





















« Prev Next »

What is Macroevolution?
The tree of life has many branches that all connect to a common ancestor, and the diversity of life on the tree results from evolutionary processes. Just as we organize life on earth into hierarchies, we would like to do the same for evolutionary processes and patterns. Thus, many scientists propose that evolution can be divided into two distinct hierarchical processes -- microevolution and macroevolution -- although the distinction between them is somewhat artificial. Microevolution describes mechanisms that alter the frequencies of alleles in gene pools within species (Rexnick & Ricklefs 2009). These mechanisms include mutation, migration, genetic drift, and natural selection. Theory suggests that the effects of these processes accumulate over time and can sometimes result in the divergence of populations and the birth of new species. In contrast, macroevolution describes patterns on the tree of life at a grand scale across vast time periods. Many different patterns can be observed across the tree of life at a grand scale (Figure 1), including stability, gradual change, rapid change, adaptive radiations, extinctions, the co-evolution of two or more species, and convergent evolution in traits between species -- just to name a few. Macroevolutionary studies tend to draw heavily from the fossil record. Fossils document the emergence of new life forms, how their geographic distribution changed over time, and ultimately when they went extinct. In contrast, microevolutionary changes are not frequently observed in the fossil record because the processes that govern evolutionary change within species are thought to occur over much shorter time scales. Thus, macroevolution is centered on explaining evolutionary patterns above the species level (Rexnick & Ricklefs 2009), and those who study it are searching for the ‘organizing principles' that explain these patterns.
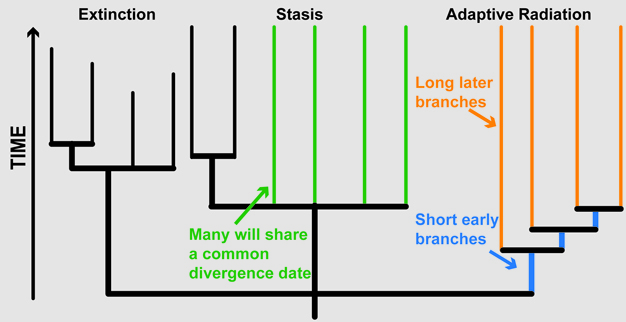
Figure 1
Example of macroevolutionary patterns as they would appear in a phylogenetic tree, including extinctions, adaptive radiations, and stasis.
© 2012 Nature Education All rights reserved. 
Why is it important?
Understanding macroevolution is important because it explains both the diversity of life and the pace of evolutionary change. Does evolution happen slowly or quickly? There are two main schools of thought about the tempo and mode of macroevolution (Eldredge & Gould 1972, Eldredge et al. 2005). One group holds that microevolutionary processes alone can sufficiently explain grand patterns and radical changes on the tree of life. In other words, mutation, migration, genetic drift, and natural selection can produce major evolutionary changes given enough time. The key element is vast amounts of time -- on a scale that is difficult for most people to imagine. This model of macroevolution is called phyletic gradualism. It proposes that most speciation events are the result of a gradual and uniform transformation of one species into a new one through a process called anagenesis.
On the other hand, many scientists propose that grand patterns in the history of life cannot be explained exclusively by changes in allele frequencies over time, even rapid ones. Instead, these scientists propose that large changes on the tree of life were preceded by events that decoupled the tempo and mode of evolutionary change from predictable microevolutionary processes. Often these decoupling events were major, even cataclysmic, environmental changes that opened up new niches or caused extinctions. A classic example of such an event was the meteor impact at the end of the Cretaceous that contributed to the extinction of the dinosaurs, and as a consequence, opened up new niches that precipitated the diversification of mammals. There are many other examples of such events, including the acquisition of novel traits like the advent of flowers as a means of reproduction in plants. This model of macroevolution is called punctuated equilibrium (Eldredge & Gould 1972). It proposes that grand patterns of evolutionary change on the tree of life involve the rapid splitting of one ancestral species into two or more descendant species through cladogenesis, often followed by long periods of stasis in the descendant species (Eldredge et al. 2005).
Are there examples of macroevolution on our twig of the tree of life?
Patterns of macroevolution are easy to spot on the tree of life when one considers big events like the abrupt appearance of tetrapods in the fossil record, long periods of stasis like that observed in sharks and crocodiles, and adaptive radiations including the (fairly!) recent diversification of mammals that began about 70 million years ago (mya). As one moves out along the branches of the tree of life, the processes that produced the rich patterns of biodiversity along a particular twig can be harder to understand and interpret.
