
Abstract
Bird fossils from Turonian (ca. 90 Ma) sediments of Axel Heiberg Island (High Canadian Arctic) are among the earliest North American records. The morphology of a large well-preserved humerus supports identification of a new volant, possibly diving, ornithurine species (Tingmiatornis arctica). The new bird fossils are part of a freshwater vertebrate fossil assemblage that documents a period of extreme climatic warmth without seasonal ice, with minimum mean annual temperatures of 14 °C. The extreme warmth allowed species expansion and establishment of an ecosystem more easily able to support large birds, especially in fresh water bodies such as those present in the Turonian High Arctic. Review of the high latitude distribution of Northern Hemisphere Mesozoic birds shows only ornithurine birds are known to have occupied these regions. We propose physiological differences in ornithurines such as growth rate may explain their latitudinal distribution especially as temperatures decline later in the Cretaceous. Distribution and physiology merit consideration as factors in their preferential survival of parts of one ornithurine lineage, Aves, through the K/Pg boundary.
Similar content being viewed by others

Introduction
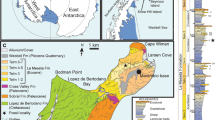
While insights have been gained into Mesozoic avian diversity in the southern high latitudes that bear on the origin of modern birds, Northern Hemisphere records are scarce. In the High Canadian Arctic, the Cretaceous is represented by sedimentary and volcanic rocks of the Sverdrup Basin1. The fossil locality reported here is from western Axel Heiberg Island near Expedition Fiord, 79° 23.5′N, 92° 10.9′W (Fig. 1). Amongst these rocks, the Late Cretaceous is represented by the marine shales of the Kanguk Formation, which overlie the Strand Fiord Formation flood basalts. The vertebrate fossil yielding layer lies between these two formations within a roughly 3-meter section of siltstones and shales2. The fossils represent a diverse assemblage including champsosaurs3, an array of fresh water fish including vidalamiines, amiines, lepisosteids, and teleosts4, at least 5 species of turtles5,6, rare juvenile elasmosaur teeth7 and the bird discussed below. These remains, while generally disarticulated, include partially articulated specimens suggesting limited transport2.

(a) Regional map of the High Canadian Arctic; field locality shown by the red star. Inset: Map of North America, box shows High Canadian Arctic. Map created with Generic Mapping Tools (GMT) (Version 5.1.1)54. (b) Stratigraphic column of the mapped units (modified from ref. 2, reprinted with permission by AAAS), fossil bed is located within siltstone layer between Strand Fiord Fm. and Kanguk Fm.
In addition to the diverse fossil assemblage of this locality, isotopic studies of foraminifera suggest extreme global warmth during the Turonian8,9,10,11. These studies indicate bottom water temperatures between 18 °C and 25 °C with surface waters as warm as 35 °C8,9,10. Available global circulation models with elevated CO2, hypothesized on the basis of a commensurate increase of global magmatism2, can reproduce some but not all of the observed Turonian polar warmth12,13,14. Other factors that may have contributed to Turonian polar warmth include reduced albedo resulting from expanding vegetation and increased radiative forcing from cloud coverage13,14.
The fossil record of birds in North America begins in the early Late Cretaceous, later than the beginning of their record in former parts of Eurasia and Gondwana. While there are substantial records, including partial to complete skeletons, from the early Cretaceous in Eurasia (e.g., La Huerguina, Jiufutang and Yixian Formations; ~120 Ma)15,16,17 the Gondwanan record begins in the Aptian and Albian (e.g., Crato and Wonthaggi Formations; ~115 Ma)18,19. The earliest articulated remains from North America date to the late Santonian-early Campanian interval (Mooreville Formation, Pierre Shale, Niobrara Formation; ~83 Ma)20,21,22,23,24,25,26.
