
Abstract
Recent findings have shown an inverse association between circulating C15:0/C17:0 fatty acids with disease risk, therefore, their origin needs to be determined to understanding their role in these pathologies. Through combinations of both animal and human intervention studies, we comprehensively investigated all possible contributions of these fatty acids from the gut-microbiota, the diet, and novel endogenous biosynthesis. Investigations included an intestinal germ-free study and a C15:0/C17:0 diet dose response study. Endogenous production was assessed through: a stearic acid infusion, phytol supplementation, and a Hacl1−/− mouse model. Two human dietary intervention studies were used to translate the results. Finally, a study comparing baseline C15:0/C17:0 with the prognosis of glucose intolerance. We found that circulating C15:0/C17:0 levels were not influenced by the gut-microbiota. The dose response study showed C15:0 had a linear response, however C17:0 was not directly correlated. The phytol supplementation only decreased C17:0. Stearic acid infusion only increased C17:0. Hacl1−/− only decreased C17:0. The glucose intolerance study showed only C17:0 correlated with prognosis. To summarise, circulating C15:0 and C17:0 are independently derived; C15:0 correlates directly with dietary intake, while C17:0 is substantially biosynthesized, therefore, they are not homologous in the aetiology of metabolic disease. Our findings emphasize the importance of the biosynthesis of C17:0 and recognizing its link with metabolic disease.
Similar content being viewed by others

Introduction
Recent findings have shown a negative association between circulating odd chain fatty acids (OC-FAs); pentadecanoic acid (C15:0) and heptadecanoic acid (C17:0), with metabolic disease risk1,2. Therefore, the determination of factors that affect their levels will be important in understanding the aetiology of metabolic disease, and could provide novel mechanistic insight into possible treatment routes.
According to the literature, the origin of C15:0 and C17:0 has long been attributed to the diet3,4, specifically from ruminant fat as the main contributor in a typical Western diet. This has been explained by the fact that these two OC-FAs are produced by the rumen microbiome5 and then incorporated into the fat deposits of the host animal destined for human consumption. All the current evidence for the dietary source of OC-FAs are based on dietary-assessment-correlation studies1,4,6,7. However, unambiguous proof of an exclusive dietary association is still lacking and there have been no investigations into the contributions from non-ruminant gut microbiota or from biosynthesis; such as α-oxidation. Interestingly, in different epidemiological studies that showed clear associations of OC-FAs with a reduced risk of metabolic disease1,2,8 there have been considerable differences in disease risk between C15:0 and C17:0, with C17:0 having the strongest inverse association, indicating different affects/effects.
We used a the combination of both different animal and human studies we are able for the first time to comprehensively study the many aspects of circulating OC-FAs.
Study Design
In each of the studies the appropriate ethical approval was obtained (see Supplementary material; appendix 1–9), where necessary signed informed consent was achieved, and all experimental protocols were approved by a named institutional and/or licensing committee. Additionally, all methods were performed in accordance with the relevant guidelines and regulations.
Several animal models were utilised in this manuscript; including rodent and canine. The rodent models were used to investigate the dietary and biochemical interactions/mechanisms and it has been previously published that a rodent is a suitable animal model for these investigations. The canine models was used due to the sample volume requirements of the study and it has been previously shown that dogs are a suitably comparable model to human glucose intolerance pathology. The animals were maintained under standard laboratory conditions with water ad libitum throughout the experiments. Animal body weight, food intake and health were monitored throughout the study. All samples were stored at sub −20 °C until analysed by gas chromatography/direct infusion with mass spectrometry detection (see Supplementary appendix 16 and 17).
Germ-free study
Four groups of mice were used to asses gut microbiota influences on the host circulating OC-FAs; two groups were kept on a chow diet (n = 8/group) and two groups on a high-fat diet (n = 6–7/group), from each subset there were both conventional or germ-free groups. Blood was collected after 75–84 days and processes into plasma/serum. All experiments were performed with permission 91–493 of the French Veterinary Services and in accordance to the European Community rules of animal care, and in compliance with Directive 86/608/EEC on the protection of animals used for scientific purposes and recommendation 2007/526/EC of the European Commission. Full detail are shown in Supplementary appendix 11 and 12.
