









« Prev Next »

GPCR
G-protein-coupled receptors (GPCRs) are the largest and most diverse group of membrane receptors in eukaryotes. These cell surface receptors act like an inbox for messages in the form of light energy, peptides, lipids, sugars, and proteins. Such messages inform cells about the presence or absence of life-sustaining light or nutrients in their environment, or they convey information sent by other cells.
GPCRs play a role in an incredible array of functions in the human body, and increased understanding of these receptors has greatly affected modern medicine. In fact, researchers estimate that between one-third and one-half of all marketed drugs act by binding to GPCRs.
What Do GPCRs Look Like?
GPCRs bind a tremendous variety of signaling molecules, yet they share a common architecture that has been conserved over the course of evolution. Many present-day eukaryotes — including animals, plants, fungi, and protozoa — rely on these receptors to receive information from their environment. For example, simple eukaryotes such as yeast have GPCRs that sense glucose and mating factors. Not surprisingly, GPCRs are involved in considerably more functions in multicellular organisms. Humans alone have nearly 1,000 different GPCRs, and each one is highly specific to a particular signal.
GPCRs consist of a single polypeptide that is folded into a globular shape and embedded in a cell's plasma membrane. Seven segments of this molecule span the entire width of the membrane — explaining why GPCRs are sometimes called seven-transmembrane receptors — and the intervening portions loop both inside and outside the cell. The extracellular loops form part of the pockets at which signaling molecules bind to the GPCR.
What Do GPCRs Do?
As their name implies, GPCRs interact with G proteins in the plasma membrane. When an external signaling molecule binds to a GPCR, it causes a conformational change in the GPCR. This change then triggers the interaction between the GPCR and a nearby G protein.
G proteins are specialized proteins with the ability to bind the nucleotides guanosine triphosphate (GTP) and guanosine diphosphate (GDP). Some G proteins, such as the signaling protein Ras, are small proteins with a single subunit. However, the G proteins that associate with GPCRs are heterotrimeric, meaning they have three different subunits: an alpha subunit, a beta subunit, and a gamma subunit. Two of these subunits — alpha and gamma — are attached to the plasma membrane by lipid anchors (Figure 1).

Figure 1: Activation of the G alpha subunit of a G-protein-coupled receptor
In unstimulated cells, the state of G alpha (orange circles) is defined by its interaction with GDP, G beta-gamma (purple circles), and a G-protein-coupled receptor (GPCR; light green loops). Upon receptor stimulation by a ligand called an agonist, the state of the receptor changes. G alpha dissociates from the receptor and G beta-gamma, and GTP is exchanged for the bound GDP, which leads to G alpha activation. G alpha then goes on to activate other molecules in the cell.
© 2002 Nature Publishing Group Li, J. et al. The Molecule Pages database. Nature 420, 716-717 (2002). All rights reserved. 
A G protein alpha subunit binds either GTP or GDP depending on whether the protein is active (GTP) or inactive (GDP). In the absence of a signal, GDP attaches to the alpha subunit, and the entire G protein-GDP complex binds to a nearby GPCR. This arrangement persists until a signaling molecule joins with the GPCR. At this point, a change in the conformation of the GPCR activates the G protein, and GTP physically replaces the GDP bound to the alpha subunit. As a result, the G protein subunits dissociate into two parts: the GTP-bound alpha subunit and a beta-gamma dimer. Both parts remain anchored to the plasma membrane, but they are no longer bound to the GPCR, so they can now diffuse laterally to interact with other membrane proteins. G proteins remain active as long as their alpha subunits are joined with GTP. However, when this GTP is hydrolyzed back to GDP, the subunits once again assume the form of an inactive heterotrimer, and the entire G protein reassociates with the now-inactive GPCR. In this way, G proteins work like a switch — turned on or off by signal-receptor interactions on the cell's surface.
Whenever a G protein is active, both its GTP-bound alpha subunit and its beta-gamma dimer can relay messages in the cell by interacting with other membrane proteins involved in signal transduction. Specific targets for activated G proteins include various enzymes that produce second messengers, as well as certain ion channels that allow ions to act as second messengers. Some G proteins stimulate the activity of these targets, whereas others are inhibitory. Vertebrate genomes contain multiple genes that encode the alpha, beta, and gamma subunits of G proteins. The many different subunits encoded by these genes combine in multiple ways to produce a diverse family of G proteins (Figure 2).

