





















« Prev Next »
It was 2004 when a team of joint Australian-Indonesian researchers announced the discovery of fossils and simple (Oldowan-like) stone tools from Liang Bua cave (which translates to "cool cave") on the remote Indonesian island of Flores. The fossils represent a small-bodied and small-brained hominin, named Homo floresiensis, but better known as the "Hobbit." The position of these fossils on the human evolutionary tree remains unclear. In fact, since the 2004 discovery, there has been an unending series of controversies surrounding these specimens.
What is perhaps most surprising is the young geological age of these fossils. The fossils range in age from 74 to 17 ka (Brown et al., 2004), which falls well within the range of modern humans elsewhere in the Old and quite possibly New World. However, only the younger layers above the Homo floresiensis fossils show evidence of modern human occupation at Liang Bua.
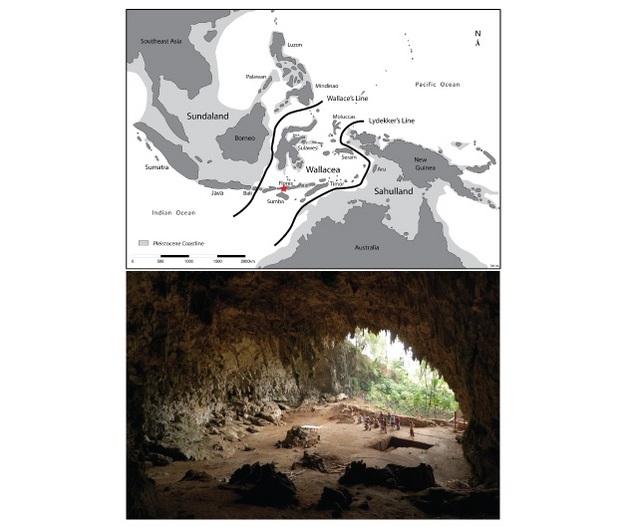
Figure 1: A map of island southeast Asia in the region of Flores and a photograph of the Liang Bua cave where the Homo floresiensis fossils were recovered.
The region between the two lines on the map is called Wallacea. Very few large terrestrial animal migrated into this region from either the east or west.
© 2012 Nature Education Map courtesy of Mike Morewood; photo via Wikipedia. Some rights reserved. 
Fossil and Archaeological Evidence
The LB1 skeleton (type specimen of the new species) preserves a nearly complete skull, a partial pelvis, several limb bones, and bones of the hands and feet. This individual is estimated to have stood about 1.06 m (3'6") tall, which is roughly the size of a 3–4 year old modern human child. To date, LB1 has the only complete skull to be recovered, but a second mandible (lower jaw) and numerous skeletal elements from a second individual (LB6) have been described (Brown et al., 2004). Fossils of at least four other individuals have also been recovered, confirming that this was a population of small-bodied individuals and that LB1 was not an anomaly.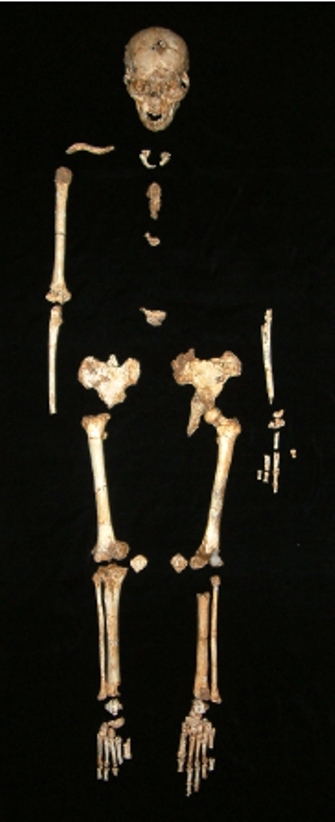
Figure 2: Photograph of the LB1 skeleton, the type specimen of Homo floresiensis, laid out in approximate anatomical position.
© 2012 Nature Education Photo courtesy of William Jungers. All rights reserved.