Dose response study
Sprague-Dawley rats (Harlan, Indianapolis, USA) were overfed using one of five experimental diets (n = 6–9/group) at 17% above matched growth as previously described9. The fatty acid composition of each diet and the dietary sources of the lipids are shown in Supplementary appendix 13. Blood was collected after 21 days and processed into serum. All experimental procedures were ethically approved by the Institutional Animal Care and Use Committee at the University of Arkansas for Medical Science.
Phytol supplementation study
C57BL/6 mice (n = 4–5/group) were used in a phytol supplementation investigation10. Mice were fed ad libitum the rodent diet (Harlan Teklad, USA). The phytol supplemented group received an additional 0·5% (w/w) phytol added to the diet. After 14 days, blood was drawn and separated into serum. All experiments were executed according to accepted criteria for the humane care and experimental use of laboratory animals. All protocols were approved by the Animal Care and Use Committee of the University of Oulu.
Dairy fat supplementation study
Healthy human subjects with stable body weight and no family history of diabetes (n = 26) were recruited for an overfeeding intervention study conducted by Alligier and colleagues11, where samples were analysed and reported in this manuscript. As previously described11, the subjects (age 33 ± 1 years, body weight 79.1 ± 1.8 kg and fat mass 19.6 ± 0.8%) followed a 56 day overfeeding intervention, where they ate an additional 760 kcal/day. The additional energy intake was derived from 100 g of cheese, 20 g of butter, and 40 g of almonds. During the study, the subjects maintained their usual eating and physical activity behaviour (monitored by questionnaires and accelerometers over three five-day periods). After an overnight fast at day zero and day 56, blood was drawn and processed into plasma. All the participants gave signed informed consent following the explanation of the experimental protocol. The protocol was approved by the ethics committee of Lyon Sud-Est according to the French ‘Huriet-Serusclat’ law and the Second Declaration of Helsinki (study registration number NCT00905892; 20th May 2009, www.clinicaltrials.gov).
Controlled diet study
Samples analysed from the publication by Chiu and colleagues12. Results from a subset of 64 participants (23 men and 41 women) were used for the present analyses (see Supplementary appendix 14 for participant eligibility and exclusion criteria).
The participants consumed a baseline diet for four weeks, after which they were randomly assigned to one of two experimental diets (high or low dairy-fat diet) for a further four weeks, the macronutrient composition of the baseline and the experimental diets are shown in Supplementary appendix 15. High-, low-, non-fat dairy products and safflower oil were used to vary fatty acid compositions but maintain total fat content. Body weight was measured weekly and, if needed, energy intake was adjusted to achieve stable weight. All diets met the recommended daily intake for vitamins and minerals. Diet compliance was scored through weekly assessments by the dietitian using interviews, menu checklists, and food receipts. Blood was collected at baseline and at the end of the experimental diets after an overnight fast where it was processed into plasma. Signed informed consent was provided and the protocol was reviewed and approved by the institutional review boards of Children’s Hospital & Research Center at Oakland (dba University of California San Francisco Benioff Children’s Hospital Oakland) and the University of California, San Francisco (study registration number NCT01404897; 22nd July 2011, www.clinicaltrials.gov).
Stearic acid (C18:0) infusion study
As described previously13, Sprague-Dawley rats (n = 11/group) received a C18:0 (250 or 1000 nmol/kg/day) intraperitoneal infusion lasting five weeks, control animals received saline only. Blood was collected after 35 days and processed into serum. The animal study was approved by the Animal Care and Use Committee of Taichung Veterans General Hospital, Taiwan.
2-hydroxyacyl-CoA lyase 1 (Hacl1) study
Hacl1+/− Swiss Webster mice were mated to obtain Hacl1 deficient mice, as described previously14. Wildtype littermates were used as the control group (n = 5–7/group). Mice were fed a regular chow diet. After 14 weeks blood was collected and processed into plasma. All animal experiments were approved by the Institutional Animal Ethical Committee of KU Leuven.
Glucose intolerance study
Healthy adult mongrel dogs (~9 months of age, starting weight is 20.5 ± 2.9 kg, n = 5) were fed a meat based 52% high fat diet (TestDiet, Indiana, United States of America) for four to eight weeks15. Before and after the high fat diet intervention a 180 minute oral glucose tolerance test (OGTT) (0.9 g/kg body weight of glucose polymer administered) was carried out following a 24 hour fast; during each OGTT plasma samples were collected ten minutes before the glucose administration for lipid analysis. The baseline plasma samples were used for fatty acid analysis. The protocol was approved by the Vanderbilt University Institutional Animal Care and Use Committee, and the animals were housed and cared for according to Association for Assessment and Accreditation of Laboratory Animal Care guidelines.