Early Late Cretaceous records of North America are limited. The most abundant remains are isolated bones from the Middle Cenomanian (~95 Ma) Ashville Formation of Saskatchewan, Canada27,28. Approximately coeval remains comprise several bones from the Greenhorn and Woodbine Formations, from Kansas and Texas respectively29,30,31. A nearly complete humerus of Icthyornis sp. was also recovered from the early Turonian Kaskapau Formation of Alberta, Canada32.
A tarsometatarsus of the foot propelled diving taxon Hesperornis has been reported from the Kanguk Formation of the Fosheim Penninsula on Ellesmere Island (79° 45′N, 83° 36′W)33. Additionally, avialan teeth referred to Hesperornithiformes have been reported from the Kanguk Formation on Devon Island (76° N, 91° W) by ref. 34. The maps in Fig. 2 show fossil bird localities of the Late Cretaceous above 25° northern latitude grouped by stage (the details of each locality can be found in Supplementary Information Table S1).

Maps showing the distribution of Late Cretaceous birds in the Northern hemisphere based by stage; colors correspond with assigned stage based on the following: Late Albian and Cenomanian (reconstructed paleopositions shown for 100 Ma), magenta; Turonian (92 Ma), red; Coniacian (90 Ma), green; Santonian (86 Ma), yellow; Campanian (84 Ma), orange; and Maastrichtian (72 Ma), blue.
Symbols with multiple colors denote localities with age uncertainties that span across multiple stages. Fossil localities and their corresponding plates have been reconstructed to their paleopositions using plate models and rotation data presented in ref. 55. Circle symbols represent hesperornithiformes, triangles represent enantiornithes, and squares represent non-hesperornithiform ornithurines. The red star represents the locality shown in Fig. 1a. For reference, a 60° north latitude band is shown. Fossil distribution reconstructions were produced with GPlates (Version 1.5) and GMT (Version 5.1.1)54.
Results
Systematic Paleontology
Avialae Gauthier 198635
Ornithurae Haeckel 186636
Tingmiatornis arctica gen. et. sp. nov.
Etymology
The genus name is from “Tingmiat”, which in Inuktitut references “those that fly”. The species name makes reference to the high Arctic provenance of the holotype and referred material.
Holotype and referred specimens
The holotype is a complete left humerus (NUFV 1960; NUFV: Canadian Museum of Nature, Ottowa, Ontario, Canada; Fig. 3). A distal humerus (UR 00.200; UR: University of Rochester, Rochester, New York, U.S.A.; Fig. 4) and proximal ulna (NUFV 1838; Fig. 5) are here referred to the species.

The holotype specimen of Tingmiatornis arctica, NUFV 1960, a complete left humerus.
Photograph (left) and x-ray computed tomography images (right) of the element in caudal, proximal, cranial, and ventral views.

Distal left humerus referred to Tingmiatornis arctica, UR 00.200.
Photograph of the element in (left to right) cranial, dorsal, ventral, and caudal views.

Proximal right ulna referred to Tingmiatornis arctica, NUFV 1838.
Photograph (left) and x-ray computed tomography images (right) of the element in ventral, proximal, dorsal and caudal views.
Locality and horizon
Units exposed at the fossil locality, located on Expedition Fiord (79° 23.5′N, 92° 10.9′W), Axel Heiberg Island, Nunavut, Canada (Fig. 1), vary in lithology from siltstone at the top of the section to shale near the base. These units, containing the new avialan remains as part of a rich fossil vertebrate assemblage, occur in a thinly deposited (~3 m) layer directly underlying Kanguk shale and overlying the subaerially-erupted Strand Fiord Formation flood basalts. The fossil beds are exposed on opposing sides of a river-cut and extend for approximately 50 m. Radiometric data and stratigraphic constraints suggest an age of ~92 Ma (Turonian) for the fossil assemblage2, which includes champsosaurs, freshwater fish, turtles, and elasmosaurs3,4,5,6,7. Depositional characteristics suggest a large bay, whereas the presence of freshwater fish, turtles, and champsosaurs indicates a freshwater to brackish environment4. The paleolatitude for the locality during deposition of the fossil-bearing strata, based on paleomagnetic analyses of the Strand Fiord basalts, is ~71° N37.
