
Abstract
Diffusion tensor imaging (DTI) provides us an insight into the micro-architecture of white-matter tracts in the brain. This method has proved promising in understanding and investigating the neuronal tracts and structural connectivity between the brain regions in primates as well as rodents. The close evolutionary relationship between canines and humans may have spawned a unique bond in regard to social cognition rendering them useful as an animal model in translational research. In this study, we acquired diffusion data from anaesthetized dogs and created a DTI-based atlas for a canine model which could be used to investigate various white matter diseases. We illustrate the application of this atlas by calculating DTI tractography based structural connectivity between the anterior cingulate cortex (ACC) and posterior cingulate cortex (PCC) regions of the default mode network (DMN) in dogs. White matter connectivity was investigated to provide structural basis for the functional dissociation observed between the anterior and posterior parts of DMN. A comparison of the integrity of long range structural connections (such as in the DMN) between dogs and humans is likely to provide us with new perspectives on the neural basis of the evolution of cognitive functions.
Similar content being viewed by others

Introduction
Diffusion tensor imaging (DTI) is a magnetic resonance imaging technique that measures the anisotropic diffusion of water molecules in tissues providing useful structural information about white matter and the orientation of neural tracts. DTI first came into existence in the mid-1980s1,2,3 and has been rapidly developing ever since. This technique is noninvasive and is now widely used for studying structural connectivity in healthy populations as well as in psychiatric and neurological diseases traditionally characterized by white matter deficits. Researchers have been using DTI to study human and monkey brains and more recently, to study rodent brains such as mice4. However, there have been only a handful of studies applying this powerful method to canines, despite the increased use of these animals in research5,6.
Dogs are an appealing species to study for biological, behavioral, and evolutionary reasons which have led to the explosion of research focused on dogs (for reviews see7,8). One of the intriguing reasons for this development is that the close evolutionary history between dogs and humans may have spawned a unique bond in regard to social cognition9. In this context, canine social cognition involves studying dog-human interactions to understand how selection pressures via domestication may have (or have not) shaped canine social cognitive abilities (for reviews see Kaminski et al.10 and Miklosi11). These abilities can be tested/observed in a variety of tasks that measure canine performance in the presence of human cues. Such tasks include manipulation of human speech (emotional and semantic content), presence (absence or not), and communicative gestures (gazing, pointing, body posture) that are passively processed or convey solutions to problems (e.g., emotion contagion, emotion discrimination, guesser-knower paradigm, object choice task, object search, responding to commands, strange situation test, unsolvable task12,13,14,15,16). For example, in the object choice task, dogs need to attend to a person’s point and/or gaze to successfully locate a hidden object. Of note, not only do dogs display joint attention with humans and respond to human pointing and gesturing but they do so better than nonhuman primates making them an excellent animal model for human social cognition9,17,18,19. From an evolutionary perspective, the canine advantage is either due to the domestication of dogs (from their predecessors, wolves), their close contact with humans during their daily lives, or is a residual mechanism present before the lineages split 97 million years ago. Unraveling the different phylogenetic and ontogenetic possibilities and their interaction can be solved via triangulating findings comparing the behavioral, anatomical, and functional neural mechanisms within and across species to understand their evolutionary origin. Canine DTI represents an important technique to explore the neural basis of the evolution of cognitive function and its anatomical correspondences. Additionally, establishing a DTI-atlas in canines will serve as an important measure to allow researchers to ask questions concerning how fiber tracts may change with experience20,21.
Studying the canine model might thus give us a perspective into the evolution of cognitive functions. Additionally, dogs serve as a large animal model, critical for translation of quantitative MRI methods to clinical populations22, share a common environment with their owners until their old age, and receive a good level of healthcare23. Furthermore, rabies has a long history of association with dogs and is almost never known to infect or to be transmitted to humans by small rodents such as rats, chipmunks or rabbits24. For these reasons dogs are an interesting species worth studying as animal models in translational research to study aging and its effect on disease progression as well as to investigate human diseases that affect white matter integrity such as neurological demyelinating conditions (e.g., Multiple Sclerosis)22, GM1 gangliosidosis, a fatal neurodegenerative lysosomal storage disease25 and Rabies, a virtually incurable disease26. Thus, non-invasive imaging of neural white matter tracts in the dog brain by using DTI could prove to be very useful.