Statistical Analysis
The statistical approaches utilised were appropriate for specific experimental designs; a paired t-test was used to asses significance/insignificance of an observational change that can be paired within a population, such as between an observation before and after an intervention within the same population (a paired t-test was used in the dairy fat supplementation study; see Table 1). A homoscedastic t-test was used between two unrelated equal variance groups to identify any significance/insignificance between them two groups (a homoscedastic t-test was used in the following studies: phytol supplementation study, controlled diet study, stearic acid (C18:0) infusion study, and the 2-hydroxyacyl-CoA lyase 1 (Hacl1) study. For both the paired and homoscedastic t-tests a value of p ≤ 0.05 was considered significant). A goodness of fit of a trendline expressed as R squared (R2) that is greater than 0.75 was considered a strong fit, an R2 greater than 0.5 was considered moderate. The area under the curve for the glucose intolerance was calculated via the trapezoidal rule, total area under the curve is the sum of all individually calculated trapezoid areas; individual trapezoid area equals (((y value two + y value one)/2)*(x value two − x value one)).
Results
Dose response study
Five groups of rats were subjected to isocaloric high-fat diets, where the composition of the fat in each diet had equal increments of beef tallow from 0% to 11·7% (see Supplementary appendix 13). To validate these being legitimate biomarkers of ruminant fat intake, we compared C15:0 and C17:0 compositions between the diets and the serum levels.
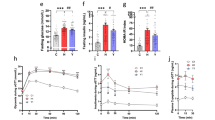
Increases in C15:0 in the diet linearly correlated (R2 = ~1) with serum levels (see Fig. 1). Although C17:0 levels increased, there was not a strong linear response suggesting other influencing factors which do not affect C15:0. Additionally, the smallest difference between C17:0 in each diet is between groups one and two, where there is actually the largest difference in C17:0 levels detected in the serum.

The serum samples were analysed by gas chromatography with mass spectrometry detection. C15:0 levels between diet and blood serum composition is highly correlated, with an R2 = 0·997. Error bars represent ± standard error of the mean. (n = 6–9/group).
This divergence between these OC-FAs is supported in published literature4,16,17. It has been proposed that C17:0 can also be biosynthesised by α-oxidation18; where phytanic acid is the primary target substrate (derived from diet or phytol metabolism19). If C17:0 is biosynthesised through α-oxidation, then any increase in the target substrate; phytanic acid, could change the in vivo C17:0 levels by increasing or inhibiting the rate of C17:0 biosynthesis20. (Phytol and phytanic acid dietary composition shown in Supplementary appendix 19).
Phytol supplementation study
To directly investigate upregulation or inhibition of C18:0 α-oxidation, we conducted a phytol supplementation study in mice, which is the target substrate for this oxidation pathway. The fatty acids were compared between the groups to assess any influences of the supplementation on the levels of serum OC-FAs.
The levels of C17:0 decreased in the phytol group compared with the control (control: 0·130 ± 0·005 Mol%, phytol: 0·089 ± 0·004 Mol%, homoscedastic t-test between the control and phytol groups; p < 0·001). There was no significant difference seen between the two groups for C15:0 (control: 0·049 ± 0·002 Mol%, phytol: 0·045 ± 0·002 Mol%, homoscedastic t-test between the control and phytol groups; p = 0·174).
Dairy fat supplementation study
The fatty acid composition of the dietary supplements are shown in Supplementary appendix 20. The plasma C15:0 and C17:0 fatty acid composition are shown in the table below (see Table 1).
Due to the human diet supplementation, the levels of C15:0 increased ~10%. However, C17:0 did not increase from baseline. Dietary assessment showed that the participants maintained their habitual diet alongside the supplementation and that the overfeeding intervention led to a substantial increase in ruminant fats (see Supplementary appendix 21).
Controlled diet study
The OC-FAs and the substrates involved in α-oxidation were inferred in the diets: baseline, low and high dairy fat diets (see Supplementary appendix 22). The experimental diets varied from the baseline diet by 82% to 152% for C15:0, 78% to 123% for C17:0, increased in phytol by 42% in both diets and phytanic acid varied by 102% to 226% from the baseline diet.