Diagnosis
The new taxon is differentiated from Ichthyornis20 by numerous features including the more globose humeral head with significant caudal extent, a narrow deltopectoral crest and more strongly developed secondary pneumotricipital fossa. The bicipital crest in the new taxon is also more elongate, convex distally, and transitions smoothly into the humeral shaft distally, rather than at an angle approaching 90 degrees. The new taxon possesses a narrower, slightly cranially deflected deltopectoral crest that is significantly less than shaft diameter in dorsal extent. It is differentiated from the proposed hesperornithiform Pasquiaornis tankei [RSM (Royal Saskatchewan Museum, Regina, Saskatchewan, Canada) P2487.4] in its larger size as well as proportions of the humerus (Supplementary Table S2, measurements from ref. 27). The new taxon is also differentiated from Pasquiaornis in the relatively more globose dorsal condyle, the weakly-projected olecranon process and smaller bicipital tubercle on the ulna. It should be noted that all elements from Pasquiaornis tankei and Pasquiaornis hardiei are isolated and that forelimb elements did not comprise holotype material in either species27. However, they are similar to the new taxon in the narrow, elongate and slightly cranially-deflected deltopectoral crest.
Description
In the holotype specimen NUFV 1960 (Fig. 3), the humeral head is globose. It is more strongly projected and ovoid in proximal view with more extent on the caudal surface than in Ichthyornis dispar. Part of the surface has been slightly abraded. The dorsal tubercle is not clearly demarcated. The m. supracoracoideus insertion appears to have been at the narrow juncture of the proximal-most deltopectoral crest and head. The capital incisure is developed between the ventral tubercule and head. The dorsal margin of the pneumotricipital fossa is demarcated by both a distal ridge and thin muscular crest. This margin is more strongly demarcated than in Ichthyornis20 and a proximal humerus from the Maastrichtian Nemegt Formation IGM (Institute of Geology, Mongolian Academy of Sciences, Ulaanbaatar, Mongolia) 100/130938. A portion of the tricipital fossa appears to extend onto the ventral tubercle giving it a hood-like appearance in caudal view. The pneumotricipital fossa itself appears rather shallow and better developed due to crushing. It is unclear if it was perforate. The m. scapulohumeralis caudalis scar is visible ventral to the margin of the pneutotripital fossa. It faces caudally. In Ichthyornis, this scar faces ventrally and is positioned at the terminus of the very-abbreviate bicipital crest seen in that taxon and other basal ornithurines20. The bicipital crest in NUFV 1960 is more elongate, approximately 1/3 the length of the deltopectoral crest and curves smoothly into the shaft. The capital ridge is strongly developed and extensive down the shaft making the humerus triangular in cross section. The deltopectoral crest is long and narrow, extending 37 mm down the shaft, slightly less than 1/3 the length of the humerus. It appears to be slightly cranially-deflected. This deflection is visible both in proximal view and caudal view. The shaft does not appear to have been strongly curved.
The distal end of the holotype humerus is crushed craniocaudally. The flexor process is short. The dorsal supracondylar process is small and distally located, at the estimated level of the proximal end of the dorsal condyle. The distal surfaces of well-developed condyles are visible. The distal surface of the humerus is perpendicular to the shaft. A slight concavity on the caudodorsal edge of the distal humerus may be consistent with the presence of a m. scapulotriceps groove.