Initial descriptions emphasized the mix of ancestral traits that remained unchanged from more ancient species and derived traits that linked it with more recent ones. The skull resembles those belonging to extinct species of our own genus Homo (Brown et al., 2004; Baab and McNulty, 2009). The skeleton, however, is considerably more primitive and in some respects aligns the LB1 specimen and the other Flores fossils with older and even more primitive species like those belonging to Australopithecus afarensis (which includes "Lucy"; Tocheri et al., 2007; Jungers et al., 2009) (link to Ward and Hammond article; link to Schrein article). Taken together, this is a puzzling pattern: a population that existed between 74 and 18 ka, with a skull that most closely resembles the much older Homo habilis or Homo erectus, and a skeleton that retains some features normally associated with australopith species at least 3 Ma.
Presumably Homo floresiensis made the simple Oldowan-like tools (the oldest and most primitive type known in the archaeological record) found in the same layers as the skeletal remains at Liang Bua cave (Moore and Brumm, 2009). These stone tools resemble those found elsewhere on the island at sites that are closer to a million years in age (Brumm et al., 2006; 2010). In addition to tools, there is also evidence in the form of cut marks on some Stegodon bones indicating that the hominins were butchering these animals (Morwood et al., 2005). There are also some burnt bones and pebbles, but whether this was the result of intentional or accidental fire is still unknown (Westaway et al., 2009).
Evolutionary Scenarios
As a result of the primitive morphology but surprisingly young geological age of the fossils, many scenarios have been proposed to explain the presence of these fossils on Flores. Homo erectus occupied Southeast Asia from about 1.5 million years to perhaps as recently as half a million years ago or even 50 thousand years ago (Larick et al., 2001; Huffman et al., 2010; Indriati et al., 2011). The low neurocranium (the part surrounding the brain) with a flat and sloping forehead, thick cranial bones, short and flat face, and other details of LB1's skull anatomy (e.g., an occipital torus and a mastoid fissure), as well as the shape of the brain provide a link to Homo erectus, but both the small body size and brain size are outside of the expected range for that species (Brown et al., 2004; Falk et al., 2005; Baab and McNulty, 2009). Therefore, based primarily on the cranial evidence, the original description proposed that these fossils represented a new species, Homo floresiensis, that was a dwarfed descendent of Homo erectus. Additional aspects of anatomy also resemble that known for Homo erectus, including the shape of the brain and the shoulder (Falk et al., 2005; Larson et al., 2009). According to this theory, the ancestors of Homo floresiensis somehow made the treacherous water crossing to reach Flores and over time there was a reduction in body size (Meijer et al., 2010).
The phenomenon of island dwarfing has been documented for a number of other large-bodied mammals, including primates, mammoths, and deer. There are also numerous instances of human populations becoming dwarfed, including on the islands of Southeast Asia. The impetus for island dwarfing is often linked to reduced availability of resources in the environment. In either case, being small-bodied may be more advantageous on an island than on the mainland. Flores exists in a region known as Wallacea where there is a low level of migration from the Asian and Australasian faunas (collections of animals) to the east and west, respectively. These islands appear to have been isolated by strong currents, even when sea levels were low. Along with the hominins, a few other large-bodied mammals have been documented in this region, including Stegodon, a type of extinct type of elephant, and komodo dragons. Both of these species were found in deposits coeval with Homo floresiensis, however, their rarity is further confirmation that it was difficult for large terrestrial animals to reach islands in Wallacea. Interestingly, the particular subspecies of Stegodon found in the same layers of Liang Bua cave as Homo floresiensis, Stegodon florensis insularis, is itself a dwarfed form whose closest relative is the distinctly larger and geologically older Stegodon florensis florensis (van den Bergh et al., 2008).
An argument against island dwarfing as an explanation for Homo floresiensis stems from the relationship between brain size and body size. LB1's endocranial volume, calculated from CT scans of the skull, is only 417 cm3, close to a third of the average modern human value (Falk et al., 2005). This small brain size is difficult to explain based on typical patterns of island dwarfing assuming that the ancestor resembled a modern human or even Homo erectus (Martin et al., 2006). This is because brain size typically "dwarfs" less than overall body size. For example, despite having bodies that are much smaller than their neighbors, modern human pygmies have brains which are only slightly smaller. However, more drastic decreases in brain size compared to body size during island dwarfing are not entirely without precedent and may have occurred in at least two other groups: Myotragus ("cave goat") on Mallorca and Hippopotamus lemerlei on Madagascar (Köhler and Moyà-Solà, 2004; Weston and Lister, 2009).