There have been a few DTI studies conducted on canine models including an ex-vivo study that was conducted on euthanized canine brains to study the white matter fibers by analyzing the fractional anisotropy (FA) and directional maps computed using the DTI images acquired within first 2 hours after death5 and an in vivo study investigating feasibility of diffusion tensor tractography in a canine model for studying the cerebral white matter in anesthetized dogs6. However, to our knowledge, ours is the first study to date, to have investigated in vivo diffusion images of dog brains to provide us with group-wise FA color maps. Therefore, in this study, we aimed to build a canine DTI-atlas, and demonstrate the utility of the atlas by testing a hypothesis derived from previous work in our lab27.
With regard to the second aim of our study based upon a resting state fMRI study recently conducted on dog brains in our lab27, we aspired to test the hypothesis that the functional connectivity dissociation between the anterior and posterior parts of the default mode network (DMN) in the canine model possesses a structural basis.
To accomplish the aims of our study, we first acquired diffusion-weighted images (DWI) in anesthetized dogs to develop a DTI-based atlas of the dog brain for use in future studies by giving an insight into the structural connectivity inside the dog brain. Second, we illustrate the utility of the dog DTI atlas to test a specific hypothesis arising from our previous resting state functional magnetic resonance imaging (fMRI) study27. Specifically, in our previous study we observed that the anterior and posterior parts of the DMN seem to be dissociated, unlike in monkey and human brains28,29. Therefore, we hypothesized that DTI tractography, specifically in the dorsal cingulum connecting the posterior cingulate cortex (PCC) and anterior cingulate cortex/medial pre-frontal cortex (ACC/MPFC), could provide a structural basis for this dissociation in the form of weaker structural connection between these regions in dogs as compared to humans.
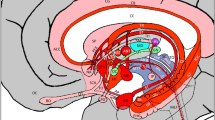
The DMN is an interconnected network of brain regions that is active when an individual is at rest and becomes less active when performing attentionally demanding tasks30,31. The network preferentially activates when an individual is daydreaming, engaged in self-retrospection and self-introspection32. The core brain regions of DMN, consist of the medial prefrontal cortex (MPFC), posterior cingulate cortex (PCC), inferior parietal cortex (IPC), inferior temporal cortex (ITC) and (para) hippocampal formation. The main hubs of the DMN are within the MPFC and along the posterior midline including PCC32. In the DMN, PCC has been proposed to be of special interest as it is the only region that directly interacts with all other DMN nodes33. The anterior regions (ACC/MPFC) of the DMN along with the medial temporal lobe facilitates flexible use of episodic memories and associations to construct self-relevant mental simulations. These regions have has also been implicated in judgment of others34. The above mentioned two subsystems converge on important nodes of integration including PCC. PCC is commonly associated with episodic memory retrieval35,36. Therefore, DMN is believed to be responsible for consciousness and self-referential processing and has been linked to human cognition32,37,38. Studies have demonstrated the existence of the DMN in humans and monkeys28,29, however there are conflicting reports on whether they exist in rodents39,40,41. Our resting state fMRI study conducted on dog brains indicated that the anterior and posterior parts of the DMN are dissociated in dogs27. Previously conducted research has illustrated that the DMN exhibits the highest overlap in its structural and functional connectivity42. Therefore, we believed that there might be a significant possibility of the existence of a structural basis for the observed dissociation in the DMN in dogs. We thus decided to investigate it in the anticipation of attaining an insight into the evolution of cognitive functions.