Yet, there are many examples of macroevolutionary phenomena found in the order Primates, including stasis, adaptive radiations, extinctions of entire lineages, co-evolution, and convergent evolution.
Adaptive radiations and stasis -- Phylogenetic trees across the order Primates
Recent studies have provided new insights about the tempo and mode of primate evolution using phylogenetic trees from genetic data gathered across the genomes of many extant primate lineages (Fabre et al. 2009, Perelman et al. 2011). These studies have revealed that the tempo and mode of evolution among the primates have been punctuated by the persistence of ancient relic lineages (i.e., stasis), bursts of speciation that may be consistent with adaptive radiations, and even by ongoing speciation that is governed by microevolutionary processes. Perelman et al. (2011) recently constructed a primate phylogenetic tree for 61 primate genera (Figure 2). The long branch that separates Tarsiers from other primates suggests that this group is an ancient relict lineage that has remained in stasis relative to other primates. In contrast, the Lemuriformes part of the tree has many early short branches followed by some long branches in the descendants (see Figure 1), which suggests that the ancestors of extant lemurs experienced a rapid adaptive radiation that likely coincided with its colonization of Madagascar about 62-65 mya (Perelman et al. 2011).
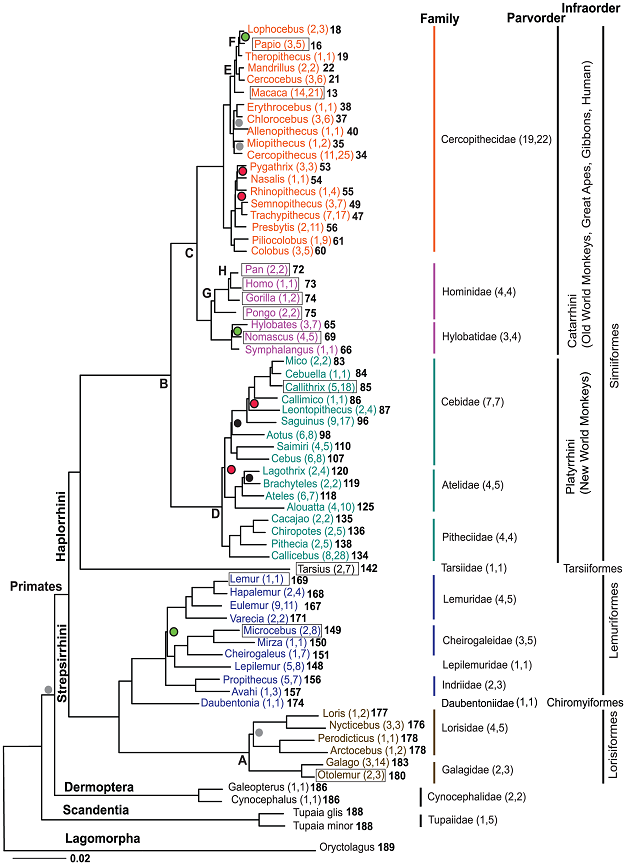
Figure 1
A molecular phylogeny of 61 primate genera.
Perelman, P. et al. A molecular phylogeny of living primates. PLoS Genetics 7 (2011).
Phylogenetic trees also allow for comparing and contrasting the tempo and mode of evolution among different groups of primates inferred from fossil and genetic data. For instance, has evolution proceeded differently in New World monkeys versus Old World monkeys? New World monkeys last shared a common ancestor with Old World monkeys about 30-50 mya, but the diversification of New World monkeys and the divergence times of these lineages are not well understood. The fossil record suggests that New World monkeys have been in stasis following their initial colonization of the Americas, while Old World monkeys show evidence of ‘faunal turnover' that closely matches the patterns predicted under the punctuated equilibrium model (Delson& Rosenberger 1984, Rosenberger 2002). Hodgson et al. (2009) used molecular data to construct phylogenetic trees and to estimate divergence dates for many New World monkey species to examine the hypothesis that they have been in stasis relative to other primates. They found that New World monkeys have experienced both successive radiations and stasis during their evolution. Specifically, they found that the earliest New World monkey fossils were much older than the divergence dates they estimated for the extant New World monkey species. Using this evidence, along with patterns observed on phylogenetic trees, these researchers suggested that there was an early radiation of New World monkey ancestors followed by a period of stasis and then the extinction of most of this group prior to the Miocene. Following this period, the survivors of the original radiation then experienced a burst of rapid diversification into what would become the extant New World monkey ‘crown lineages' (Hodgson et al. 2009).