Referred specimen UR 00.200 (Fig. 4) comprises a distal left humerus. The specimen has a diameter of 13.4 mm and a mid-shaft diameter 8.45 mm and does not show much crushing. The cross section of the shaft does not appear to show significant osteosclerotic thickening; it appears to be comparable to the proposed volant and diving hesperornithiform Pasquaornis. The distal humeral shaft is curved, however, the degree of this curvature appears artifactual. Breaks in several places in the distal shaft have been filled with black epoxy resin. In caudal view, the edges of the brachial fossa are demarcated dorsally and ventrally. An angled distal ridge is located just proximal to the ventral condyle and angles dorsally. The flexor process is short. Muscular scars on the ventral supracondylar tubercle and ventral epicondyle are complex. The ventral epicondyle shows a large muscular pit. Just proximal and slightly ventral to this feature, a smaller round pit is developed that is topologically consistent with the m. pronator superficialis attachment in Aves. The facet-like scar of the collateral ligament, located just dorsal to it, has a cranially projected proximal margin. The dorsal condyle is subrounded and larger than the ventral. The dorsal supracondylar tubercle is flange-like and extends just proximal to the tip of the dorsal condyle. No muscle attachment (or origin/insertion) scars were discernable.
The referred right proximal ulna (NUFV 1838; Fig. 5) has a preserved length of 88.32 mm and appears to be missing, perhaps, a quarter of its length. The shaft is not strongly bowed. Its estimated length would be just slightly shorter than the complete humerus. A small bicipital tubercle is developed. The dorsal cotyla is not well exposed. The ventral cotyla extends proximally onto the base of the olecranon, which is abbreviate.
Two other possible avian elements were recovered from the same deposits, NUFV 1781 (UR 06.042) (Supplementary Fig. S1) and NUFV 1763 (UR 06.023), but these elements are either too damaged or too embedded in rock to identify to Tingmiatornis arctica with confidence.
Discussion
Fossils of Tingmiatornis arctica, while well-preserved, are few in number thus limiting inferences about the anatomy and ecology of this species. From the bone cortex thickness (Supplementary Table S3, Supplementary Figure S2) and proportions of the humerus, Tingmiatornis appears to be volant and likely a diving taxon like Pasquaornis27. However, given that these taxa are known from isolated primarily forelimb remains and known Hesperornithiformes preserve highly reduced forelimbs with few proposed diagnositic characters, the potential hesperornithiform affiinty of these taxa27 is supported by distributional data and diving ecology, an ecology that has been acquired multiple times in birds.
The Turonian Arctic locality, with calm water and the warm climate2,3,4 would offer birds like Tingmiatornis arctica ample suitable environments for both breeding and survival. However, direct evidence for breeding (e.g., nests or eggs) has not been recovered at the Axel Heiberg site. A correlation has also been found between larger avian body size, avian abundance, and increased abundance of carnivorous fish39,40. Based on complete skeletons of the bowfin, Amia calva41, as a most conservative estimate, the amiid fish known at the Arctic locality would be roughly 0.3–0.6 meters long. This suggests these fish may have been able to out-compete birds below the size of Tingmiatornis arctica, if, as we propose, Tingmiatornis arctica was a diving bird potentially feeding on the fish documented in the Axel Heiberg fossil assemblage4. That Tingmiatornis arctica foraged at night also cannot be excluded considering that one potential modern analogue, the Western Grebe (Aechmophorus occidentalis), is known to nocturnally forage while in its wintering habitats. If Tingmiatornis arctica lived in the Arctic during the winter months when the area (presently at 79°N latitude, but at ~71°N during the Cretaceous37), it would have experienced prolonged periods of twilight in addition to approximately 2 months of total darkness each year.
The discovery of Tingmiatornis arctica and the Axel Heiberg birds contributes to the earliest fossil record of birds from North America. While there are abundant records of birds from the Campanian and Maastrichtian, early Late Cretaceous remains (Coniacian, Turonian and earlier) are more limited. Interestingly, high latitude Campanian taxa of North America are ornithurines and not parts of more basally-divergent lineages42 (Fig. 2). A similar pattern has been observed in the high southern latitudes in the Campanian to Maastrichtian of Antarctica, although records are more limited43,44. Furthermore, lower latitude Patagonia during this interval shows a variety of more basal lineages such as parts of Enantiornithes (e.g., refs. 45, 46) as well as ornithurine lineages45,47.
