





















« Prev Next »

Why Do Primates Communicate?
Like other animals, primates communicate to satisfy their biological and social needs, such as avoiding predators, interacting with other group members, or maintaining cohesion during travel. To this end, they use a range of different signals, many of which have directly evolved as ritualised abbreviations of more basic behavioural or physiological processes. For example, chimpanzees sometimes react with pilo-erection (bristling of hair) during conflicts, which makes them appear bigger and more dangerous and conveys their willingness to escalate (van Hooff 1973). Communication signals have thus evolved partly to be psychologically effective on receivers (Guilford & Dawkins 1991).
Is Primate Communication More Complex Compared to Other Groups of Animals?
Most primates live in groups in which members know each other individually and maintain multifaceted social relations; factors which are thought to favour the evolution of advanced communication skills (McComb & Semple 2005). However, other animals with complex social behaviour, such as dolphins, also show sophisticated communication skills, suggesting that complex communication is not limited to primates (Janik 2009).
Do Primates ‘Cry Wolf'?
Monkeys sometimes produce terrestrial predator alarms when competing over food, even though no predator is around. As a result, other group members run to safety, which then gives the caller a foraging advantage (Wheeler 2009). In general, however, primates rarely produce such dishonest signals, or ‘cry wolf'. Why is dishonest signalling not more common? One solution has been given by Zahavi's (1975) ‘handicap principle', which states that receivers will only attend to signals that are difficult to fake by low-quality or poorly motivated individuals. It has also been argued that, in primates, individuals know and need each other and thus gain little from deception (Silk et al. 2000). Moreover, primates can learn to ignore unreliable signallers (Cheney & Seyfarth 1988), suggesting that ‘reputation' acts as a further safeguard against dishonest signalling. Honest signalling prevails because of sceptical receivers.
Channels of Primate Communication
Primate communication takes place in all major modalities. Olfaction is one of the least researched modalities, partly because it is difficult to measure and manipulate olfactory cues, especially in the wild. Nevertheless, probably all primates secrete scents that influence others. An interesting human example is women apparently influencing each other's ovulation through odourless cues (Stern & McClintock 1998). Another remarkable example is the ‘stink fights' of male ring-tailed lemurs. During conflicts, males rub their tails across their wrist and chest glands before waving them at each other (Jolly 1966). Generally, olfactory cues play important roles in stating claims over resources and displaying individual characteristics, such as reproductive state, social rank, immuno-compatibility, and other genetic traits (Wedekind et al. 1995). One difficulty with research on olfactory communication is that it is often unclear whether scent-bearing substances are actively and strategically released into the environment, or whether they are mere by-products of general metabolic processes. Active scent marking and self-anointment (applying scent-bearing substances onto a substrate or body) are notable exceptions, but in many cases it is unclear whether olfactory cues qualify as proper communication signals.
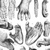
In the visual domain, primates use a range of facial displays and body part movements as communication signals, sometimes combined with tactile components. Gorilla chest beating and Rhesus monkeys bared-teeth displays are examples of how different species express social rank with visual signals. Current research has focused much on gestures, which are interesting because of their partly flexible, partly species-specific use in a variety of social contexts. Gestures have been studied mainly in great apes, where considerable variation between individuals and groups has been found. Whether some of this variation is socially learned and thus potentially ‘cultural' is still an unresolved question. A more established finding is that, during gesturing, apes take into account each other's attention and deploy their signals accordingly (Call & Tomasello 2007; Figure 1). Interestingly, however, there is almost no evidence that primate gestures, or combinations thereof, carry symbolic meaning by referring to external entities (Cartmill & Byrne 2010). Instead, they appear to function primarily to facilitate ongoing social interactions, to bond with others, or to persuade others to behave in a desired way.

Figure 1: Gestural communication.
Gestural communication in the Sonso chimpanzee community of Budongo Forest, Uganda. The young infant ‘Cathy’ reaches out to the older infant ‘Zak’, who interprets and accepts this gesture as a play invitation. ‘Zak’ responds with a ‘play-face’ and both begin to wrestle. Primate gestures have been defined as 'discrete, mechanically ineffective physical movements of the body observed during periods of intentional communication' (Hobaiter & Byrne 2011).
