Logan J, Wolf A, Brodie J, Angrist B, Fowler J, Volkow N . (1991): Amphetamine induced decreases in 18F-N-methylspiroperidol binding in the baboon brain using positron emission tomography (PET). Synapse 7: 324–327
Dewey S, Smith G, Logan J, Alexoff D, King P, Pappas N, Ashby C, Brodie J . (1995): The serotonergic-dopaminergic interaction in vivo with positron emission tomography (PET) and microdialysis. J Neurosci 15: 821–829
Dewey SL, Smith GS, Logan J, Brodie JD, Fowler JS, Wolf AP . (1993a): Striatal binding of the PET ligand 11C-raclopride is altered by drugs that modify synaptic dopamine levels. Synapse 13: 350–356
Dewey SL, Smith G, Logan J, Simkowitz P, Brodie JD, Fowler JS, Volkow N, Wolf AP . (1993b): Effects of central cholinergic blockade on striatal dopamine release measured with positron emission tomography (PET) in normal human subjects. Proc Natl Acad Sci 90: 11816–11820
Dewey S, Smith G, Logan J, Brodie J, Yu D,: Ferrieri R, King P, MacGregor R, Martin T, Wolf A, Volkow N, Fowler J . (1992): GABAergic inhibition of endogenous dopamine release measured in vivo with 11C-raclopride and positron emission tomography. J Neurosci 12: 3773–3780
Ellison G . (1995): The N-methyl-D-aspartate antagonists phencyclidine, ketamine and dizocilpine as both behavioral and anatomical models of the dementias. Brain Res Rev 20: 250–267
Farde L, Hall H, Ehrin E, Sedvall G . (1986): Quantitative analysis of D2 receptor binding in the living human brain by PET. Science 231: 258–261
Farde L, Eriksson L, Blomquist G, Halldin C . (1989): Kinetic analysis of central [11C]-raclopride binding to D2-dopamine receptors studied by PET—A comparison to the equilibrium analysis. J Cereb Blood Flow Metab 9: 696–708
French E . (1994): Phencyclidine and the midbrain dopamine system: Electrophysiology and behavior. Neurotoxicol Teratol 16: 355–362
Gao X, Shirakawa F, Tamminga C . (1993): Delayed regional metabolic actions of phencyclidine. Eur J Pharmacol 241: 7–15
Gribkoff V . (1991): Electrophysiological evidence for N-methyl-D-asparate excitatory amino acid receptors in the rat supraoptic nucleus in vitro. Neurosci Lett 131: 260–262
Halldin C, Foged C, Karlsson P, Swahn CG, Andersen PH, Farde L . (1994): [C-11]NNC 112, a selective PET radioligand for examination of extrastriatal dopamine D1 receptors. J Nucl Med 35: 122P
Hollander E, DeCaria C, Nitescu A, Gully R, Suckow R, Cooper T, Gorman J, Klein D, Liebowitz M . (1992): Serotonergic function in obsessive-compulsive disorder. Arch Gen Psychiatry 49: 21–28
Holthoff V, Koeppe R, Frey K, Paradise A, Kuhl D . (1991): Differentiation of radioligand delivery and binding in the brain: validation of a two-compartment model for [11C]-flumazenil. J Cereb Blood Flow Metab 11: 745–752
Innis R, Mallison R, Al-Tikriti M, Hoffer M, Sybirska E, Seibyl J, Zoghbi S, Baldwin R, Laruelle M, Smith E, Charney D, Henninger G, Elsworth J, Roth R . (1992): Amphetamine stimulated dopamine release competes in vivo for [123I]-IBZM binding. Synapse 10: 177–184
Javitt D, Zukin S . (1991): Recent advances in the phencyclidine model of schizophrenia. Am J Psychiatry 148: 1301–1308
Karreman M, Moghaddam B . (1996): The prefrontal cortex regulates the basal release of dopamine in the limbic striatum: An effect mediated by ventral tegmental area. J Neurochem 66: 589–598
Krystal J, Karper L, Seibyl J, Freeman G, Delaney R, Bremner J, Heninger G, Bowers M, Charney D . (1994): Subanesthetic effects of the non-competitive NMDA antagonist, ketamine, in humans. Arch Gen Psychiatry 51: 199–214
Lahti A, Koffel B, LaPorte D, Tamminga C . (1995a): Subanesthetic doses of ketamine stimulate psychosis in schizophrenia. Neuropsychopharmacology 13: 9–19
Lahti A, Holcomb H, Medoff D, Tamminga C . (1995b): Ketamine activates psychosis and alters limbic blood flow in schizophrenia. Neuroreport 6: 869–872.
