 intestine, (b) esophagus, and (c) stomach are shown. Through asymmetric division, quiescent stem cells (d) probably give rise to more rapidly dividing active stem cells, which then produce progenitor cells while losing their multipotency and ability to proliferate. All these progeny cells have defined positions in the different organs. To maintain its function and continue to produce new stem cells, a stem cell can also divide into and produce more stem cells at the same position (symmetric division).
", "
true
", "
All rights reserved.
", '
700
', '
825
', '
http://www.nature.com
');)










« Prev Next »

Cell Differentiation and Tissue
Within multicellular organisms, tissues are organized communities of cells that work together to carry out a specific function. The exact role of a tissue in an organism depends on what types of cells it contains. For example, the endothelial tissue that lines the human gastrointestinal tract consists of several cell types. Some of these cells absorb nutrients from the digestive contents, whereas others (called goblet cells) secrete a lubricating mucus that helps the contents travel smoothly.
However, the multiple cell types within a tissue don't just have different functions. They also have different transcriptional programs and may well divide at different rates. Proper regulation of these rates is essential to tissue maintenance and repair. The spatial organization of the cells that form a tissue is also central to the tissue's function and survival. This organization depends in part on polarity, or the orientation of particular cells in their place. Of course, external signals from neighboring cells or from the extracellular matrix are also important influences on the arrangement of cells in a tissue.
What Is the Source of New Cells for Tissues?
Without cell division, long-term tissue survival would be impossible. Inside every tissue, cells are constantly replenishing themselves through the process of division, although the rate of turnover may vary widely between different cell types in the same tissue. For example, in adult mammal brains, neurons rarely divide. However, glial cells in the brain continue to divide throughout a mammal's adult life. Mammalian epithelial cells also turn over regularly, typically every few days.
Neurons are not the only cells that lose their ability to divide as they mature. In fact, many differentiated cells lose this ability. To help counteract this loss, tissues maintain stem cells to serve as a reservoir of undifferentiated cells. Stem cells typically have the capacity to mature into many different cell types. Transcription factors — proteins that regulate which genes are transcribed in a cell — appear to be essential to determining the pathway particular stem cells take as they differentiate. For example, both intestinal absorptive cells and goblet cells arise from the same stem cell population, but divergent transcriptional programs cause them to mature into dramatically different cells (Figure 1).
Whenever stem cells are called upon to generate a particular type of cell, they undergo an asymmetric cell division. With asymmetric division, each of the two resulting daughter cells has its own unique life course. In this case, one of the daughter cells has a finite capacity for cell division and begins to differentiate, whereas the other daughter cell remains a stem cell with unlimited proliferative ability.
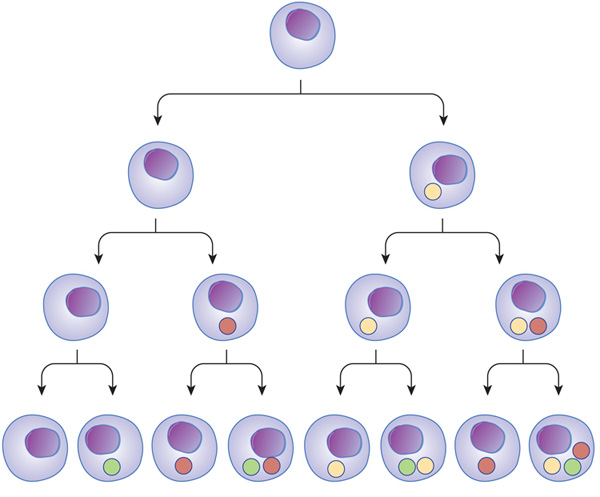
Figure 1: Transcriptional regulators can act at different stages, and in different combinations, through the path of cell development and differentiation.
Transcription factors can turn on at different times during cell differentiation. As cells mature and go through different stages (arrows), transcription factors (colored balls) can act on gene expression and change the cell in different ways. This change affects the next generation of cells derived from that cell. In subsequent generations, it is the combination of different transcription factors that can ultimately determine cell type.
© 2010 Nature Education All rights reserved. 
How Do Non-Growing Tissues Maintain Themselves?
Although most of the tissues in adult organisms maintain a constant size, the cells that make up these tissues are constantly turning over. Therefore, in order for a particular tissue to stay the same size, its rates of cell death and cell division must remain in balance.
A variety of factors can trigger cell death in a tissue. For example, the process of apoptosis, or programmed cell death, selectively removes damaged cells — including those with DNA damage or defective mitochondria. During apoptosis, cellular proteases and nucleases are activated, and cells self-destruct. Cells also monitor the survival factors and negative signals they receive from other cells before initiating programmed cell death. Once apoptosis begins, it proceeds quickly, leaving behind small fragments with recognizable bits of the nuclear material. Specialized cells then rapidly ingest and degrade these fragments, making evidence of apoptosis difficult to detect.