Intriguingly, differences in physiology and growth rate may distinguish these close ornithurine relatives of crown clade Aves43,48,49. Two factors proposed to explain the distribution of ornithurines at high latitudes in the Campanian of North America included ecological and physiological explanations: 1. that ornithurines include more wading, swimming and diving forms than Enantiornithes and may have differentially exploited aquatic niches42,50; and 2., that a highly seasonal climate may have favored high growth rates such as those exhibited by Aves and some other ornithurines48. Of these factors, we note that high seasonality (i.e., presence/absence of freezing conditions) was most likely not an important factor for the occurrence of Tingmiatornis arctica given the extreme warmth of the Turonian Arctic2. In contrast, Tingmiatornis arctica is clearly associated with a freshwater aquatic environment, and this factor, in the context of the extreme warmth allowing establishment of the Axel Heiberg ecosystem, was probably most important for explaining its presence in the Turonian Arctic. However, some as yet unidentified physiological factors enabling rapid growth rates, foraging during periods of darkness or facilitating long-distance migration to lower latitudes may explain their occurance in high latitudes especially during cooler climates of the Campanian and Maastrichtian. While data on the high latitude distribution of Mesozoic birds remains limited, Tingmiatornis arctica and the Axel Heiberg birds may support hypotheses favoring a physiological and/or distributional explanation43 for the the preferential survivorship of some ornithurines, Aves, across the K/Pg boundary, ultimately resulting in the ~10,000 species of living birds seen today. Parts of Aves are known from the latest Cretaceous of mid-latitude regions38,42,51,52 and high latitude sites42,43,44,53.
Methods
Fossil preparation and photography was performed at the University of Rochester. X-ray computed tomography images were aquired at the High-Resolution X-ray Computed Tomography Facility of the University of Texas at Austin (UTCT) using a Bio-Imaging Research, Inc. Advanced Computed Tomography Inspection System (ACTIS) with a 450-kV tungsten x-ray source high-energy subsystem. Digital isolation of the fossils was done at the UTCT processing laboratory using Avizo and VG Studio Max software.
Additional Information
How to cite this article: Bono, R. K. et al. A Large Ornithurine Bird (Tingmiatornis arctica) from the Turonian High Arctic: Climatic and Evolutionary Implications. Sci. Rep. 6, 38876; doi: 10.1038/srep38876 (2016).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Balkwill, H. R. Evolution of Sverdrup Basin, Arctic Canada. AAPG Bull. 62, 1004–1028 (1978).
Tarduno, J. A. et al. Evidence for extreme climatic warmth from Late Cretaceous Arctic vertebrates. Science 282, 2241–2243 (1998).
Vandermark, D., Tarduno, J. A. & Brinkman, D. B. A fossil champsosaur population from the High Arctic: Implications for Late Cretaceous paleotemperatures. Palaeogr. Palaeocl. 248, 49–59 (2007).
Friedman, M., Tarduno, J. A. & Brinkman, D. B. Fossil fishes from the High Canadian Arctic: further palaeobiological evidence for extreme climatic warmth during the Late Cretaceous (Turonian–Coniacian). Cretaceous Res. 24, 615–632 (2003).
Brinkman, D. B. & Tarduno, J. A. A Late Cretaceous (Turonian–Coniacian) high-latitude turtle assemblage from the Canadian Arctic. Can. J. Earth Sci. 42, 2073–2080 (2005).
Vandermark, D., Tarduno, J. A., Brinkman, D. B., Cottrell, R. D. & Mason, S. New Late Cretaceous macrobaenid turtle with Asian affinities from the High Canadian Arctic: dispersal via ice-free polar routes. Geology 37, 183–186 (2009).
Vandermark, D., Tarduno, J. A. & Brinkman, D. B. Late Cretaceous plesiosaur teeth from Axel Heiberg Island, Nunavut, Canada. Arctic 59, 79–82 (2006).