© 2012 Nature Education Courtesy of C. Hobaiter. All rights reserved. 
For many primates, vocalizations are the main channel of communication. Vocal repertoires tend to be species-specific, indicating that they develop under strong genetic control. Humans also possess a specific repertoire of such context-specific calls but, in addition, they also possess extensive control over vocal production, an ability that develops early and is crucial for the acquisition of speech. Such high degrees of vocal control is not seen in other primates and one interesting hypothesis is that it is the product of relatively recent genetic changes during human evolution (Enard et al. 2002). If this is correct then our hominid ancestors must have relied on a primate-like communication system.
In sum, primates communicate using all major modalities. Olfactory communication is poorly researched but is probably widespread, mostly inflexible and contextually confined to basic biological functions. Within the visual modality, gestures are somewhat of an exception because of their flexibility and socially targeted use. Finally, vocal communication is based on species-specific repertoires, with some flexibility in use but little in structure.
The Origins of Syntax and Meaning
What are the evolutionary origins of syntax? A persistent claim has been that human languages are the product of a genetically endowed, universal grammatical deep structure that — amongst other things — enables children to acquire language without much help (Chomsky 2005). Although extremely influential, it has been impossible to find convincing empirical support for this idea, for example by identifying grammatical features that are shared amongst all the world's languages (Evans & Levinson 2009). Nevertheless, humans possess the capacity for syntactic communication, so what are the biological roots of this ability? At a basic level, various primates produce utterances that consist of combinations of calls that can be meaningful to others. For example, male putty-nosed monkeys produce series of two basic loud calls to external threats, such as eagles and leopards. In addition, males sometimes combine the two calls into a unique sequence, which conveys its own meaning (‘move away'), regardless of the nature of the external disturbance (Arnold & Zuberbühler 2006; Figure 2). Similarly, adult male Campbell's monkeys produce different loud call types, which they combine into structurally unique context-specific sequences (Ouattara et al. 2009). Meaning at the sequence level has also been found in ape vocalisations, including gibbons and more recently bonobos (Clay & Zuberbühler 2011), suggesting that syntactically organised sequences are widespread in primate communication. In contrast to human language, there is no evidence that primates use such call combinations in a generative and creative way to ‘make infinite use of finite means' (von Humboldt 1836).
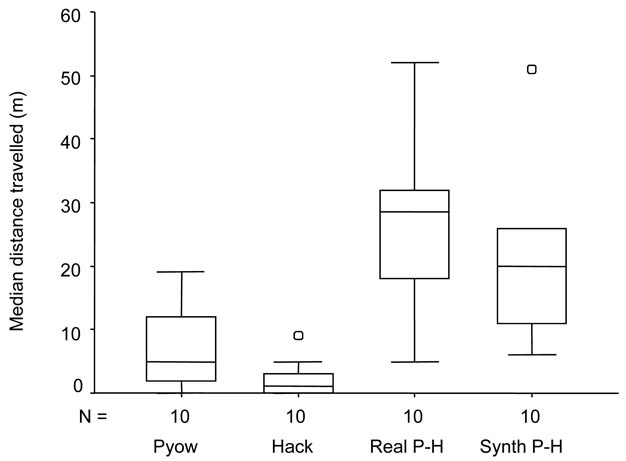
Figure 2: Putty-nosed monkeys combine two call types.
Putty-nosed monkeys combine two call types (‘pyows’ and ‘hacks’) into meaningful sequences. Females travelled further after hearing playbacks of ‘pyow-hack’ combinations of their own male (both real and synthetically composed) compared to his ‘pyow’ series (indicating a disturbance on the ground) or ‘hack’ series (indicating the presence of a crowned eagle).
© 2012 Nature Education Data from Arnold & Zuberbühler (2008). All rights reserved.
Another feature of human communication is that speech acts can be symbolic by referring to both mental entities and events in the outside world. Furthermore, humans make ample use of indexical and iconic signals, such as pointing or pantomiming, and from an early age, infants understand that symbolic and indexical signals complement each other in their capacity to refer to external objects (Gliga & Csibra 2009). Although laboratory research has shown that great apes can learn a considerable number of arbitrary symbols and follow indexical signals, there is little evidence that they use such acquired skills in creative ways or when communicating with each other (e.g., Savage-Rumbaugh et al. 1986).