A second model advanced to explain the presence of the hominin fossils on the island of Flores in the Pleistocene states that this population was the offshoot of a more primitive, pre-erectus hominin species with a small body size and small brain. Evidence from the mandible and the rest of the skeleton supports this hypothesis (Argue et al., 2009). The size and morphology of the teeth and mandible share more resemblances to Australopithecus and the earliest Homo species than to Homo erectus (Brown and Maeda, 2009). In particular, the very short legs (relative both to the arms and to the feet) are a pattern seen in apes and australopiths rather than Homo erectus (a good Homo habilis skeletal comparison has not yet been discovered). LB1 was also disproportionately heavy for her height — a pattern closely approximated by the famous 3.2 million year old Australopithecus afarensis skeleton of "Lucy" (Jungers and Baab, 2009). Despite being only 106 cm in height, LB1 is estimated to have weighed close to 32.5 kg (71.7 lbs.). The carpal bones (bones of the wrist) in Homo floresiensis look more like those of chimpanzees than of modern humans (Tocheri et al., 2007). While the evolution of the wrist is not well documented in early members of the genus Homo (e.g., Homo habilis and Homo erectus), it is clear that the wrist morphology is more primitive than that of modern humans and Neanderthals. Like the wrist, the foot morphology, although exhibiting some human-like traits (including a non-grasping big toe), also retains several quite primitive features: a long forefoot with curved toes and the lack of a medial longitudinal arch (Jungers et al., 2009). The latter trait is indicated by the particular shape of the navicular bone, one of the bones close to the heel. This medial longitudinal arch is located on the inside aspect of the foot, forming the instep, and acts both as a shock absorber and a spring during walking. It seems likely that the relatively long foot and particular combination of traits present in the foot would have resulted in a slightly different walking gait than seen in modern humans. This combination of traits in the skeleton appears to place the ancestry of the Homo floresiensis skeleton earlier than Homo erectus.
One major challenge to the idea that Homo floresiensis has an ancestry deeper than Homo erectus is the absence of fossils of any such species in either island or mainland Southeast Asia. The hominin fossil record prior to Homo erectus is found only in Africa. A second complication is that the fossil record of postcranial anatomy for pre-erectus species of Homo is poor and their morphology is not as well documented as other species, so comparison with Homo floresiensis is limited. As a result, it is not clear whether a species such as Homo habilis, which precedes Homo erectus geochronologically, is in fact a good model for the ancestor of Homo floresiensis.
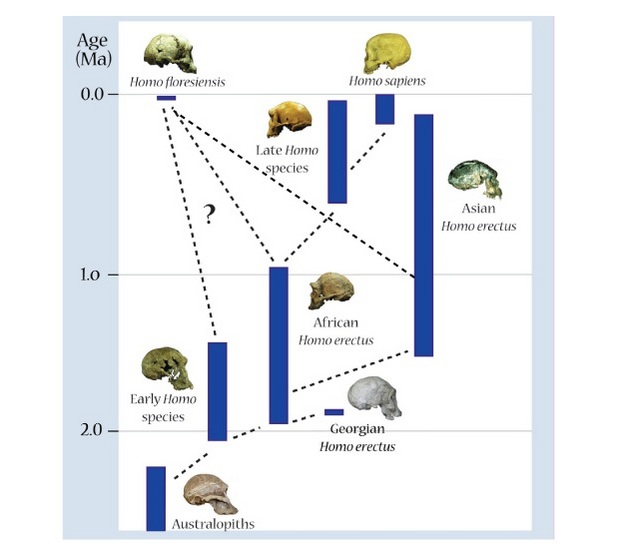
Figure 3: Phylogenetic tree demonstrating possible evolutionary relationships among Homo taxa, illustrating some of the possible ancestors of Homo floresiensis: Asian Homo erectus or earlier Homo populations from Africa.