Methods
All methods and experiments were approved by the Auburn University Institutional Animal Care and Use Committee. We confirm that all methods were performed in accordance with the relevant guidelines and regulations. DWI data were acquired using a 3T MAGNETOM Verio (Siemens Healthcare, Erlangen, Germany) MRI scanner from 23 anaesthetized Labrador retriever dogs (age range 12–60 months) using a human knee coil (serving as a dog head coil) and an EPI (Echo Planar Imaging) based diffusion sequence with the following parameters: TR = 3.6 s, TE = 95 ms, flip angle = 90 °, 128 × 128 acquisition matrix, 30 diffusion directions, b = 0 and 1000, voxel size = 3 × 3 × 3 mm3. Dogs were sedated and lightly anesthetized with intramuscularly administered xylazine (2.2 mg/kg) and ketamine HCL (11 mg/kg), respectively. Details regarding the imaging procedure can be found in our previous publication27 describing resting state fMRI analysis on a subset of the dog sample employed here. Data were preprocessed using the standard FMRIB’s Diffusion Toolbox (FDT, which is part of the FMRIB software library (FSL)43,44,45. Data were first brain extracted, and then corrected for eddy current induced distortions.
DTI-based Atlas
Following eddy current correction46, diffusion tensors were fit to the corrected data, creating tensor and fractional anisotropy (FA) maps for each dog43. Individual FA maps were then registered to a high-resolution canine ex vivo template created in a previous study47 using FMRIB’s linear registration tool (FLIRT)48,49,50. The resulting transformation matrix, computed while registering FA maps to the template, was applied to the tensor maps for each dog, thus obtaining the vector data of each subject in the same standard space using the “vecreg” command line tool in FSL as described in ref. 51. By averaging FA maps as well as vector data in the template space for all the dogs, an average FA map and average color-coded tensor map were computed to reduce the inter-subject variability in canine brain.
Diffusion Tractography
Diffusion parameters were modeled using the BEDPOSTX tool in FSL for the eddy corrected data, and then probabilistic tractography was implemented as described previously52,53,54 using the PROBTRACKX tool in FSL to estimate the likely connections between two regions of interest (ROIs). The following parameters were used for BEDPOSTX: number of fibers per voxel = 2, burnin period = 1000, number of jumps = 1250, ‘sample every’ = 25, model with sticks, gradient nonlinearities considered and ARD weight = 1. For PROBTRACKX, we used the following parameters: number of samples = 5000, curvature threshold = 0.2, step length = 0.5, and maximum number of steps = 2000. The ROIs were defined as 4 mm radius spheres in the PCC and ACC/MPFC. The ROIs were positioned based on the co-ordinates of the peak z-values in PCC and ACC/MPFC obtained from the group independent component corresponding to the DMN which was derived from an ICA analysis of resting state fMRI data in the current sample. Probabilistic tracking was achieved by creating a probability density function at each voxel on the principal diffusion directions. Tractography analysis was performed at the whole brain level and not confined to individual hemispheres. Connectivity probabilities were estimated between the ROIs by repetitive sampling of the streamlines through the probability distribution function. The ROIs were defined in the template/atlas space and then registered to each subject’s native space using the inverse of the previously computed transformation matrix. In this way, fiber tracts were calculated in the subject space, and then transformed to the template/atlas space by applying the appropriate transformation computed earlier. The ACC-PCC tracts of each subject were then binarized and compiled to create a group level probability map of the tracts for dogs with a subject count threshold in which the value of each voxel represents the number of subjects that had a tract passing though that voxel. The group level probability map of the ACC-PCC tracts was binarized and overlaid on the group-wise mean FA map computed earlier. A group-wise mean FA value was extracted from the ACC-PCC tracts by averaging the FA values for the voxels constituting the ACC-PCC group-level connectivity map.
Results
Average FA and tensor-based maps were computed for individual dogs using the diffusion tensor computed from the acquired DWI data (Fig. 1A), and a group-wise average FA map with associated color map representing the first principal vector (i.e., the main diffusion direction) are shown in Fig. 1B. This provides a robust atlas void of disproportionate influence from any one subject. Fractional anisotropy values in the group FA map ranged from 0 to 1, with values closer to 1 in regions with higher anisotropy and more directional diffusion of water such as white matter, and closer to 0 in the areas of the brain with isotropic diffusion such as cerebrospinal fluid (CSF). Group mean FA was extracted from the ACC-PCC tract which was found to be 0.32 which is much lower compared to the mean FA previously calculated for humans (young adults)55, which was 0.57 (the caveats associated with this comparison are discussed in the next section). Additionally, the group level probability map of the tracts is shown in Fig. 2. The map shows the number of dogs having a tract passing through each voxel. Tracts connecting ACC to PCC were detected in only 9 out of 23 dogs. Even though, the tracts were detected in 9 of 23 dogs, the probability maps show voxel values less than 9 due to the inter-subject variability in the precise fiber pathways from one voxel to the next.