In natural communication, however, some primate signals function in symbolic ways, the classic example being the vervet monkey alarm calls. These monkeys produce acoustically distinct alarm calls in response to different predators, which in turn elicit adaptive responses in listeners, as if the corresponding predator were present (Seyfarth et al. 1980). As there is nothing in the calls' acoustic features that would allow receivers to infer the cause of calling, the calls function like symbols. By the same logic, however, honeybee dance language also qualifies as symbolic, suggesting that some animals may not appreciate the symbolic nature of their signals but operate with cognitively less complicated mechanisms. For primate communication, several studies have sought to address this point. In one experiment, Diana monkeys were exposed to pairs of playback stimuli that simulated the presence of a predator — either an eagle or a leopard. In the key experimental condition, the predator vocalization was replaced with another monkey's corresponding alarm calls. Subjects responded in ways that suggested that they were guided by a mental representation of the corresponding event, independent of the acoustic features of the stimuli presented to them (Zuberbühler et al. 1999; Figure 3). This and other work suggest that primates have some understanding that calls can refer to external events, which they mentally represent even if they cannot see them.
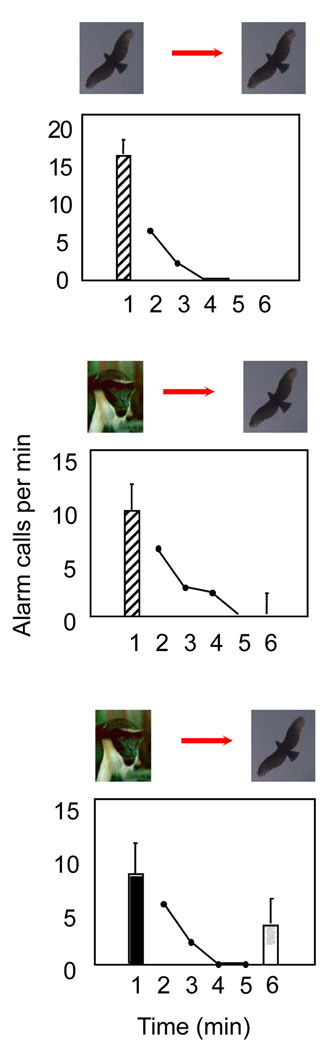
Figure 3: Diana monkeys understand the predator-alarm calls of other monkeys.
Diana monkeys understand the predator-alarm calls of other monkeys, (within and across species) and perceive them as representations of the corresponding predator class. Top panel: baseline - eagle alarm calls by female Diana monkeys given in response to playback of eagle shrieks; intermediate panel: test condition – eagle alarm calls given by female Diana monkeys in response to a male’s eagle alarm calls followed by eagle shrieks; bottom panel: control condition – leopard alarm calls given by female Diana monkeys in response a male’s leopard alarm calls followed by eagle shrieks. Females only ceased to respond to probe stimuli if they were primed with a semantically corresponding stimulus (baseline and test conditions).
© 2012 Nature Education Data from Zuberbühler et al. (1999). All rights reserved.
In sum, although primates sometimes combine different call types into complex utterances that are meaningful to others, it is still unclear how this behaviour is related to syntax in human language. One key difference appears to be that human languages, and their grammars, are socially learned, while there is no such evidence for primate call sequences. The difference between human and primate communication is less obvious in the domain of call comprehension. Both human and non-human primates extract meaning by relating arbitrary sound structures to real-world events and their mental representations.
Do Primates Actively Inform Each Other?
Another contentious issue in the evolution of language debate is whether primates communicate to actively inform each other. Humans tend to interpret communicative acts as an underlying intention to be informative (Grice 1969). Thus, meaning does not solely emerge from the speech signal, but also from the ‘common ground' shared by the interlocutors. There is currently no good evidence that primate communication operates this way, although some key abilities seem to be present. For example, primates can predict the impact of their signals on others (e.g., Hopkins et al. 2007). Similarly, both apes and monkeys have demonstrated considerable audience awareness that suggest that they have some understanding of the effects of their calls and gestures (e.g., Wich & de Vries 2006, Laporte & Zuberbühler 2010, Genty et al. 2009; Crockford et al. 2012; Figure 4).