© 2012 Nature Education Photo courtesy of Mike Morwood, Karen Baab, Peter Brown, and the Kenya National Museums. All rights reserved.
Pathological Hypotheses
In contrast to the two evolutionary hypotheses, a third theory has been advanced: the Flores remains represent modern humans suffering from some type of pathological condition (a disease). The initial suggestion was that LB1 was a modern human that suffered from microcephaly, a condition in which the neurocranium is considerably smaller than that of normal, healthy people (Henneberg and Thorne, 2004; Martin et al., 2006; Jacob et al., 2006). This can be the result of a genetic disorder whose main symptoms are under-development of the brain and the overlying cranial vault — a condition known as primary microcephaly. However, microcephaly is a symptom of a range of other disorders, some of which include short stature. The proponents of the pathology hypothesis have thus far failed to identify exactly what disorder can account for the large number of apparently primitive traits in the LB1 skeleton. There is a clear similarity in the shape of the skull between archaic species of Homo and humans with microcephaly — primarily in the high ratio of facial to neurocranial size. This convergence in shape occurs for two very different reasons: microcephalic humans have pathologically underdeveloped brains whereas early hominins belonged to species normally characterized by smaller brains than those of modern humans. These similarities are only superficial, however, and more detailed examination of the cranial vault shape, as well as the shape of the underlying brain, shows that LB1 shares important characteristics with fossil Homo species (Falk et al., 2005, 2007; Baab, 2010).
Figure 4: The LB1 cranium (center) compared to skulls of a modern human with endemic hypothyroidism (left) and a modern human showing microcephaly (right).
All photographs were scaled to the same maximum cranial length to emphasize shape differences among them. For example, the skull of the "cretin" (on the left) is much taller but shorter front-to-back than LB1. Despite the superficial similarities due to a small neurocranium relative to face in both LB1 and the microcephalic modern human, the midline profile of the neurocranium and the facial skeleton differ among them. The photograph of the human with endemic hypothyroidism was mirrored for easier comparison with the other photographs.
© 2012 Nature Education Photo courtesy of Karen Baab and Peter Brown. All rights reserved.
In addition to microcephaly, two other very different pathologies have been implicated: myxoedematous endemic hypothyroidism ("cretinism"; Obendorf et al., 2007; Oxnard et al., 2010) and Laron Syndrome (Hershkovitz et al., 2007). The former is caused by a lack of iodine in the diet during both pre- and post-natal development (Halpern et al., 1991); iodine is crucial for the proper production of hormones in the thyroid gland and for the development of the central nervous system. The most notable symptoms of this disease include mental retardation and other neurological defects as well as short stature due to delayed skeletal maturity (Boyages et al, 1988). The skull is often described as brachycephalic, and the sutures between individual bones of the skull frequently remain open even once brain growth has ended (Koplik, 1918, p. 44). However, the LB1 skull is not brachycephalic, and many of its cranial sutures are fully fused. In fact, the skulls of cretins are not particularly small, and their shape more closely resembles healthy humans than LB1 (Baab, 2010). Some skulls of patients with congenital hypothyroidism exhibit an enlarged pituitary fossa. This feature was initially attributed to the skull of LB1 (Obendorf et al., 2007), but this has been refuted by measurements from CT scans of the skull, as have a number of other alleged similarities between LB1 and humans with myxoedematous hypothyroidism (Jungers et al., 2009). While the skeletons of both human cretins and Homo floresiensis differ from that of healthy modern humans, they do not resemble one another. For example, the limb proportions of LB1 are not comparable to those of humans with cretinism (Jungers et al., 2009; Groves and Fitzgerald, 2010).