The participants had plasma collected after the baseline diet and again after the experimental diets, the plasma fatty acid changes due to the experimental diet are shown below (see Table 2).
In the high dairy fat diet there was a ~52% increase in C15:0 which caused a ~22% increase in the plasma C15:0. This directly reflects the expected increase by the diet according to the dose response study (see Fig. 1). The low dairy fat diet had ~18% decrease in C15:0 which in turn caused a ~30% decrease in the participant plasma composition; a larger decrease than expected.
The food products used to vary the C15:0 and C17:0 composition of the experimental diets were selected to assess the biomarker response, whilst reducing any risk of pathology to the participants and maintaining palatability over the four week period. Therefore in the high dairy fat diet, milk and cheese were used rather than ruminant tallow. The selected food products have a slightly higher C15:0 to C17:0 composition resulting in a greater overall intake of C15:0. Even though there was a greater C15:0 intake, there was also a significant increase in dietary C17:0, which represented a greater than normal dietary variation; this should have been sufficient to give a significant increase in C17:0 if it was a true dietary biomarker of intake, however, the circulating C17:0 composition decreased across both the experimental diets. To account for this decrease in serum C17:0 we compared the dietary phytol, phytanic acid, and, C17:0 (see Supplementary appendix 22).
Stearic acid (C18:0) infusion study
The results above suggest that α-oxidation could be important in the biosynthesis of C17:0 but not for C15:0. To investigate if C18:0 (the precursor of C17:0 from α-oxidation) could increase circulating C17:0 we conducted a controlled intraperitoneal infusion of C18:0 in rats and measured the absolute change in C17:0. While there was no significant change in the levels of C15:0 (control: 3·308 ± 0·310 μmol, 250 nmol/kg/day stearic acid: 4·447 ± 0·466 μmol, homoscedastic t-test p = 0·055), there was a significant increase in C17:0 (control: 5·683 ± 0·542 μmol, 250 nmol/kg/day stearic acid: 8·101 ± 0·651 μmol, homoscedastic t-test p < 0·001).
A higher concentration (1000 nmol/kg/day) of infused C18:0 did not significantly increase C17:0 further (p = 0.810), which suggests that C18:0 does not directly increase the rate of α-oxidation on straight chain fatty acids and that C18:0 is just an untargeted substrate.
2-hydroxyacyl-CoA lyase 1 (Hacl1) study
To further investigate OC-FAs biosynthesis via α-oxidation we examined a Hacl1−/− mouse model. This gene is responsible for catalysing a carbon-carbon cleavage reaction, removing formyl-CoA from 2-hydroxy-fatty-acyl-CoA to produce a fatty aldehyde that is shorter in chain length by one carbon unit21.
The Hacl1 gene knockout mouse model caused a reduction in C17:0 by 32·5% from the control group (control: 0·200 ± 0·022 Mol%; Hacl1−/−: 0·135 ± 0·008 Mol%, homoscedastic t-test p = 0·017). There was no significant difference in any other fatty acid measured (for C15:0: control: 0·023 ± 0·006 Mol%, Hacl1−/−: 0·016 ± 0·001 Mol%, homoscedastic t-test p = 0·176).
Glucose intolerance study
The relationship between C15:0 and C17:0 with glucose intolerance has been reported through a number of epidemiology studies1,8. Any variation in circulating C15:0 and C17:0 has been attributed to variations in ruminant fat intake with no consideration of biosynthesis. Therefore, we investigated how these OC-FAs correlate with the development of glucose intolerance during a controlled high fat diet in a canine model; the canine model was selected due to the sample volume requirements of the study and it has been previously shown that dogs are a suitably comparable model to human glucose intolerance pathology.
The glucose and insulin response to an OGTT was determined at baseline and again at the end of a glucose intolerance inducing diet. The difference between the canine baseline OGTT results and the subsequent results were compared to the baseline plasma C15:0 and C17:0 levels (see Fig. 2). Across the intervention, the dogs gained an average of 4.6 ± 1.3 kg.