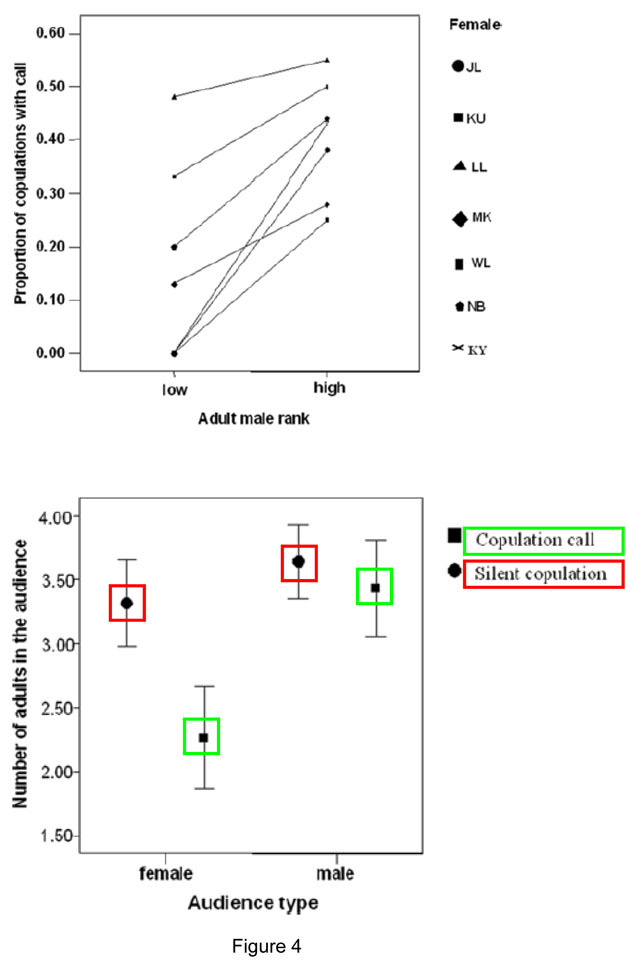
Figure 4: Female chimpanzees produce copulation calls during sexual interactions with males.
Call rates are affected by the social rank of the male partner and the composition of the nearby audience. Both mating with low-ranking males and the presence of large female audiences tend to inhibit copulation calls in different females.
© 2012 Nature Education Data from Townsend et al. (2008). All rights reserved.
Humans, however, go a step further, with signallers and receivers communicating in relation to their shared intentions (Tomasello 2008). As a consequence, human communication is bi-directional, conversational, and based on conventions as to how intentions are to be communicated. A particularly interesting example is pointing, which emerges early and universally during human development, with no direct equivalent in primate communication. Although the pointing gesture has no independent meaning, it is very effective in triggering shared attention between signallers and receivers in relation to an external referent. Apes also produce gestures to request objects, but there is no good evidence that they point for others solely to be informative. The degree to which primates perceive and share each other's intentions, and to communicate as part of this cooperative experience, is the object of much ongoing research. Results are likely to delineate more clearly the relationship between primate communication and human language. As such they will provide progress towards one of the most interesting questions in science. What part of our biology is uniquely human and what is part of our primate heritage?
References and Recommended Reading
Arnold, K. & Zuberbühler, K. Language evolution: Semantic combinations in primate calls. Nature 441, 303 (2006).
———— Meaningful call combinations in a non-human primate. Current Biology 18, R202-R203 (2008).
Call, J. & Tomasello, M. eds. The Gestural Communication of Apes and Monkeys. Mahwah, NJ and London, UK: Lawrence Erlbaum Associates, 2007.
Cartmill, E. A. & Byrne, R. W. Semantics of primate gestures: Intentional meanings of orangutan gestures. Animal Cognition 13, 793-804 (2010).
Cheney, D. L. & Seyfarth, R. M. Assessment of meaning and the detection of unreliable signals by vervet monkeys. Animal Behaviour 36, 477-486 (1988).
Chomsky, N. Three factors in language design. Linguistic Inquiry 36, 1-22 (2005).
Clay, Z. & Zuberbühler, K. Bonobos extract meaning from call sequences. PLoS ONE 6, e18786 (2010).