Laron Syndrome is characterized by insensitivity to growth hormones; although the body manufactures the correct growth hormones, the receptors in the body do not respond properly to them. As in myexedemotous hypothyroidism, patients suffering from Laron Syndrome are of shorter than average stature. Although the original paper proposing that LB1 had Laron Syndrome found a number of similarities between the LB1 skeleton and individuals with Laron Syndrome (Hershkovitz et al., 2007), it is now clear that the case was overstated (Falk et al., 2009), and that the evidence for this particular syndrome in LB1 is nonexistent. For example, with regards to the skull, patients with Laron Syndrome typically have a protruding forehead, underdeveloped facial bones such that the face looks small compared to the rest of the head, and a skull that is disproportionately wide across the parietal bones compared to the base of the skull. The LB1 skull shows the opposite pattern: the forehead slants backwards rather than protruding, the face is large relative to the rest of the skull, and it is wide at the base rather than the parietals (Falk et al., 2009; Baab, 2010).
Conclusions
The rather unusual combination of short stature, a small brain, primitive skeletal anatomy, simple stone tools, all occurring at a time when fully modern humans inhabited other parts of the globe has led to a wide range of explanations (summarized above and in more detail in Aiello [2010]). Consensus among researchers regarding the interpretation of the Liang Bua fossils remains elusive as the study of the fossils is ongoing and apparently contradictory interpretations have been published. How can we make sense of this? Both cretinism and Laron Syndrome result in humans of small body size, which is consistent with LB1. However, the other symptoms of these diseases are not observed in the LB1 skeleton. Furthermore, LB1 was not an isolated find, and the other individuals found in Liang Bua cave are of a similarly diminutive size. As it is highly improbable that all members of this population were diseased, short stature should be viewed as a characteristic of the population rather than a pathological symptom of a single individual. Microcephaly is a more difficult argument to dismiss because it is not a disease per se but rather a symptom that accompanies many disorders. However, the shape of the LB1 neurocranium does not resemble skulls of human microcephalics thus far examined (Baab, 2010) and a single disorder that explains the apparently primitive features found elsewhere in the skeleton has yet to be described. Taken together, the weight of evidence does not support a pathological explanation for the particular characteristics found in LB1 and her kin in Liang Bua cave.Rather, the various lines of evidence, from the simple stone tools, the Homo-like skull morphology, and the even more primitive skeletal anatomy, paint a more complicated evolutionary picture. The two most popular evolutionary hypotheses position Homo floresiensis as the dwarfed descendant of Homo erectus or descendant of an even more primitive species. If the former is correct, then Homo floresiensis certain skeletal traits reappeared in this lineage that were seen in earlier australopith species but lost prior to or at the origin of Homo erectus. If the latter is true, then Homo floresiensis was descended from a species such as Homo habilis for which there is no evidence elsewhere in Asia. Only additional fossils or analyses will determine the evolutionary history of the "Hobbit" of Flores island.
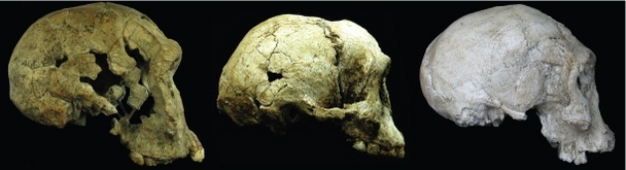
Figure 5: The LB1 cranium (center) compared to Homo habilis from Kenya (1.9 Ma) (left) and the slightly younger Homo erectus cranium from the Republic of Georgia (1.8 Ma) (right).
Previous investigations of the shape of the LB1 neurocranium indicate close similarities to this Georgian Homo erectus individual.
© 2012 Nature Education Photo courtesy of Karen Baab, Peter Brown, and the Kenya National Museums. All rights reserved.
Glossary
ancestral trait: Characteristic shared between two or more groups inherited from an ancestor prior to their most recent common ancestor
Australopithecus: An extinct hominin genus found only in Africa that preceded the genus Homo chronologically and contains several species spanning the time interval of 4.4 to 2.0 Ma
Australopithecus afarensis: An extinct hominin species in the genus Australopithecus from East Africa in the time period 3.9 to 3 Ma; the most famous representative of this species is the well-known fossil skeleton named "Lucy" from Ethiopia
brachycephalic: Term referring to skulls that are relatively wide from left to right compared to their length from front to back
cranial: Refers to the cranium (the bony skull minus the lower jaw)
CT: Acronym for computed tomography which is an X-ray-based imaging technique used to see both the external and internal anatomy of structures such as skulls and teeth (also known as CAT scanning in a clinical context)
derived trait: A characteristic shared between two or more groups that was inherited from their most recent common ancestor; this trait indicates a close evolutionary relationship among those groups that share the trait
hominin: Any member of the tribe Hominini, the group that includes modern humans and those fossil species that are more closely related to humans than any other animal
Homo: An extinct hominin genus that includes our own species (Homo sapiens) among others. Members of this genus first appear in the fossil record about 2.3 Ma in Africa. The Liang Bua fossils have been assigned to this genus.