Glucose and insulin were measured from 0 to 180 minutes and the area under the curve (AUC) was calculated using the trapezoidal rule. The graph data points represent the difference between the insulin AUC and the glucose AUC, and from the beginning to the end of the four to eight week high fat diet in relation to the fatty acid levels at baseline (Mol%). Top left: INSULIN (Ins.) - C15:0 R2 = 0·053, Top right: INSULIN (Ins.) - C17:0 R2 = 0·957. Bottom left: GLUCOSE (Glc.) - C15:0 R2 = 0·048, Bottom right: GLUCOSE (Glc.) - C17:0 R2 = 0·755. (n = 5).
When comparing other fatty acid levels from baseline with the progression of glucose intolerance in the dogs, only palmitic acid (C16:0) showed a strong positive linear correlation (glucose R2 = 0·891, insulin R2 = 0·818). This relationship between C16:0 and glucose intolerance has been previously reported in other studies22,23 and enforces the validity of this study.
Discussion
This is the first report that combines the results of different animal and human studies to comprehensively investigate the factors that affect the circulating levels of C15:0 and C17:0.
Firstly, we found there were no differences related to the presence or absence of gut microbiota on the mouse circulating OC-FA levels (see Supplementary appendix 18), and hence we can conclude that there was no evidence to suggest that the gut microbiota has an impact on circulating OC-FAs in the mice.
We conducted a five stage ruminant-fat dose response study in rats to assess the relationship between intake of C15:0 and C17:0 with their in vivo circulating levels. The diet resulted in a beef-tallow composition varying from 0% to 11·7% by increments of 2·7%, which represent normal ranges of dairy fat intake in typical Western countries. Circulating C15:0 levels directly correlated with intake (see Fig. 1), however, C17:0 does not show the same linear relationship suggesting other influencing factors. There is increasing evidence that C17:0 can be biosynthesised in vivo, which could account for its non-dietary origin24,25.
Several in vitro investigations have suggested that C17:0 is produced endogenously both through elongation of propionyl-CoA26 and alternatively, through α-oxidation of C18:021,27,28. However, the results for the C17:0 biosynthesis through propionyl-CoA do not directly explain tissue C17:0 levels in whole organisms, since the comparison between the expected and the actual fatty acid levels do not match, which suggests that propionyl-CoA plays a minor role. C17:0 could be biosynthesised through α-oxidation of C18:0, which was investigated in the subsequent studies.
The target substrate for α-oxidation is phytanic acid has been shown to competitively inhibit C17:0 biosynthesis (see the phytol supplementation study). As shown in Supplementary appendix 19 we investigated the diet composition of phytol and phytanic acid, comparing these for each diet in the ruminant-fat dose response study in rats. The dietary combination of phytanic acid and the phytol had a direct inverse correlation with the changes in the plasma C17:0 (as dietary phytanic acid and phytol increases the plasma C17:0 decreases proportionately), which is reinforced by the results in the mouse phytol supplementation study, and explains why the smallest difference between C17:0 in each diet is between diet groups one and two, where there is actually the largest difference in C17:0 levels in plasma. The correlations between circulating C17:0 and the dietary phytol/phytanic acid in the diet are stronger than with the dietary contributions of C17:0 which suggests that C17:0 is not a direct ruminant-fat biomarker but actually a product of biosynthesis regulated by dietary substrate intake.
In the human dairy fat supplementation study the plasma C15:0 level increased by ~10% (p = 0·031) as a result of a dietary 30% increase, this directly concurs with the expected increase according to the ruminant-fat dose response study (see Fig. 1). However, the plasma C17:0 levels did not change across the supplementation study; agrees with previous findings29. This can be explained by the levels of phytanic acid and its precursor phytol. The human supplementation included an increase of phytol by ~25% and an increase of phytanic acid by ~100%, which both reduces the biosynthesis of C17:0. This reduction in the biosynthesis along with the increased dietary consumption of C17:0 accounts for no change seen in the plasma.
To further investigate this, we performed analyses in a controlled, high or low dairy fat diet intervention following a baseline diet in healthy volunteers. For the low dairy fat diet, the C15:0 levels decreased as expected and increased for the high dairy fat intervention participants. The percentage increase and decrease of C15:0 in either of the experimental diets agrees with the expected changes, according to the linear relationship seen in figure one further reinforcing the biomarker relationship of C15:0. The changes in the plasma C17:0 can be explained by the variation in the phytol and phytanic acid in the diet. In the experimental diets there was an increase in phytol by 42% and phytanic acid varied by ~100% to ~230% from the baseline diet. The increases in the target substrates of α-oxidation considerably decreases C17:0 biosynthesis via α-oxidation by competitive inhibition, even when there was an increase in ruminant-fat.