Adaptive radiations and extinctions -- The rise and fall of Miocene Apes
Phylogenetic trees based on genetic data cannot reveal much about what might have caused adaptive radiations or extinctions. Careful examination of fossils combined with an understanding about what Earth's environment was like when these fossils were living can be used to infer what might have precipitated different macroevolutionary events. For example, during the Miocene the ancestors of Old World monkeys and apes experienced both radiations and extinctions that have been linked to climate change (Harrison 2010). In the early Miocene, primates found in Africa and the Arabian Peninsula were a diverse group that occupied tropical forests and woodlands (Figure 3). During the mid-Miocene, Africa reconnected with Eurasia and a major period of global warming caused the expansion of tropical habitats northward. These developments allowed the nascent hominoid lineage to branch off and colonize newly available Eurasian habitats, leading to a major proliferation of ape species across much of Eurasia. However, around 9.6 mya, a major shift to drier climates created more open habitats that led to a decline of hominoid taxa in Eurasia. By 5 mya, most ape species were extinct, except for a few that eventually led to modern-day orangutans and gibbons ( Moyà-Solà et al. 2009, Harrison 2010).
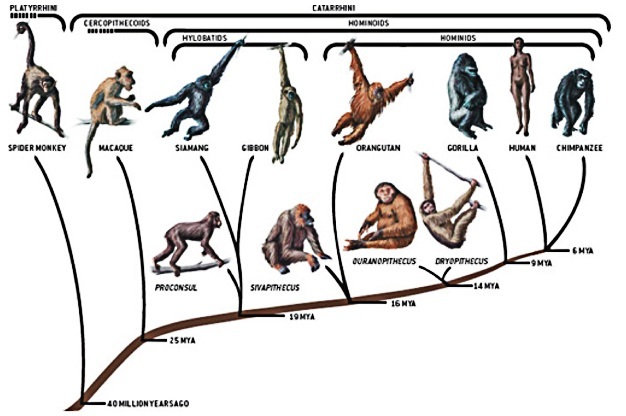
Figure 3
The family tree of extant hominoids includes only a small fraction of the diversity of apes that have lived on this planet. During the Miocene, up to 100 ape species once lived throughout much of Europe and Asia, but ultimately went extinct. Proconsul may have been the last common ancestor of extant hominoids. Sivapithecus was probably an ancestor to orangutans. Ouranopithecus or Dryopithecus appeared in the fossil record later in the Miocene than Proconsul and Sivapithecus. Both have been proposed as ancestors shared by all living hominoids.
Source: Planet of the Apes. David Begun and John Gurche, Scientific American, 16, 4-13 (2006) doi: 10.1038/scientificamerican0606-4sp
Anagenesis -- A gradual transition from Homo heidelbergensis to H. neanderthalensis
Understanding the tempo and mode of primate evolution is challenging because primate fossils are quite rare. Pleistocene hominins in Europe are an exception, however, since a more complete fossil record and complementary genetic data are available for this group. Comparison of Neanderthal and modern human DNA suggests that these two species shared a last common ancestor, most likely Homo heidelbergensis, sometime between 0.35 and 0.40 mya (Green et al. 2006, Noonan et al. 2006). These big-brained, Middle Pleistocene hominins are found in many places across the Old World and occur in the fossil record from about 1.3 mya to about 0.20 mya. They are sometimes called the ‘muddle in the middle'. This group got its nickname because there are no morphological features that definitively distinguish H. heidelbergensis from its predecessor (H. erectus) and its likely descendants in Africa (H. sapiens) and in Europe (H. neanderthalensis) (Figure 4). This lack of diagnostic characteristics is important for understanding the tempo and mode of the later stages of human evolution. Some paleoanthropologists recognize nascent Neanderthal-like characteristics in the European branch of H. heidelbergensis by a gradual change over time in the fossil record towards distinctive Neanderthal traits, including large brow ridges, a slightly protruding face, and the absence of a prominent chin (Dean et al. 1998, Harvarti 2007, Hublin 2009). These findings have led some researchers to propose that this is evidence for a continuous evolution (i.e., anagenesis) from H. heidelbergenensis to H. neanderthalensis (Dean et al. 1998, Hublin 2009).