Homo erectus: An extinct species of hominin that is in the same genus as modern humans (Homo) that was found in Africa, Eurasia and east Asia from 1.8 Ma to 0.5 or 0.05 Ma
Homo habilis: An extinct species of hominin that is in the same genus as modern humans (Homo) that was found in Africa from 2.3–1.4 Ma
island dwarfing: Phenomenon where large-bodied mammals may become dwarfed on after immigrating to an island
ka: Acronym referring to kiloannum (thousand years); used to express ages in thousands of years (e.g., a skull that is 20 ka is 20,000 years old)
lineage: A restricted segment of the phylogenetic tree that includes an ancestor and those species that it gave rise to, either directly or indirectly (can also refer to only a single species)
Ma: Acronym referring to megaannum (million years); used to express ages in millions of years (e.g., a skull that is 1.2 Ma is 1,200,000 years old)
mastoid fissure: A groove located on the underside of the skull between the mastoid process and the petrosal crest of the tympanic bone (common in skulls of Homo erectus)
morphology: Pertaining to the form or appearance of an organism or a particular anatomical structure
occipital torus: Elevated mound of bone that crosses the occipital bone located at the back of the skull (common in skulls of Homo erectus)
Oldowan: Oldest known stone tool technology (most common in Africa 1.6–1.6 Ma); flakes are produced by knocking them off of a core using a hammerstone
parietal bones: The large and fairly flat bones that surround the brain on the right and left sides; they are joined by the sagittal suture which traverses the midline of the skull on its top surface
pituitary fossa: A depression inside of the skull (on the sphenoid bone) where the pituitary gland (hypophysis) sits during life
Pleistocene: An epoch (a unit of geological time) that spans 1.8 Ma through 11.7 ka
sutures: A joint between bones of the skull; these are generally open during growth of the skull and begin to fuse later in life, after growth has ended
type specimen: The specimen for which a particular species is named for; intended to typify the characteristics of that species
Wallacea: Geographic region in Southeast Asia delimited by two imaginary lines: Wallace's Line to the west and Lydekker's line to the east. The region between these lines contains animals that are distinct from those in Asia (to the west) or Australia (to the east). See Figure 1.
References and Recommended Reading
Aiello, L.C. Five years of Homo floresiensis. American Journal of Physical Anthropology 142, 167-179 (2010).
Argue, D. et al. Homo floresiensis: a cladistics analysis. Journal of Human Evolution 57, 623-639 (2009).
Baab, K. L. Comparison of the LB1 neurocranium to extinct hominins, normal and pathological modern humans. American Journal of Physical Anthropology 141, 59 (2010).
Baab, K. L. & McNulty, K. P. Size, shape, and asymmetry in fossil hominins: the status of the LB1 cranium based on 3D morphometric analyses. Journal of Human Evolution 57, 608-622 (2009).
Brumm, A. et al. Hominins on Flores, Indonesia, by one million years ago. Nature 464, 748-752 (2010).
Boyages, S. C. et al. A comparative study of neurological and myxedematous endemic cretinism in western China. Journal of Clinical Endocrinology and Metabolism 67, 1262-1271 (1988).
Brown, P. et al. A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia. Nature 431, 1055-1061 (2004).
Brumm, A. et al. Early stone technology on Flores and its implications for Homo floresiensis. Nature 441, 624-628 (2006).
Falk, D. et al. The brain of LB1, Homo floresiensis. Science 308, 242 - 245 (2005).
Falk, D. et al. Brain shape in human microcephalics and Homo floresiensis. Proceedings of the National Academy of Sciences USA 7, 2513-2518 (2007).