Since it is well understood that C18:0 can be the substrate for C17:0 when studied in vitro, we conducted a C18:0 infusion study in rats to assess the effect on in vivo C17:0 levels. The absolute serum quantities of C17:0 increased significantly by ~70% (p < 0·001) in the rats receiving the C18:0 infusion compared to a control rat group. Further increasing the infusion of C18:0 above 250 nmol/kg/day did not further increase the serum C17:0 levels. This suggests that C18:0 itself does not increase the rate of C18:0 α-oxidation.
We also showed that Hacl1 plays a substantial role in the biosynthesis of C17:0. Using the Hacl1 knockout mouse model, there was a 32·5% reduction in plasma C17:0 compared to wildtype mouse controls. There were no other differences in any other fatty acids measured. It is unlikely to expect a complete C17:0 deficiency since there are other salvage pathways that could compensate for a deficiency in Hacl1; such as hydroxylation by fatty acid 2-hydroxylase (FA2H) followed by oxidation by 2-hydroxy acid oxidase (HAO2), additionally, dietary C17:0 sources still contribute to circulating levels.
The significance of the different associations of C15:0 and C17:0 with glucose intolerance has been previously reported, however, no consideration for the factors affecting biosynthesis of C17:0 has been taken into account. We compared the baseline fatty acid levels with the change in glucose and insulin measured during an oral glucose tolerance test after a high fat diet in a suitable canine model; the canine model was selected due to the sample volume requirements of the study and it has been previously shown that dogs are a suitably comparable model to human glucose intolerance pathology. There was no correlation between C15:0 at baseline with the change in either glucose or insulin across the high fat diet, however, a strong correlation was seen with C17:0. These results highlight the importance of understanding the origin of these fatty acids and indicate that the biosynthesis (not the dietary source) of C17:0 has a strong relationship with the development of glucose intolerance, either as a marker of healthy lipid metabolism or indeed the fatty acid may have protective properties.
To summarise, it is clear that individual OC-FAs are the result of very different processes and that their relations with disease risk cannot be generalised. From our results we can conclude that C15:0 is a direct, linear biomarker of dietary C15:0 intake. C17:0 has been misidentified as a ruminant fat intake biomarker due to its relationship with dietary phytol, phytanic acid and C18:0. However, the primary relationship between diet and in vivo C17:0 levels is highly influenced by biosynthesis. Moreover, the disparity between C15:0 and C17:0 agrees with data collected in epidemiological studies showing that C17:0 has a stronger inverse association with diseases. This highlights the importance for further investigation into the biosynthesis of C17:0 and how these pathways relate to metabolic disease.
Additional Information
How to cite this article: Jenkins, B. J. et al. Odd Chain Fatty Acids: New Insights of the Relationship Between the Gut Microbiota, Dietary Intake, Biosynthesis and Glucose Intolerance. Sci. Rep. 7, 44845; doi: 10.1038/srep44845 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Forouhi, N. G. et al. Differences in the prospective association between individual plasma phospholipid saturated fatty acids and incident type 2 diabetes: the EPIC-InterAct case-cohort study. The Lancet Diabetes & Endocrinology, doi: 10.1016/S2213-8587(14)70146-9 (2014).
Khaw, K.-T., Friesen, M. D., Riboli, E., Luben, R. & Wareham, N. Plasma phospholipid fatty acid concentration and incident coronary heart disease in men and women: the EPIC-Norfolk prospective study. PLoS Med. 9, e1001255 (2012).
Smedman, A. E., Gustafsson, I.-B., Berglund, L. G. & Vessby, B. O. Pentadecanoic acid in serum as a marker for intake of milk fat: relations between intake of milk fat and metabolic risk factors. Am J Clin Nutr 69, 22–29 (1999).
Brevik, A., Veierød, M. B., Drevon, C. A. & Andersen, L. F. Evaluation of the odd fatty acids 15:0 and 17:0 in serum and adipose tissue as markers of intake of milk and dairy fat. Eur J Clin Nutr 59, 1417–1422 (2005).