DTI Atlas.
(A) FA map and Tensor based color map for one individual dog. (B) Average FA map and color map atlas using a group of 23 dogs. Color map represents left-right tracts in Red, anterior-posterior tracts in Green and superior-inferior tracts in Blue.

Group level probabilistic map of ACC-PCC tracts.
The color scale indicates the percentage of dogs that had a tract in a given voxel. The green ROI corresponds to the ACC/MPFC while the yellow one shows the PCC.
Discussion
The average color map gives a qualitative characterization of the neural tracts in the white matter of a canine brain. The group-wise average FA map and tensor based map provides us with a DTI-based atlas for dogs by accounting for the inter-subject variability. This atlas can be utilized in future studies in both veterinary medicine and translational research. Readers can download the dog DTI atlas using the following link for such investigations: http://aucanlab.com/wp-content/uploads/2015/05/DogDTIAtlas.zip
As previously discussed, studying canines as an animal model in translational research could prove promising, keeping in mind that canines are phylogenetically distant from humans. Given their co-evolution with humans1,9,17 and capabilities to respond to human cues17,18,19,56, they are an interesting species worth studying to investigate how similar traits may have arisen via evolution. One brain network that has been implicated in cognition in general (as well as in social cognition)57,58 is the default mode network. The DMN is also known to underlie self-referential processing, consciousness, self-awareness and higher cognition in humans32,37,38.
A previous resting state fMRI study investigating the existence of the DMN in dogs, demonstrated disconnected ACC and PCC, the anterior and posterior regions of the DMN27. There is evidence that suggests that the DMN has a strong correlation between its structural and functional connectivity42. Therefore, in this study we tested the hypothesis that the functional finding about the anterior-posterior dissociation of the DMN in canines possesses a structural basis. The group mean FA for the MPFC/ACC-PCC tracts identified in the dog brain was notably low and also comparably lower than that observed in humans55. Additionally, the MPFC/ACC-PCC tract was found only in 9 out of 23 dogs indicating significant individual variability. As previously reported in a study conducted with humans59, tracts connecting MPFC/ACC to PCC were detected in 22 of 23 subjects. These observations lend support for our hypothesis that the weaker resting state functional connectivity found between the ACC and PCC in dogs compared to humans may have a structural basis.
Our results point toward certain hypotheses which can be tested in future studies. Given dogs perform better than nonhuman primates in several tests of social cognition, it raises questions about the centrality of the strong connection between the ACC and PCC found in humans and nonhuman primates for social cognition. In order to test related hypotheses, performance in behavioral tasks could be correlated with DTI-based metrics such as FA values. An additional advantage with DTI studies in animal research is that tract reconstruction can be compared with tracer studies to analyze corticocortical connectivity60. Such analyses could potentially give insight into individual canine performance as well.
While the current findings lend some support to convergent evolution (as opposed to homology due to the weak ACC and PCC connection in canines), it may well be that dogs are well equipped to learn the appropriate cues ontologically to perform well in social cognition tasks. Likely, it is an interaction between phylogenetic and ontogenetic factors that underlie the social cognition traits in dogs. The DTI-atlas can serve to allow researchers to ask questions concerning how fiber tracts may change with experience20,21. If such canine brain plasticity occurs, it could reveal an important piece of the puzzle as to how dogs become expert detector dogs, therapy dogs, and succeed in social cognition and other cognitive tasks. These aspects are worthy of future investigations.