Huber, B. T., Hodell, D. A. & Hamilton, C. P. Middle–Late Cretaceous climate of the southern high latitudes: stable isotopic evidence for minimal equator-to-pole thermal gradients. Geol. Soc. Am. Bull. 107, 1164–1191 (1995).
Huber, B. T., Norris, R. D. & MacLeod, K. G. Deep-sea paleotemperature record of extreme warmth during the Cretaceous. Geology 30, 123–126 (2002).
MacLeod, K. G., Huber, B. T., Berrocoso, Á. J. & Wendler, I. A stable and hot Turonian without glacial δ18O excursions is indicated by exquisitely preserved Tanzanian foraminifera. Geology 41, 1083–1086 (2013).
Berrocoso, Á. J. et al. The Lindi Formation (upper Albian–Coniacian) and Tanzania Drilling Project Sites 3640 (Lower Cretaceous to Paleogene): Lithostratigraphy, biostratigraphy and chemostratigraphy. J. Afr. Earth Sci. 101, 282–308 (2015).
Poulsen, C. J., Gendaszek, A. S. & Jacob, R. L. Did the rifting of the Atlantic Ocean cause the Cretaceous thermal maximum? Geology 31, 115–118 (2003).
Zhou, J., Poulsen, C. J., Rosenbloom, N., Shields, C. & Briegleb, B. Vegetation-climate interactions in the warm mid-Cretaceous. Clim. Past 8, 565–576 (2012).
Poulsen, C. J. & Zhou, J. Sensitivity of Arctic climate variability to mean state: insights from the Cretaceous. J. Climate 26, 7003–7022 (2013).
Hou, L. H., Zhou, Z., Martin, L. D. & Feduccia, A. A beaked bird from the Jurassic of China. Nature 377, 616–618 (1995).
Qiang, J., Chiappe, L. M. & Shu’An, J. A new late Mesozoic confuciusornithid bird from China. J. Vertebr. Paleontol. 19, 1–7 (1999).
Chiappe, L. M., Norell, M. & Clark, J. A new skull of Gobipteryx minuta (aves: Enantiornithes) from the Cretaceous of the Gobi Desert. Am. Mus. Novit. 3346, 1–15 (2001).
Close, R. A. et al. Earliest Gondwanan bird from the Cretaceous of southeastern Australia. J. Vertebr. Paleontol. 29, 616–619 (2009).
Naish, D., Martill, D. & Merrick, I. Birds of the Crato Formation in The Crato Fossil Beds of Brazil: Window into an Ancient World (eds. Martill, D. M., Bechly, G. & Loveridge R. F. ) 525–533 (Cambridge University Press, 2007).
Clarke, J. A. Morphology, phylogenetic taxonomy, and systematics of Ichthyornis and Apatornis (Avialae: Ornithurae). B. Am. Mus. Nat. Hist. 286, 1–179 (2004).
Martin, L. D. A new hesperornithid and the relationships of the Mesozoic birds. T. Kansas Acad. Sci. 87, 141–150 (1984).
Martin, J. E. & Cordes-Pearson, A. A new species of the diving bird Baptornis (Ornithurae: Hesperornithoformes) from the lower Pierre Shale Group (upper Cretaceous) of southwestern South Dakota. Geol. S. Am. S. 427, 227–237 (2007).
Martin, L. & Lim, J. New information on the hesperornithiform radition in Proceedings of the 5th Symposium of the Society of Avian Paleontology & Evolution (eds. Zhou, Z. & Zhang, F. ) 165–174 (Science Press, Beijing, 2002).
Martin, L. & Tate Jr., J. A. Hesperornis from the Pierre Shale. Nebraska Acad. Sci. Proc. 77, 40 (1967).
Martin, J. E. & Varner, D. W. The occurence of Hesperornis in the Late Cretaceous Niobrara Formation of South Dakota. Proc. South Dakota Acad. Sci. 71, 95–97 (1992).