Vlaeminck, B., Fievez, V., Cabrita, A. R. J., Fonseca, A. J. M. & Dewhurst, R. J. Factors affecting odd- and branched-chain fatty acids in milk: A review. Animal Feed Science and Technology 131, 389–417 (2006).
Wolk, A., Vessby, B., Ljung, H. & Barrefors, P. Evaluation of a biological marker of dairy fat intake. Am J Clin Nutr 68, 291–295 (1998).
Sofie Biong, A., Berstad, P. & Pedersen, J. I. Biomarkers for intake of dairy fat and dairy products. European Journal of Lipid Science and Technology 108, 827–834 (2006).
Yakoob, M. Y. et al. Circulating biomarkers of dairy fat and risk of incident stroke in US men and women in 2 large prospective cohorts. Am J Clin Nutr ajcn. 083097, doi: 10.3945/ajcn.114.083097 (2014).
Ronis, M. J. J. et al. Medium chain triglycerides dose-dependently prevent liver pathology in a rat model of non-alcoholic fatty liver disease. Exp Biol Med (Maywood) 238, 151–162 (2013).
Selkälä, E. M. et al. Phytol is lethal for Amacr-deficient mice. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids 1851, 1394–1405 (2015).
Alligier, M. et al. Subcutaneous Adipose Tissue Remodeling during the Initial Phase of Weight Gain Induced by Overfeeding in Humans. The Journal of Clinical Endocrinology & Metabolism 97, E183–E192 (2012).
Chiu, S. et al. Diets High in Protein or Saturated Fat Do Not Affect Insulin Sensitivity or Plasma Concentrations of Lipids and Lipoproteins in Overweight and Obese Adults. J. Nutr. 144, 1753–1759 (2014).
Pan, P.-H. et al. Stearic acid attenuates cholestasis-induced liver injury. Biochemical and Biophysical Research Communications 391, 1537–1542 (2010).
Mezzar, S., Baes, M. & Van Veldhoven, P. P. Hacl1−/− mice, a new animal model for α-oxidation deficiency. Chemistry and Physics of Lipids 163, S23 (2010).
Coate, K. C. et al. Hepatic glucose uptake and disposition during short-term high-fat vs. high-fructose feeding. American Journal of Physiology - Endocrinology and Metabolism 307, E151–E160 (2014).
Baylin, A. et al. Fasting Whole Blood as a Biomarker of Essential Fatty Acid Intake in Epidemiologic Studies: Comparison with Adipose Tissue and Plasma. Am. J. Epidemiol. 162, 373–381 (2005).
Nestel, P. J. et al. Specific plasma lipid classes and phospholipid fatty acids indicative of dairy food consumption associate with insulin sensitivity. Am J Clin Nutr 99, 46–53 (2014).
Takahashi, T., Takahashi, H., Takeda, H. & Shichiri, M. Alpha-oxidation of fatty acids in fasted or diabetic rats. Diabetes Research and Clinical Practice 16, 103–108 (1992).
Van Veldhoven, P. P. Biochemistry and genetics of inherited disorders of peroxisomal fatty acid metabolism. J Lipid Res 51, 2863–2895 (2010).
Cornish-Bowden, A. Fundamentals of Enzyme Kinetics (John Wiley & Sons, 2013).
Foulon, V. et al. Breakdown of 2-Hydroxylated Straight Chain Fatty Acids via Peroxisomal 2-Hydroxyphytanoyl-CoA Lyase a Revised Pathway for the Α-Oxidation of Straight Chain Fatty Acids. J. Biol. Chem. 280, 9802–9812 (2005).
Mozaffarian, D. Saturated fatty acids and type 2 diabetes: more evidence to re-invent dietary guidelines. The Lancet Diabetes & Endocrinology 2, 770–772 (2014).
Ma, W. et al. Prospective association of fatty acids in the de novo lipogenesis pathway with risk of type 2 diabetes: the Cardiovascular Health Study. Am J Clin Nutr 101, 153–163 (2015).
Mead, J. F. & Levis, G. M. A 1 Carbon Degradation of the Long Chain Fatty Acids of Brain Sphingolipids. J. Biol. Chem. 238, 1634–1636 (1963).
Wakil, S. Lipid Metabolism (Elsevier, 2012).