Limitations
While creating a DTI atlas for the dog brain is a clear contribution of the paper, direct comparison of FA values between dogs and humans for testing our specific structural hypothesis regarding the DMN involves certain caveats: (i) the size of the brain in dogs is smaller than in humans, but the spatial resolution of diffusion data acquired was not proportionally higher in dogs, (ii) the exact placement of seed ROIs in humans is likely to be more accurate than in dogs given that labeled atlases are available in humans, and (iii) the anatomy of dog and human brains, especially the frontal cortex, may not be exactly comparable. Further, the atlas itself is limited to canines of a specific breed (Labradors in this case) and age range. Larger studies in the future employing a larger sample of canines drawn from various breeds and age ranges are required to develop a more general canine DTI atlas. The current investigation is also constrained by the general limitations of diffusion tensor imaging, including spatial resolution, which lends itself to multiple fiber orientations being present in a single voxel (and thus, their contributions to values such as anisotropy may be averaged). Given that high diffusion anisotropy values depend on microscopic sources of anisotropy and macroscopic homogeneity within a voxel, some regions may be misrepresented as having no fibers (FA < 0.2), when really they just have multiple orientations. Relatedly, there is a vast data reduction that occurs in the analysis as algorithms typically assign 1 or 2 directions within a given voxel. Finally, DTI is sensitive to the motion of water which is relatively minor (~5–10 um), lending itself vulnerable to physiological artifacts that could interfere with DTI. Notwithstanding these limitations, our preliminary results regarding the anterior-posterior structural (as well as functional) dissociation in the canine DMN is worth exploration in the future for understanding the evolutionary significance of the DMN as well as the evolutionary bond between dogs and humans.
Additional Information
How to cite this article: Robinson, J. L. et al. Characterization of Structural Connectivity of the Default Mode Network in Dogs using Diffusion Tensor Imaging. Sci. Rep. 6, 36851; doi: 10.1038/srep36851 (2016).
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Le Bihan, D. & Breton, E. Imagerie de diffusion in-vivo par résonance. C R Acad Sci (Paris) 301(15), 1109–1112, doi: 10.1002/nbm.3530 (1985).
Merboldt, K. D., Hanicke, W. & Frahm, J. Diffusion imaging using stimulated echoes. Magn. Reson. Med. 19, 233–239, doi: 10.1002/mrm.1910190208 (1991).
Taylor, D. G. & Bushell, M. The spatial mapping of translational diffusion coefficients by the NMR imaging technique. Phys. Med. Biol. 30(4), 345–349, doi: 10.1088/0031-9155/30/4/009 (1985).
Baloch, S. et al. Quantification of Brain Maturation and Growth Patterns in C57BL/6J Mice via Computational Neuroanatomy of Diffusion Tensor Images. Cerebral Cortex (New York, NY) 19(3), 675–687, doi: 10.1093/cercor/bhn112 (2009).
Jacqmot, O. et al. Diffusion Tensor Imaging of White Matter Tracts in the Dog Brain. Anat Rec 296, 340–349, doi: 10.1002/ar.22638 (2013).
Anaya García, M. S., Hernández Anaya, J. S., Marrufo Meléndez, O., Velázquez Ramírez, J. L. & Palacios Aguiar, R. In vivo study OF cerebral white matter in the dog using diffusion tensor tractography. Veterinary Radiology & Ultrasound 56(2), 188–195, doi: 10.1111/vru.12211 (2015).
Bensky, M. K., Gosling, S. D. & Sinn, D. L. The World from a Dog’s Point of View: A Review and Synthesis of Dog Cognition Research. Advances in the Study of Behavior 45, 209–406, doi: 10.1016/B978-0-12-407186-5.00005-7 (2013).
Feuerbacher, E. N. & Wynne, C. D. L. A history of dogs as subjects in North American experimental psychological research. Comparative Cognition and Behavior Reviews 6, 46–71, doi: 10.3819/ccbr.2011.60001 (2011).
Hare, B. & Tomasello, M. Human-like social skills in dogs? Trends Cogn Sci. 9(9), 439–444, doi: 10.1016/j.tics.2005.07.003 (2005).
Kaminski, J. & Marshall-Pescini, S. The Social Dog: Behavior and Cognition (Elsevier Science, Burlington, VT, 2014).
Miklósi, Á. Dog Behavior, Evolution, and Cognition (Oxford University Press, New York, NY, 2007).
Andics, A., Gácsi, M., Faragó, T., Kis, A. & Miklósi, Á. Voice-sensitive regions in the dog and human brain are revealed by comparative fMRI. Current Biology 24(5), 574–578, doi: 10.1016/j.cub.2014.01.058 (2014).