What Cellular Components Support Tissue Structure?
 intestine, (b) esophagus, and (c) stomach are shown. Through asymmetric division, quiescent stem cells (d) probably give rise to more rapidly dividing active stem cells, which then produce progenitor cells while losing their multipotency and ability to proliferate. All these progeny cells have defined positions in the different organs. To maintain its function and continue to produce new stem cells, a stem cell can also divide into and produce more stem cells at the same position (symmetric division).", "true", "All rights reserved.", '700', '825', 'http://www.nature.com');)
© 2009 Nature Publishing Group Quante, M. & Wang, T. C. Stem cells in gastroenterology and hepatology. Nature Reviews Gastroenterology & Hepatology 6, 724-737 (2009) doi:10.1038/nrgastro.2009.195. All rights reserved.
The gut endothelium — actually an epithelium that lines the inner surface of the digestive tract — is an excellent example of these structures at work. Here, tight junctions between cells form a seal that prevents even small molecules and ions from moving across the endothelium. As a result, the endothelial cells themselves are responsible for determining which molecules pass from the gut lumen into the surrounding tissues. Meanwhile, adherens junctions based on transmembrane cadherin proteins provide mechanical support to the endothelium. These junctions are reinforced by attachment to an extensive array of actin filaments that underlie the apical — or lumen-facing — membrane. These organized collections of actin filaments also extend into the microvilli, which are the tiny fingerlike projections that protrude from the apical membrane into the gut lumen and increase the surface area available for nutrient absorption. Additional mechanical support comes from desmosomes, which appear as plaque-like structures under the cell membrane, attached to intermediate filaments. In fact, desmosome-intermediate filament networks extend across multiple cells, giving the endothelium sheetlike properties. In addition, within the gut there are stem cells that guarantee a steady supply of new cells that contribute to the multiple cell types necessary for this complex structure to function properly (Figure 2).
How Does the Extracellular Matrix Support Tissue Structure?
The extracellular matrix (ECM) is also critical to tissue structure, because it provides attachment sites for cells and relays information about the spatial position of a cell. The ECM consists of a mixture of proteins and polysaccharides produced by the endoplasmic reticula and Golgi apparatuses of nearby cells. Once synthesized, these molecules move to the appropriate side of the cell — such as the basal or apical face — where they are secreted. Final organization of the ECM then takes place outside the cell.
To understand how the ECM works, consider the two very different sides of the gut endothelium. One side of this tissue faces the lumen, where it comes in contact with digested food. The other side attaches to a specialized ECM support structure called the basal lamina. The basal lamina is composed of collagen and laminin proteins, as well as various other macromolecules. On this side of the endothelium, adhesive junctions attach cells to the ECM. Transmembrane integrin proteins in the junctions bind components of the ECM and recruit signaling proteins to their cytoplasmic sides. From there, the signals travel to the nucleus of each cell.
Conclusion
Tissues are communities of cells that have functions beyond what any single cell type could accomplish. Healthy tissues require the proper mix of cells, and the cells within them must be oriented correctly and dividing at an appropriate rate. In order to coordinate their function, organization, and rates of death and division, the cells in a tissue are constantly processing and responding to signals from one another and from the ECM around them.
eBooks
This page appears in the following eBook












 intestine, (b) esophagus, and (c) stomach are shown. Through asymmetric division, quiescent stem cells (d) probably give rise to more rapidly dividing active stem cells, which then produce progenitor cells while losing their multipotency and ability to proliferate. All these progeny cells have defined positions in the different organs. To maintain its function and continue to produce new stem cells, a stem cell can also divide into and produce more stem cells at the same position (symmetric division).
", "
true
", "
All rights reserved.
", '
700
', '
825
', '
http://www.nature.com
');){kind=link}