Falk, D. et al. The type specimen (LB1) of Homo floresiensis did not have Laron Syndrome. American Journal of Physical Anthropology 140, 52-63 (2009).
Groves, C. & Fitzgerald, C. Healthy hobbits or victims of Sauron. HOMO - Journal of Comparative Biology 61, 211 (2010).
Halpern, J.-P. et al. The neurology of endemic cretinism. Brain 114, 825-841 (1991).
Henneberg, M. & Thorne, A. Flores may be pathological Homo sapiens. Before Farming 1, 2-4 (2004).
Hershkovitz, I. et al. Comparative skeletal features between Homo floresiensis and patients with primary growth hormone insensitivity (Laron syndrome). American Journal of Physical Anthropology 134, 198-208 (2007).
Huffman, O.F. et al. Provenience reassessment of the 1931-1933 Ngandong Homo erectus (Java), confirmation of the bone-bed origin reported by the discoverers. PaleoAnthropology, 1-60 (2010).
Indriati, E. et al. The age of the 20 meter Solo River terrace, Java, Indonesia and the survival of Homo erectus in Asia. PLoS ONE 6, e21562.
Jacob, T. et al. Pygmoid Australomelanesian Homo sapiens skeletal remains from Liang Bua, Flores: population affinities and pathological abnormalities. Proceedings of the National Academy of Sciences USA 103, 13421-13426 (2006).
Jungers. W. L. et al. The hobbits (Homo floresiensis) were not cretins. American Journal of Physical Anthropology S48, 161 (2009).
Jungers, W. L. et al. The foot of Homo floresiensis. Nature 459, 81-84 (2009).
Jungers, W. & Baab, K. The geometry of hobbits: Homo floresiensis and human evolution. Significance 6, 159-164 (2009).
Köhler, M. & Moyà-Solà, S. Reduction of brain size and sense organs in the fossil insular bovid Myotragus. Brain Behavior and Evolution 63, 125-140 (2004).
Koplik, H. The Diseases of Infancy and Childhood. New York, NY: Lea & Febiger, (1 918).
Larick, R. et al. Early Pleistocene 40Ar/39Ar ages for Bapang Formation hominins, Central Jawa, Indonesia. Proceedings of the National Academy of Sciences USA 98, 4866-4871 (2001).
Larson, S. G. et al. Descriptions of the upper limb skeleton of Homo floresiensis. Journal of Human Evolution 57, 555-570 (2009).
Martin, R. D. et al. Comment on "The Brain of LB1, Homo floresiensis." Science 312, 999 (2006).
Meijer, H. J. M. et al. The fellowship of the hobbit: the fauna surrounding Homo floresiensis. Journal of Biogeography 37, 995-1006 (2010).
Moore, M. W. & Brumm, A. Homo floresiensis and the African Oldowan. In Hovers, E. & Braun, D.R. (Eds.) Interdisciplinary Approaches to the Oldowan. Springer: New York, 61-69 (2009).
Obendorf, P. J. et al. Are the small human-like fossils found on Flores human endemic cretins? Proceedings of the Royal Society B: Biological Sciences 275, 1287-1296 (2008).
Oxnard, C. et al. Post-cranial skeletons of hypothyroid cretins show a similar anatomical mosaic as Homo floresiensis. PLoS ONE 5, e13018 (2010).
Tocheri, M. W. Derived wrist anatomy in the genus Homo as evidenced by 3D quantitative analyses: the implications for understanding the evolution of stone tool behaviors in hominins. American Journal of Physical Anthropology 132, 232-322 (2007).
Van den Bergh, G.D. et al. The youngest Stegodon remains in Southeast Asia from the Late Pleistocene archaeological site Liang Bua, Flores, Indonesia. Quaternary International 182, 16-48 (2008).
Weston, E. M. & Lister, A. M. Insular dwarfism in hippos and a model for brain size reduction in Homo floresiensis. Nature 459, 85-88 (2009).
Westway, K. E. et al. Reconstructing the geomorphic history of Liang Bua, Flores, Indonesia: a stratigraphic interpretation of the occupational environment. Journal of Human Evolution 57, 465-483 (2009).