Tokaryk, T. T. A toothed bird Hesperornis sp. (Hesperornithiformes) from the Pierre Shale (Late Cretaceous) of Saskatchewan. Can. Field Nat. 113, 670–672 (1999).
Sanchez, J. Late Cretaceous (Cenomanian) Hesperornithiformes from the Pasquia Hills, Saskatchewan, Canada. Dissertation. Carleton University, Ottawa (2010).
Tokaryk, T. T., Cumbaa, S. L. & Storer, J. E. Early Late Cretaceous birds from Saskatchewan, Canada: the oldest diverse avifauna known from North America. J. Vertebr. Paleontol. 17, 172–176 (1997).
Bell, A. & Everhart, M. J. Remains of small ornithurine birds from a Late Cretaceous (Cenomanian) microsite in Russell County, north-central Kansas. Trans. Kansas Acad. Sci. 114, 115–123 (2011).
Everhart, M. J. & Bell, A. A hesperornithiform limb bone from the basal Greenhorn Formation (Late Cretaceous; Middle Cenomanian) of north central Kansas. J. Vertebr. Paleontol. 29, 952–956 (2009).
Tykoski, R. S. & Fiorillo, A. R. An enantiornithine bird from the lower Middle Cenomanian of Texas. J. Vertebr. Paleontol. 30, 288–292 (2010).
Fox, R. C. Ichthyornis (Aves) from the early Turonian (Late Cretaceous) of Alberta. Can. J. Earth Sci. 21, 258–260 (1984).
Hills, L. V., Nicholls, E. L., Núñez-Betelu, L. K. M. & McIntyre, D. J. Hesperornis (Aves) from Ellesmere Island and palynological correlation of known Canadian localities. Can. J. Earth Sci. 36, 1583–1588 (1999).
Chin, K. et al. Life in a temperate polar sea: a unique taphonomic window on the structure of a Late Cretaceous Arctic marine ecosystem. P. Roy. Soc. Lond. B. Bio. 275, 2675–2685 (2008).
Gauthier, J. Saurischian monophyly and the origin of birds. Mem. Calif. Acad. Sci. 8, 1–55 (1986).
Haeckel, E. H. Allgemeine Entwickelungsgeschichte der Organismen: kritische Grundzüge der mechanischen Wissenschaft von den entstehenden Formen der Organismen, begründet durch die Descendenz Theorie. (Reimer, Berlin, 1866).
Tarduno, J. A., Cottrell, R. D. & Smirnov, A. V. The Cretaceous superchron geodynamo: observations near the tangent cylinder. P. Nat. Acad. Sci. 99, 14020–14025 (2002).
Clarke, J. A. & Norell, M. A. New avialan remains and a review of the known avifauna from the Late Cretaceous Nemegt Formation of Mongolia. Am. Mus. Novit. 3447, 1–12 (2004).
Haas, K. et al. Influence of fish on habitat choice of water birds: a whole system experiment. Ecology 88, 2915–2925 (2007).
Kloskowski, J., Nieoczym, M., Polak, M. & Pitucha, P. Habitat selection by breeding waterbirds at ponds with size-structured fish populations. Naturwissenschaften 97, 673–682 (2010).
Grande, L. & Bemis, W. E. A comprehensive phylogenetic study of amiid fishes (Amiidae) based on comparative skeletal anatomy. An empirical search for interconnected patterns of natural history. J. Vertebr. Paleontol. 18, 1–696 (1998).
Longrich, N. An ornithurine-dominated avifauna from the Belly River Group (Campanian, Upper Cretaceous) of Alberta, Canada. Cretaceous Res. 30, 161–177 (2009).
Clarke, J. A., Tambussi, C. P., Noriega, J. I., Erickson, G. M. & Ketcham, R. A. Definitive fossil evidence for the extant avian radiation in the Cretaceous. Nature 433, 305–308 (2005).