Andics, A. et al. Neural mechanisms for lexical processing in dogs. Science 353, 1030–1032, doi: 10.1126/science.aaf3777 (2016).
Kaminski, J., Call, J. & Fischer, M. Word learning in a domestic dog: Evidence for fast mapping. Science 304, 1682–1683, doi: 10.1126/science.1097859 (2004).
Pilley, J. & Reid, A. Border collie comprehends object names as verbal referents. Behavioral Processes 86, 184–195, doi: 10.1016/j.beproc.2010.11.007 (2011).
Teglas, E., Gergely, A., Kupan, K., Miklosi, A. & Topal, J. Dog’s gaze following is tuned to human communicative signals. Current Biology 22, 209–222, doi: 10.1016/j.cub.2011.12.018 (2012).
Reid, P. Adapting to the human world: dogs’ responsiveness to our social cues. Behav Processes, 80(3), 325–333, doi: 10.1016/j.beproc.2008.11.002 (2009).
Soproni, K., Miklósi, Á., Topál, J. & Csányi, V. Dogs’ (Canis familaris) responsiveness to human pointing gestures. Journal of Comparative Psychology 116(1), 27–34, doi: 10.1037/0735-7036.116.1.27 (2002).
Lazarowski, L. & Dorman, D. C. A comparison of pet and purpose-bred research dog (Canis familiaris) performance on human-guided object-choice tasks. Behav Processes 110, 60–67, doi: 10.1016/j.beproc.2014.09.021 (2015).
Voelker, P. et al. How changes in white matter might underlie improved reaction time due to practice. Cognitive Neuroscience, in press, doi: 10.1080/17588928.2006.1173664 (2016).
Wang, S. & Young, K. White matter plasticity in adulthood. Neuroscience 276, 148–160, doi: 10.1016/j.neuroscience.2013.10.018 (2014).
Wu, Y. C. et al. High b-value and diffusion tensor imaging in a canine model of dysmyelination and brain maturation. Neuroimage 58(3), 829–837, doi: 10.1016/j.neuroimage.2011.06.067 (2011).
Kingfisher. Biotech Inc., Kingfisher Biotech Circular Subject: Canine as an Animal Model Volume 1; No. 1, Available at http://www.kingfisherbiotech.com/newsletter/Canine_Animal_Model_Newsletter.pdf. Date accessed: (6th October 2016).
Centers for Disease Control and Prevention., Rabies. Other Wild Animals: Terrestrial carnivores: raccoons, skunks and foxes., Available at http://www.cdc.gov/rabies/exposure/animals/other.html Date accessed: 6th October 2016 (2011).
Hasegawa, D. et al. Serial MRI Features of Canine GM1 Gangliosidosis: A Possible Imaging Biomarker for Diagnosis and Progression of the Disease. The Scientific World Journal 250197, doi: 10.1100/2012/250197 (2012).
Laothamatas, J., Sungkarat, W. & Hemachudha, T. Neuroimaging in rabies. Advances in Virus Research Volume 79, 309–327, doi: 10.1016/B978-0-12-387040-7.00014-7 (2011).
Kyathanahally, S. P. et al. Anterior–posterior dissociation of the default mode network in dogs. Brain Structure and Function 220(2), 1063–1076, doi: 10.1007/s00429-013-0700-x (2015).
Mantini, D. et al. Default mode of brain function in monkeys. J Neurosci. 31(36), 12954–12962, doi: 10.1523/JNEUROSCI.2318-11.2011 (2011).
Greicius, M. D., Krasnow, B., Reiss, A. L. & Menon, V. Functional connectivity in the resting brain: A network analysis of the default mode hypothesis. Proceedings of the National Academy of Sciences of the United States of America 100(1), 253–258, doi: 10.1073/pnas.0135058100 (2003).
Fox, M. D. & Raichle, M. Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nat Rev Neurosci. 8, 700–711, doi: 10.1038/nrn2201 (2007).
Raichle, M. E. et al. A default mode of brain function. Proc Natl Acad Sci USA 98, 676–682, doi: 10.1073/pnas.98.2.676 (2001).
Buckner, R. L., Andrews-Hanna, J. R. & Schacter, D. L. The brain’s default network: anatomy, function, and relevance to disease. Ann N Y Acad Sci. 1124, 1–38, doi: 10.1196/annals.1440.011 (2008).