Logan J, Fowler J, Volkow N, Wolf A, Dewey S, Schleyer D, Macgregor R, Hitzemann R, Bendriem B, Gatley S, Christman D . (1990): Graphical analysis of reversible radioligand binding from time-activity measurements applied to N-11C-methyl-(−)-Cocaine PET studies in human subjects. J Neurochem 10: 740–747
Logan J, Volkow N, Fowler J, Wang G, Dewey S, Macgregor R, Schleyer D, Gatley S, Pappas N, King P . (1995): Effects of blood flow on 11C-raclopride binding in the brain: model simulations and kinetic analysis of PET data. J Cereb Blood Flow Metab 14: 995–1010
McBride P, Tierney H, DeMeo M, Chen J, Mann J . (1990): Effects of age and gender on CNS serotonergic responsivity in normal adults. Biol Psychiatry 27: 1143–1155
McGeer P, McGeer E, Scherrer U, Singh K . (1977): A glutamatergic corticostriatal path? Brain Res 128: 369–373
Moghaddam B, Gruen R, Roth R, Bunney B, Adams R . (1990): Effects of l-glutamate on the release of striatal dopamine: In vivo dialysis and electrochemical studies. Brain Res 518: 55–60
Monaghan D, Cotman C . (1985): Distribution of N-methyl-D-aspartate sensitive L-[3H]-glutamate binding sites in the rat brain. J Neurosci 5: 2909–2919
Mukherjee J, Yang Z, Brown T, Roemer J, Cooper M . (1996): 18F-desmethoxyfallypride: A fluorine-18 labeled radiotracer with properties similar to carbon-11 raclopride for PET imaging studies of dopamine D2 receptors. Life Sci 59: 669–678
Nomikos G, Damsma G, Wenkstern D, Fibiger H . (1990): In vivo characterization of locally applied dopamine uptake inhibitors by striatal microdialysis. Synapse 6: 106–112
Oye I, Hustveit O, Moberg E, Pausen O, Skoglund L . (1991): The chiral forms of ketamine as probes for NMDA receptor function in humans. In Kamewyama T, Nabeshima T, Domino E (eds), NMDA Receptor Related Agents: Biochemistry, Pharmacology and Behavior. Ann Arbor, MI, NPP Books, pp 381–389
Sesack S, Pickel V . (1992): Prefrontal cortical afferents in the rat synapse on unlabeled neuronal targets of catecholamine terminals in the nucleus accumbens septi and on dopamine neurons in the ventral tegmental area. J Comp Neurol 320: 145–160
Siegel R . (1978): Phencyclidine and ketamine intoxication: A study of four populations of recreational users. In Petersen R, Stillman R (eds), Phencyclidine (PCP) Abuse: An Appraisal. NIDA Research Monograph 21, Rockville, MD, National Institute on Drug Abuse, pp 119–147
Smith D, Azzaro A, Zaldivar S, Palmer S, Lee H . (1981): Properties of the optical isomers and metabolites of ketamine on the high affinity transport and catabolism of monoamines. Neuropharmacology 20: 391–396
Smith G, Dewey S, Logan J, Brodie J, Simkowitz P, Vitkun S, Volkow N, Alexoff D, Martin T, Jenkins D, Shea C, Fowler J, Wolf A . (1994): Dopamine modulation of acetylcholine activity measured with positron emission tomography (PET) and 11C-benztropine in the human brain. Soc Neurosci Abs 20: 12
Smith G, Dewey SL, Brodie JD, Logan J, Vitkun S, Simkowitz P, Alexoff D, Volkow N . (1997): Serotonin modulation of dopamine measured with positron emission tomography (PET) and 11C-raclopride in normal human subjects. Am J Psychiatry 154: 490–496
Thomson A, West D, Lodge D . (1985): An n-methylasparate receptor-mediated synapse in rat cerebral cortex: A site of action of ketamine? Nature 313: 479–481
Ulas J, Cotman C . (1993): Excitatory amino acid receptors in schizophrenia. Schizophr Bull 19: 105–117