Tambussi, C. & Acosta Hospitaleche, C. Antarctic birds (Neornithes) during the Cretaceous-Eocene times. Rev. Assoc. Geol. Argent. 62, 604–617 (2007).
Chiappe, L. M. & Calvo, J. O. Neuquenornis volans, a new Late Cretaceous bird (Enantiornithes, Avisauridae) from Patagonia, Argentina. Journal of Vertebrate Paleontology 14, 239 (1994).
Lawver, D. R., Debee, A. M., Clarke, J. A. & Rougier, G. W. A new enantiornithine bird from the Upper Cretaceous La Colonia Formation of Patagonia, Argentina. Ann. Carnegie Mus. 80, 35–42 (2011).
Clarke, J. A. & Chiappe, L. M. A new carinate bird from the Late Cretaceous of Patagonia (Argentina). Am. Mus. Novit. 3323, 1–24 (2001).
Chinsamy, A., Martin, L. D. & Dobson, P. Bone microstructure of the diving Hesperornis and the voltant Ichthyornis from the Niobrara Chalk of western Kansas. Cretaceous Res. 19, 225–233 (1998).
Clarke, J. A. & Norell, M. A. The morphology and phylogenetic position of Apsaravis ukhaana from the Late Cretaceous of Mongolia. Am. Mus. Novit. 3387, 1–46 (2002).
You, H. L. et al. A nearly modern amphibious bird from the Early Cretaceous of northwestern China. Science 312, 1640–1643 (2006).
Martin, J. E. & Crame, J. A. Palaeobiological significance of high-latitude Late Cretaceous vertebrate fossils from the James Ross Basin, Antarctica. Geological Society, London, Special Publications 258, 109–124 (2006).
Hope, S. The Mesozoic record of Neornithes (modern birds), in Above the Heads of the Dinosaurs (eds. Chiappe, L. M. & L. Witmer ) 339–388 (University of California Press, 2002).
Longrich, N. R., Tokaryk, T. & Field, D. J. Mass extinction of birds at the Cretaceous-Paleogene (K-Pg) boundary. Proceedings of the National Academy of Sciences 108, 15253–15257 (2011).
Wessel, P., Smith, W. H. F., Scharroo, R., Luis, J. & Wobbe, F. Generic Mapping Tools: Improved Version Released, EOS Trans. AGU 94(45), 409–410 (2013).
Seton, M. et al. Global continental and ocean basin reconstructions since 200 Ma. Earth-Sci. Rev. 113, 212–270 (2012).
Acknowledgements
We thank G. Kloc for fossil preparation. We are also grateful to N. Hebdon and M. Friedman for the estimate and relevance of fish size at the Turonian Axel Heiberg site, and to N. Hebdon for discussions on environmental interpretations. Special thanks to the Canadian Polar Shelf Project for their superb logistic support. This work was supported by the National Science Foundation.
Author information
Authors and Affiliations
Contributions
R.K.B. took part in field studies, analyses of fossils and latitudinal analysis. D.B. provided initial identifications and environmental interpretation. J.C. provided the fossil description, identification, comparative analysis and related interpretation of ornithurines including high latitude forms. J.A.T. led the overall study, including field identification, collection, subsequent analyses and interpretation. All authors contributed to the writing of the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/4.0/ http://creativecommons.org/licenses/by-nc-sa/4.0/
About this article

Cite this article
Bono, R., Clarke, J., Tarduno, J. et al. A Large Ornithurine Bird (Tingmiatornis arctica) from the Turonian High Arctic: Climatic and Evolutionary Implications. Sci Rep 6, 38876 (2016). https://doi.org/10.1038/srep38876
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep38876
This article is cited by
-
A juvenile bird with possible crown-group affinities from a dinosaur-rich Cretaceous ecosystem in North America
BMC Ecology and Evolution (2024)
-
Vegaviidae, a new clade of southern diving birds that survived the K/T boundary
The Science of Nature (2017)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