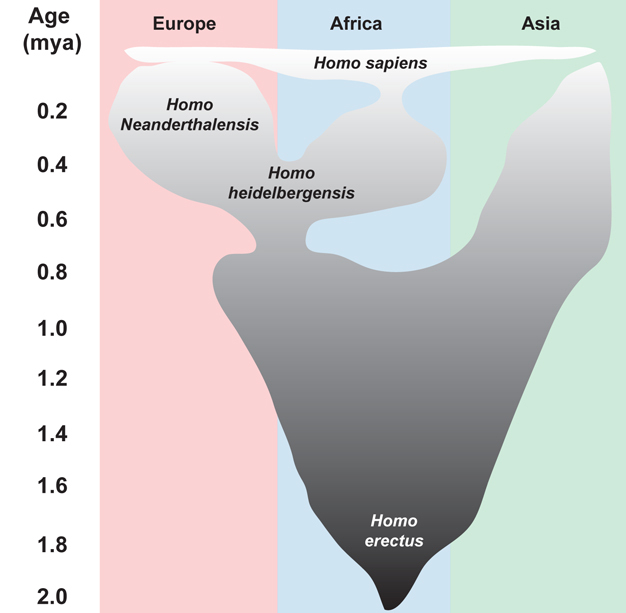
Figure 4
One model of Middle Pleistocene human evolution. There is evidence to suggest that a gradual transition from Homo heidelbergensis to H. neanderthalensis occurred in Europe.
Adapted from Harvarti, K. 100 years of Homo heidelbergensis – life and times of a controversial taxon. Mitteilungen der Gesellschaft für Urgeschichte 16, 85-94 (2007) and Hublin, J. J. The origin of Neandertals. Proceedings of the National Academy of Sciences (USA) 106, 16022-16027 (2009).
Convergent evolution -- Color vision in howler monkeys
A macroevolutionary perspective can also reveal patterns of convergent evolution, such as the evolution of color vision in primates (Figure 5). Most mammals are dichromats. This means that their vision is based on two kinds of visual pigments, or opsins. Many primates, including humans, are unique among mammals because they are trichromats who possess three types of pigments that allow them to perceive a richer array of colors compared to dichromats (Jacobs 2008). There are two ways for a primate to be a trichromat. The S opsin is encoded by a gene located on chromosome 7, and is shared by all primates. Old World monkeys, hominoids, and humans have two additional opsin genes, located on the X chromosome, that encode pigments called ‘L' and ‘M'. New World monkeys, however, have only a single, polymorphic M/L opsin gene. Since the X chromosome is inherited from the mother and males are the heterogametic sex, every New World monkey male is a dichromat because he can only have a single M or L pigment along with his S photopigment. Females have two copies of the X chromosome, and thus, New World monkey females can be either dichromats or trichromats (Jacobs 2008). Dirunal howler monkeys (Alouatta) are exceptions among New World primates. Both males and females can be trichromats due to a recent gene duplication of the M/L opsin gene (Jacobs et al. 1996, Boissinot et al. 1998). This ‘reinvention' of trichromacy suggests that it provided them with an evolutionary advantage (Lucas et al. 2003). This example illustrates the importance of having a macroevolutionary perspective on primate evolution because convergent evolution in color vision would not be obvious without a broad perspective on primate evolution.
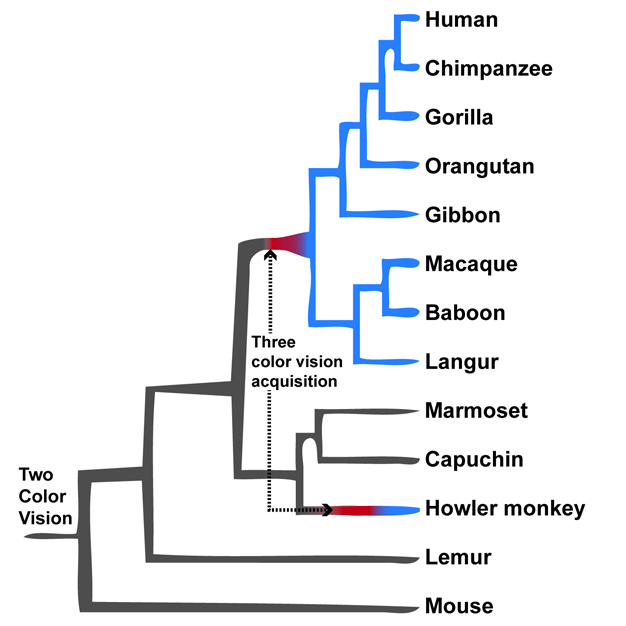
Figure 5
Types of color vision among major primate groups. Apes and Old World monkeys have three types of visual pigments, whereas nearly all male New World monkeys and prosimians have two types of visual pigments. Howler monkeys are unusual New World monkeys who have ‘re-evolved’ a third visual pigment through a gene duplication event.