Verma A, Moghaddam B . (1996): NMDA receptor antagonists impair prefrontal cortex function as assessed via spatial delayed alternation performance in rats: Modulation by dopamine. J Neurosci 16: 373–379
Vincent J, Kartalovski B, Geneste P, Kamenka J, Lazdunski M . (1979): Interaction of phencyclidine (“angel dust”) with a specific receptor in rat brain membrane. Proc Natl Acad Sci 76: 4678–4682
Volkow N, Fowler J, Wang G-J, Dewey S, Schleyer D, MacGregor R, Logan J, Alexoff D, Shea C, Hitzemann R, Angrist B, Wolf A . (1993): Reproducibility of repeated measures of Carbon-11 raclopride binding in the human brain. J Nucl Med 34: 609–613
Volkow N, Wang G-J, Fowler J, Logan J, Schleyer D, Hitzemann R, Liegerman J, Angrist B, Pappas N, MacGregor R, Burr G, Cooper T, Wolf A . (1994): Imaging dopamine release in the human brain. Synapse 16: 255–262
Weissman A, Dam M, London E . (1987): Alterations in local cerebral glucose utilization induced by phencyclidine. Brain Res 435: 29–40
Wolkin A, Angrist B, Wolf A, Brodie J, Wokkin B, Jaeger J, Cancro R, Rotrosen J . (1987): Effects of amphetamine on local cerebral metabolism in normal and schizophrenic subjects as determined by positron emission tomography. Psychopharmacology 92: 241–246
Zukin S, Zukin R . (1976): Specific [3H]phencyclidine binding in the rat central nervous system. Proc Natl Acad Sci 76: 5372–5376
Acknowledgements
This study was supported in part by Department of Energy/Office of Environmental Research; Grants NS15638 and NS15380 from National Institute of Neurological Disorders and Stroke; Grants MH49936, MH47277 and MH49165 from the National Institute for Mental Health; Grant RR-00096 from the National Center for Research Resources; Young Investigator Awards (to GSS and PS) and Established Investigator Awards (to SLD and JDB) from the National Alliance for Research in Schizophrenia and Depression (NARSAD) and a grant from the Whitehead Foundation (to GSS). The following individuals are gratefully acknowledged for their contribution to the studies: Noelwah Netusil and Theodore Johnson for patient care, Donald Warner and Naomi Pappas for PET operations, Colleen Shea, Thomas P. Martin, and Darren Jenkins for radiopharmaceutical preparation, and Clarence Barrett and Robert Carciello for cyclotron operations.
This manuscript is dedicated to the memory of Dr. Elsa J. Bartlett.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Smith, G., Schloesser, R., Brodie, J. et al. Glutamate Modulation of Dopamine Measured in Vivo with Positron Emission Tomography (PET) and 11C-Raclopride in Normal Human Subjects. Neuropsychopharmacol 18, 18–25 (1998). https://doi.org/10.1016/S0893-133X(97)00092-4
Received:
Revised:
Accepted:
Issue Date:
DOI: https://doi.org/10.1016/S0893-133X(97)00092-4
Keywords
This article is cited by
-
Are there links between Alzheimer’s disease and ADHD? The efficacy of acetylcholinesterase inhibitors and NMDA receptor antagonists in controlling ADHD symptoms: a systematic review
Middle East Current Psychiatry (2024)
-
Glutamate-Weighted CEST Contrast After Removal of Magnetization Transfer Effect in Human Brain and Rat Brain with Tumor
Molecular Imaging and Biology (2020)
-
The effects of ketamine on dopaminergic function: meta-analysis and review of the implications for neuropsychiatric disorders
Molecular Psychiatry (2018)
-
Potential Effect of Prolonged Sevoflurane Anesthesia on the Kinetics of [11C]Raclopride in Non-human Primates
Molecular Imaging and Biology (2018)