Weitkunat, K. et al. Effects of dietary inulin on bacterial growth, short-chain fatty acid production and hepatic lipid metabolism in gnotobiotic mice. The Journal of Nutritional Biochemistry 26, 929–937 (2015).
Croes, K., Foulon, V., Casteels, M., Veldhoven, P. P. V. & Mannaerts, G. P. Phytanoyl-CoA hydroxylase: recognition of 3-methyl-branched acyl-CoAs and requirement for GTP or ATP and Mg2+ in addition to its known hydroxylation cofactors. J. Lipid Res. 41, 629–636 (2000).
Guo, L., Zhou, D., Pryse, K. M., Okunade, A. L. & Su, X. Fatty Acid 2-Hydroxylase Mediates Diffusional Mobility of Raft-associated Lipids, GLUT4 Level, and Lipogenesis in 3T3-L1 Adipocytes. J Biol Chem 285, 25438–25447 (2010).
Benatar, J. R. & Stewart, R. A. The effects of changing dairy intake on trans and saturated fatty acid levels- results from a randomized controlled study. Nutrition Journal 13, 32 (2014).
Acknowledgements
The authors are grateful to the Medical Research Council for core funding (Lipid Profiling and Signalling programme grant; number UD99999906, Cambridge Lipidomics Biomarker Research Initiative; grant G0800783, MRC Human Nutrition Research PhD programme). Grant GAČR: GA15–09518S and grant Czech Science Foundation GACR: 16-06326S funded part of the gut microbiota investigation. The authors would like to acknowledge the USDA (ACNC-USDA-CRIS 6251-51000-005-03S) for funding of the dose response animal study within this manuscript. The Human study “Dairy Fat supplementation” was supported by research grants from the Hospices Civils de Lyon (Actions Incitatives); from the Programme Hospitalier de Recherche Clinique Interregional; from the Agence Nationale de la Recherche (Programme de Recherche en Nutrition Humaine and the Programme National de Recherche en Alimentation); and from the Innovation Stratégique Industrielle program of the Agence pour l’Innovation OSEO (Innovation Thérapeutique – Diabète project). K. Seyssel and M. Alligier were recipients of a doctoral fellowship from the Ministère de l’Enseignement Supérieur et de la Recherche (France). The phytol supplementation animal study was supported by grants from the Academy of Finland (138690), the Sigrid Juselius Foundation and NordForsk under the Nordic Centres of Excellence Programme in Food, Nutrition and Health, project “Mitohealth” (070010). The NIH Grant R01-DK-18243 for funding of the canine study. HACL1 knockout mouse model was supported by grants from the Flemish “Fonds Wetenschappelijk Onderzoek” (G.0721.10N) and KU Leuven (OT/14/100).
Author information
Authors and Affiliations
Contributions
B.J.J. designed and conducted the research, analysed samples & data and wrote & designed the manuscript. K. Se, S.C., P.H.P., S.Y.L., W.V., K.J.A., K.H., S.H., R.S., C.J.C., M.A., M.L., M.M., G.K., A.C., S.K., R.M.K., E.D.S., P.P.V.V. and M.R. provided essential materials/data/samples and wrote the manuscript. E.S. designed/wrote analysis software. Z.A., J.A.W. and J.L.G. wrote the manuscript. K. Su. was responsible for preliminary work. A.K. is the PhD supervisor of B.J.J.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Jenkins, B., Seyssel, K., Chiu, S. et al. Odd Chain Fatty Acids; New Insights of the Relationship Between the Gut Microbiota, Dietary Intake, Biosynthesis and Glucose Intolerance. Sci Rep 7, 44845 (2017). https://doi.org/10.1038/srep44845
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep44845
This article is cited by
-
Daphnane diterpenoid orthoesters with an odd-numbered aliphatic side chain from Daphne pedunculata
Journal of Natural Medicines (2024)
-
Air pollution exposure and plasma fatty acid profile in pregnant women: a cohort study
Environmental Science and Pollution Research (2023)
-
Lipid signatures of chronic pain in female adolescents with and without obesity
Lipids in Health and Disease (2022)
-
A mouse model of gestational diabetes shows dysregulated lipid metabolism post-weaning, after return to euglycaemia
Nutrition & Diabetes (2022)
-
Targeting de novo lipogenesis and the Lands cycle induces ferroptosis in KRAS-mutant lung cancer
Nature Communications (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