Fransson, P. & Marrelec, G. The precuneus/posterior cingulate cortex plays a pivotal role in the default mode network: Evidence from a partial correlation network analysis. Neuroimage 42, 1178–1184, doi: 10.1016/j.neuroimage.2008.05.059 (2008).
Mitchell, J. P., Heatherton, T. F. & Macrae, C. N. Distinct neural systems subserve person and object knowledge. Proc Natl Acad Sci U S A. 99(23), 15238–15243, doi: 10.1073/pnas.232395699 (2002).
Lou, H. C. et al. Parietal cortex and representation of the mental Self. Proc Natl Acad Sci USA 101(17), 6827–6832, doi: 10.1073/pnas.0400049101 (2004).
Lundstrom, B. N., Ingvar, M. & Petersson, K. M. The role of precuneus and left inferior frontal cortex during source memory episodic retrieval. Neuroimage 27(4), 824–834, doi: 10.1016/j.neuroimage.2005.05.008 (2005).
Greicius, M. Resting-state functional connectivity in neuropsychiatric disorders. Curr Opin Neurol 21, 424–430, doi: 10.1097/WCO.0b013e328306f2c5 (2008).
Gusnard, D. A. & Raichle, M. Searching for a baseline: functional imaging and the resting human brain. Nat Rev Neurosci. 2, 685–694, doi: 10.1038/35094500 (2001).
Becerra, L., Pendse, G., Chang, P.-C., Bishop, J. & Borsook, D. Robust reproducible resting state networks in the awake rodent brain. PloS One 6(10), e25701, doi: 10.1371/journal.pone.0025701 (2011).
Lu, H. et al. Rat brains also have a default mode network. Proc Natl Acad Sci USA 109(10), 3979–3984, doi: 10.1073/pnas.1200506109 (2011).
Upadhyay, J. et al. Default-mode-like network activation in awake rodents. PloS One 6(11), e27839, doi: 10.1371/journal.pone.0027839 (2011).
Horn, A., Ostwald, D., Reisert, M. & Blankenburg, F. The structural-functional connectome and the default mode network of the human brain. NeuroImage 13, 1053–8119, doi: 10.1016/j.neuroimage.2013.09.069 (2013).
Smith, S. M. et al. Advances in functional and structural MR image analysis and implementation as FSL. NeuroImage 23(S1), 208–219, doi: 10.1016/j.neuroimage.2004.07.051 (2004).
Woolrich, M. W. et al. Bayesian analysis of neuroimaging data in FSL. NeuroImage 45, S173–S186, doi: 10.1016/j.neuroimage.2008.10.055 (2009).
Jenkinson, M., Beckmann, C. F., Behrens, T. E., Woolrich, M. W. & Smith, S. M. FSL. NeuroImage 62, 782–790, doi: 10.1016/j.neuroimage.2011.09.015 (2012).
Jenkinson, M., Pechaud, M. & Smith, S. BET2: MR-based estimation of brain, skull and scalp surfaces. In Eleventh Annual Meeting of the Organization for Human Brain Mapping Québec City, Canada (2005).
Datta, R. et al. A digital atlas of the dog brain. PLoS One 7(12), e52140, doi: 10.1371/journal.pone.0052140 (2012).
Jenkinson, M., Bannister, P., Brady, J. M. & Smith, S. M. Improved Optimisation for the Robust and Accurate Linear Registration and Motion Correction of Brain Images. NeuroImage 17(2), 825–841, doi: 10.1006/nimg.2002.1132 (2002).
Jenkinson, M. & Smith, S. A Global Optimisation Method for Robust Affine Registration of Brain Images. Medical Image Analysis 5(2), 143–156, doi: 10.1006/nimg.2002.1132 (2002).
Greve, D. N. & Fischl, B. Accurate and robust brain image alignment using boundary-based registration. NeuroImage 48(1), 63–72, doi: 10.1016/j.neuroimage.2009.06.060 (2009).
Alexander, D. C., Basser, P. J., Pierpaoli, C. & Gee, J. C. Spatial transformations of diffusion tensor Images. IEEE Trans. Med. Imag. 20(11), 1131–1139, doi: 10.1109/42.963816 (2001).