© 2012 Nature Education All rights reserved.
Summary
Macroevolution involves studying patterns on the tree of life above the species level, and inferring the processes that are likely to have generated these patterns. Thus, macroevolutionary thinking helps to build a more complete picture about the evolutionary history of primates.
Glossary
adaptive radiation: A rapid increase in the number of species in a lineage with new members showing differences in traits that are likely adaptations to different niches
anagenesis: The process by which a species gradually transitions into a new species. Anagenesis involves change across an entire population rather than a branching event, as in cladogenesis
cladogenesis: The process by which a species splits rapidly into two distinct species, rather than one species gradually transforming into another
co-evolution: Adaptive changes in an organism triggered by an adaptive change in another organism
convergent evolution: The acquisition of the same biological trait in unrelated or distantly related organisms
opsin: A protein found in the photoreceptor cells of the retina
phylogenetic tree : A branching diagram showing the inferred evolutionary relationships among different species or other biological classifications. Connections between nodes implies common ancestry of two or more branches, and sometimes branch length can be proportional to time since separation from a common ancestorReferences and Recommended Reading
Boissinot, S. et al. Origins and antiquity of x-linked triallelic color vision systems in new world monkeys. Proceedings of the National Academy of Sciences (USA) 95, 13749-13754 (1998).
Dean, D. et al. On the phylogenetic position of the pre-Neandertal specimen from Reilingen, Germany. Journal of Human Evolution 34, 485-508 (1998).
Delson, E. & Rosenberger, A. L. "Are there any anthropoid primate living fossils?" in Living Fossils, eds. N. Eldridge and S. M. Stanley (New York, Springer Verlag: 1984), 50-61.
Eldredge, N. & Gould, S. J. "Punctuated equilibrium: An alternative to phyletic gradualism," in Models in Paleobiology, ed. T. J. M. Schopf (San Francisco, Freeman Cooper: 1972), 82-115.
Eldredge, N. et al. The dynamics of evolutionary stasis. Paleobiology 31, 133-145 (2005).
Fabre, P. H. et al. Patterns of macroevolution among primates inferred from a supermatrix of mitochondrial and nuclear DNA. Molecular Phylogenetics and Evolution 53, 808-825 (2009).
Green, R. E. et al. Analysis of one million base pairs of Neanderthal DNA. Nature 444, 330-336 (2006).
Harrison, T. Apes among the tangled branches of human origins. Science 327, 532-534 (2010).
Harvarti, K. 100 years of Homo heidelbergensis - life and times of a controversial taxon. Mitteilungen der Gesellschaft für Urgeschichte 16, 85-94 (2007).
Hodgson, J. A. et al. Successive radiations, not stasis, in the South American primate fauna. Proceedings of the National Academy of Sciences (USA) 106, 5534-5539 (2009).
Hublin, J. J. The origin of Neandertals. Proceedings of the National Academy of Sciences (USA) 106, 16022-16027 (2009).
Jacobs, G. H. Primate color vision: A comparative perspective. Visual Neuroscience 25, 619-633 (2008).
Jacobs, G. H. et al. Trichromatic colour vision in New World monkeys. Nature 382, 156-158 (1996).
Lucas, P. W. et al. Evolution and function of routine trichromatic vision in primates. Evolution 57, 2636-2643 (2003).
Moyà-Solà, S. et al. A unique Middle Miocene European hominoid and the origins of the great ape and human clade. Proceedings of the National Academy of Sciences (USA) 106, 9601-9606 (2009).
Noonan, J. P. et al. Sequencing and analysis of Neanderthal genomic DNA. Science 314, 1113-1118 (2006).
Perelman, P. et al. A molecular phylogeny of living primates. PLoS Genetics 7 (2011).
Reznick, D. N. & Ricklefs, R. E. Darwin's bridge between microevolution and macroevolution. Nature 457, 837-842 (2009).
Rosenberger, A. L. "Platyrrhine paleontology and systematics: The paradigm shifts," in The Primate Fossil Record, ed. W. C. Hartwig (Cambridge, UK, Cambridge University Press: 2002) 151-159.