Crockford, C. et al.. Wild chimpanzees inform ignorant group members of danger. Current Biology 22, 142-146 (2012).
Enard, W. et al. Molecular evolution of FOXP2, a gene involved in speech and language. Nature 418, 869-872 (2002).
Evans, N. & Levinson, S. C. The myth of language universals: Language diversity and its importance for cognitive science. Behavioral and Brain Sciences 32, 429-448 (2009).
Genty, E. et al. Gestural communication of the gorilla (Gorilla gorilla): Repertoire, intentionality and possible origins. Animal Cognition 12, 527-546 (2009).
Gliga, T. & Csibra, G. One-year-old infants appreciate the referential nature of deictic gestures and words. Psychological Science 20, 347-353 (2009).
Grice, H. P. Utterers' meaning and intentions. Philosophical Review 78, 147-177 (1969).
Guilford, T. & Dawkins, M. S. Receiver psychology and the evolution of animal signals. Animal Behaviour 42, 1-14 (1991).
Hobaiter, C. & Byrne, R. W. The gestural repertoire of the wild chimpanzee. Animal Cognition 14, 745-767 (2011).
Janik, V. M. Acoustic communication in delphinids. Advances in the Study of Behavior 40, 123-157 (2009).
Jolly, A. Lemur social behavior and primate intelligence. Science 153, 501-506 (1966).
Laporte, M. N. C. & Zuberbühler, K. Vocal greeting behaviour in wild chimpanzee females. Animal Behaviour 80, 467-473 (2010).
McComb, K. & Semple, S. Coevolution of vocal communication and sociality in primates. Biology Letters 1, 381-385 (2005).
Ouattara, K., Lemasson, A. & Zuberbühler, K. Campbell's monkeys concatenate vocalizations into context-specific call sequences. Proceedings of the National Academy of Sciences of the United States of America 106, 22026-22031 (2009).
Savage-Rumbaugh, S. et al. Spontaneous symbol acquisition and communicative use by pygmy chimpanzees (Pan paniscus). Journal of Experimental Psychology: General 115, 211-235 (1986).
Seyfarth, R. M., Cheney, D. L. & Marler, P. Monkey responses to three different alarm calls: Evidence of predator classification and semantic communication. Science 210, 801-803 (1980).
Silk, J. B., Kaldor, E. & Boyd, R. Cheap talk when interests conflict. Animal Behaviour 59, 423-432 (2000).
Stern, K. & McClintock, M. K. Regulation of ovulation by human pheromones. Nature 392, 177-179 (1998).
Tomasello, M. Origins of Human Communication. Cambridge, MA: MIT Press, 2008.
Townsend, S. W., Deschner, T. & Zuberbühler, K. Female chimpanzees use copulation calls flexibly to prevent social competition. PLoS ONE 3, e2431 (2008).
van Hooff, J. "A structural analysis of the social behavior of a semi-captive group of chimpanzees" in Social Communication and Movement: Studies of Interaction and Expression in Man and Chimpanzee, eds. M. von Cranach & I. Vine (London, UK: Academic Press, 1973) 75-162.
von Humboldt, K. W. Über die Verschiedenheit des menschlichen Sprachbaues und ihren Einfluss auf die geistige Entwickelung des Menschengeschlechts. Berlin, Germany: Druckerei der Königlichen Akademie der Wissenschaften, 1836/1935.
Wedekind, C. et al. MHC-dependent mate preferences in humans. Proceedings of the Royal Society B: Biological Sciences 260, 245-249 (1995).
Wheeler, B. C. Monkeys crying wolf? Tufted capuchin monkeys use anti-predator calls to usurp resources from conspecifics. Proceedings of the Royal Society B: Biological Sciences 276, 3013-3018 (2009).
Wich, S. A. & de Vries, H. Male monkeys remember which group members have given alarm calls. Proceedings of the Royal Society B: Biological Sciences 273, 735-740 (2006).
Zahavi, A. Mate selection: A selection for a handicap. Journal of Theoretical Biology 53, 205-214 (1975).
Zuberbühler, K., Cheney, D. L. & Seyfarth, R. M. Conceptual semantics in a nonhuman primate. Journal of Comparative Psychology 113, 33-42 (1999).