Figure 2: The relationships of G proteins to the plasma membrane
In this diagram of G-protein-coupled receptor activation, the alpha, beta, and gamma subunits are shown with distinct relationships to the plasma membrane. After exchange of GDP with GTP on the alpha subunit, both the alpha subunit and the beta-gamma complex may interact with other molecules to promote signaling cascades. Note that both the alpha subunit and the beta-gamma complex remain tethered to the plasma membrane while they are activated. These activated subunits can act on ion channels in the cell membrane, as well as cellular enzymes and second messenger molecules that travel around the cell.
© 2010 Nature Education All rights reserved.
What Second Messengers Do GPCR Signals Trigger in Cells?
Activation of a single G protein can affect the production of hundreds or even thousands of second messenger molecules (Figure 3). (Recall that second messengers — such as cyclic AMP [cAMP], diacylglycerol [DAG], and inositol 1, 4, 5-triphosphate [IP3] — are small molecules that initiate and coordinate intracellular signaling pathways.) One especially common target of activated G proteins is adenylyl cyclase, a membrane-associated enzyme that, when activated by the GTP-bound alpha subunit, catalyzes synthesis of the second messenger cAMP from molecules of ATP. In humans, cAMP is involved in responses to sensory input, hormones, and nerve transmission, among others.
Phospholipase C is another common target of activated G proteins. This membrane-associated enzyme catalyzes the synthesis of not one, but two second messengers — DAG and IP3 — from the membrane lipid phosphatidyl inositol. This particular pathway is critical to a wide variety of human bodily processes. For instance, thrombin receptors in platelets use this pathway to promote blood clotting.
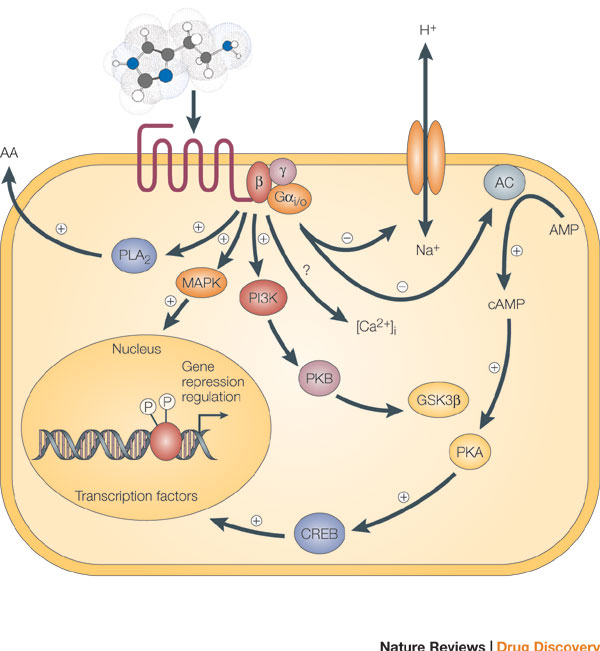
Figure 3: Signaling cascades within a cell can interact to affect multiple molecules in the cell, leading to secretion of substances from the cell, ion channel opening, and transcription.
Binding of an agonist to the seven-transmembrane G-protein-coupled receptor in the plasma membrane activates a pathway that involves G proteins as well as cAMP-related pathways that modulate cellular signaling. In this example, the activated G alpha (Gαi/0) proteins inhibit (-) adenylyl cyclase (AC, on the right), the enzyme that induces formation of cAMP, which in turn results in the activation of protein kinase A (PKA). This in turn activates a molecule called cAMP-responsive element-binding protein (CREB), which modulates gene transcription. The activated G alpha proteins can also have a variety of other effects, shown at the left. These effects include activating the mitogen-activated protein kinase (MAPK) and phosphatidylinositol 3-kinase (PI3K) pathways. Activation of the enzyme phospholipase A2 (PLA2) may also occur, which induces the release of arachidonic acid (AA), as well as inhibition of the Na+/H+ exchanger in the plasma membrane, and the lowering of intracellular Ca2+ levels (exact mechanism unknown, ?). Subsequent activation of the MAPK and PI3K pathways results in the phosphorylation of extracellular signal-regulated kinases (ERKs) and protein kinase B (PKB), respectively. Activated PKB will subsequently phosphorylate and thereby inhibit the action of glycogen synthase kinase 3beta (GSK3beta), a major kinase in the brain.
© 2005 Nature Publishing Group Leurs, R. et al. The histamine H3 receptor: from gene cloning to H3 receptor drugs. Nature Reviews Drug Discovery 4, 107-120 (2005). All rights reserved.
Conclusion
GPCRs
are a large family of cell surface receptors that respond to a variety
of
external signals. Binding of a signaling molecule to a GPCR results in G
protein activation, which in turn triggers the production of any number
of
second messengers. Through this sequence of events, GPCRs help regulate
an
incredible range of bodily functions, from sensation to growth to
hormone
responses.
eBooks
This page appears in the following eBook