Behrens, T. E. J. et al. Non-invasive mapping of connections between human thalamus and cortex using diffusion imaging. Nature Neuroscience 6(7), 750–757, doi: 10.1038/nn1075 (2003).
Behrens, T. E. J. et al. Characterization and propagation of uncertainty in diffusion-weighted MR imaging. Magn Reson Med. 50(5), 1077–1088, doi: 10.1002/mrm.10609 (2003).
Johansen-Berg, H. et al. Changes in connectivity profiles define functionally distinct regions in human medial frontal cortex. Proc Natl Acad Sci USA 101(36), 13335–13340, doi: 10.1073/pnas.0403743101 (2004).
Supekar, K. et al. Development of functional and structural connectivity within the default mode network in young children. Neuroimage 52(1), 290–301, doi: 10.1016/j.neuroimage.2010.04.009 (2010).
Merola, I., Prato-Previde, E. & Marshall-Pescini, S. Dogs’ social referencing towards owners and strangers. PLoS One 7(10), e47653, doi: 10.1371/journal.pone.0047653 (2012).
Li, W., Mai, X. & Liu, C. The default mode network and social understanding of others: what do brain connectivity studies tell us. Front Hum Neurosci. 8, 74, doi: 10.3389/fnhum.2014.00074 (2014).
Che, X. et al. Synchronous activation within the default mode network correlates with perceived social support. Neuropsychologia 63, 26–33, doi: 10.1016/j.neuropsychologia.2014.07.035 (2014).
Greicius, M. D., Supekar, K., Menon, V. & Dougherty, R. F. Resting-State Functional Connectivity Reflects Structural Connectivity in the Default Mode Network. Cerebral Cortex (New York, NY) 19(1), 72–78, doi: 10.1093/cercor/bhn059 (2009).
Jbabdi, S., Sotiropoulos, S., Haber, S., Van Essen, D. & Behrens, T. Measuring macroscopic brain connections in vivo. Nature Neuroscience 18, 1546–1555, doi: 10.1038/nn.4134 (2015).
Acknowledgements
Funding from Auburn University’s Intramural Grant and from the Defense Advanced Research Projects Agency (government contract/grant number W911QX-13-C-0123) is gratefully acknowledged. The views, opinions, and/or findings contained in this article are those of the authors and should not be interpreted as representing the official views or policies, either expressed or implied, of the Defense Advanced Research Projects Agency, US Department of Defense or the federal Government of the United States.
Author information
Authors and Affiliations
Contributions
G.D., V.V., T.S.D., E.M. and P.W. conceived the idea and designed the experiment. G.D., N.S. and R.B. optimized sequences and acquired the data. P.W. handled the dogs. J.L.R., M.B. and G.D. analyzed the data. J.L.R., M.B., J.K. and G.D. interpreted the results. All authors contributed towards writing the manuscript.
Ethics declarations
Competing interests
Author N.S. is an employee of Siemens Healthcare, Malvern, PA who is stationed at the MRI Research Center in Auburn University. She contributed by optimizing the sequences used to acquire data in this study. However, note that she had no role in data analysis and its interpretation. These competing interests do not alter the authors’ adherence to common policies on sharing data and materials.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Robinson, J., Baxi, M., Katz, J. et al. Characterization of Structural Connectivity of the Default Mode Network in Dogs using Diffusion Tensor Imaging. Sci Rep 6, 36851 (2016). https://doi.org/10.1038/srep36851
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep36851
This article is cited by
-
Central nodes of canine functional brain networks are concentrated in the cingulate gyrus
Brain Structure and Function (2023)
-
Increased resting state connectivity in the anterior default mode network of idiopathic epileptic dogs
Scientific Reports (2021)
-
Human–dog relationships as a working framework for exploring human–robot attachment: a multidisciplinary review
Animal Cognition (2021)
-
Default mode network integrity changes contribute to cognitive deficits in subcortical vascular cognitive impairment, no dementia
Brain Imaging and Behavior (2021)
-
Dog–human social relationship: representation of human face familiarity and emotions in the dog brain
Animal Cognition (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
