
Abstract
Glomeruli are highly sophisticated filters and glomerular disease is the leading cause of kidney failure. Morphological change in glomerular podocytes and the underlying basement membrane are frequently observed in disease, irrespective of the underlying molecular etiology. Standard electron microscopy techniques have enabled the identification and classification of glomerular diseases based on two-dimensional information, however complex three-dimensional ultrastructural relationships between cells and their extracellular matrix cannot be easily resolved with this approach. We employed serial block face-scanning electron microscopy to investigate Alport syndrome, the commonest monogenic glomerular disease, and compared findings to other genetic mouse models of glomerular disease (Myo1e−/−, Ptpro−/−). These analyses revealed the evolution of basement membrane and cellular defects through the progression of glomerular injury. Specifically we identified sub-podocyte expansions of the basement membrane with both cellular and matrix gene defects and found a corresponding reduction in podocyte foot process number. Furthermore, we discovered novel podocyte protrusions invading into the glomerular basement membrane in disease and these occurred frequently in expanded regions of basement membrane. These findings provide new insights into mechanisms of glomerular barrier dysfunction and suggest that common cell-matrix-adhesion pathways are involved in the progression of disease regardless of the primary insult.
Similar content being viewed by others
Introduction
Glomerular disease is characterised by persistent proteinuria, with or without haematuria, and a progressive decline in renal function. Currently, no curative therapy exists and patients are reliant upon non-specific treatments. Glomerular dysfunction can result from genetic defects, inflammatory insults or can be secondary to systemic disease such as diabetes mellitus. Alport syndrome is the commonest monogenic glomerular disease and is characterised by progressive glomerular dysfunction, sensorineural deafness and ocular abnormalities1. Patients with Alport syndrome are treated with renin-angiotensin-aldosterone system (RAAS) inhibitors and this therapy has been shown to prolong renal function by over 15 years2. However, many patients ultimately progress to end stage renal disease and more effective therapies are urgently needed.
The molecular origin of Alport Syndrome was first described in 1990 when mutations in COL4A5 were reported3. COL4A5 is expressed on the X-chromosome and therefore hemizygous males are more severely affected than heterozygous females3,4. Subsequently, mutations in COL4A3 and COL4A4 were discovered and in this scenario two mutated alleles cause autosomal recessive disease5,6,7. Within the kidney, glomerular podocytes are the only cells known to express COL4A3, COL4A4 and COL4A58, and these genes are required for the assembly of the type IV collagen α3α4α5 network in the glomerular basement membrane (GBM). The GBM is a complex niche of extracellular matrix (ECM) proteins that is essential for normal glomerular function9. In the adult glomerulus the GBM is composed of predominately type IV collagen α3α4α5, laminin α5β2γ1 and up to 70 additional structural and regulatory components10. Evidence suggests that type IV collagen α3α4α5 is important for the lifelong structural integrity of the GBM11, as the type IV collagen α1α1α2 network, produced by both podocytes and glomerular endothelial cells12, is unable to compensate for the absence of the α3α4α5 network in Alport Syndrome13,14.
Renal biopsy is a valuable diagnostic tool for defining the sub-type of glomerular disease; indeed a diagnostic marker for Alport Syndrome is the pathological basket weave appearance of the GBM, although genetic testing is now superseding biopsy15. Other histopathological features of glomerular disease include altered GBM thickness, loss of unique podocyte structures called foot processes (FPs) and a reduction in podocyte number16. Interestingly, these features are reported across the spectrum of glomerular disease regardless of the underlying molecular aetiology. Yet the mechanisms underpinning these morphological changes are poorly understood. In particular we have not defined the ultrastructural relationship between podocytes and the GBM during the progression of glomerular disease, however this knowledge could provide insight into disease mechanisms.
To address this knowledge gap we employed serial block face-scanning electron microscopy (SBF-SEM) to examine the relationships of cells and ECM in three-dimensions (3D). This approach was recently used to investigate cell-ECM interactions17,18,19,20 and the complexities of normal podocyte structure21. However, until now, the approach has not been used to investigate changes in podocytes and the GBM during disease progression. We hypothesized that SBF-SEM over a time course would enable the construction of detailed 3D models of the glomerulus in health and disease and would reveal novel pathological features.
Results
3D modelling of glomerular structure in Alport nephropathy
Over a 28-week time course we investigated Col4a3−/− male mice on a C57BL/6J genetic background. We selected this background, which has a slow progression of glomerular disease, to allow a greater window of observation. We analyzed glomeruli using SBF-SEM and created 3D glomerular reconstructions (Supplementary Movie S1). The glomerulus is composed of interwoven capillary loops forming a complex pattern of lumens when viewed in 3D. The localization of the GBM, podocyte cell bodies and foot processes (FPs) can be easily discerned using SBF-SEM (Fig. 1a). From wild type 16–18 week (older adult) mouse glomeruli we generated models of podocytes and GBM and these revealed an organized glomerular structure (Fig. 1a,b). We compared these models with glomeruli from 6-week (young adult) Col4a3+/− and Col4a3−/− animals and found that the majority of podocytes had regular FPs (Fig. 2). This finding was also observed in older adult Col4a3−/− mice although some FPs were flattened or effaced (Fig. 2). In contrast, 28-week (aged) Col4a3−/− mice had striking global loss of podocyte FP organization, GBM thickening and thinning (Fig. 2). Further analysis revealed focal regions of reduced podocyte FP density in all groups of Col4a3−/− mice, even in young adult mice, when compared with age matched wild type and Col4a3+/− mice (Fig. 3a,b). These areas were often concomitant with thickened, non-uniform sheets of GBM (Fig. 3a,c and Supplementary Movies S2 and S3). Moreover, the density of podocyte FPs decreased with age in Col4a3−/− mice (Fig. 3b) and correlated with increased thickness and variation in thickness of the GBM (Fig. 3c).

The 3D morphology of podocyte foot processes and the GBM.
3D reconstructions from SBF-SEM images of wild type glomerulus from an 18-week C57Bl/6 mouse. (a) A single slice from an SBF-SEM stack showing a glomerulus in cross section. Glomerular capillary lumens are highlighted in red, the GBM in blue and podocyte cell bodies in green. Regions of interest (ROIs) show that podocyte FPs from two adjacent podocytes (one green the other magenta) can clearly be discerned using this technique. (b) Panel displays a transverse view of reconstructed podocyte FPs and GBM. Removing slices (increasing Z distance) reveals the 3-dimensional organization of the foot processes. Serial block face-scanning electron microscopy, SBF-SEM; glomerular basement membrane, GBM; foot processes, FPs.

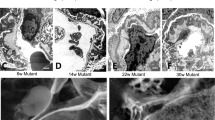
Normal podocyte FP and GBM structure is lost with age in Alport mice.
Transverse views of reconstructed GBMs (blue) and podocyte FPs (green and magenta) from wild type, Col4a3−/− mice and Col4a3−/− mice. Many regions in young adult (young) and older adult (adult) Col4a3−/− mice display organized podocyte FPs and uniform GBM of thickness comparable to wild type adult animals. GBM, glomerular basement membrane; FPs, foot processes.

3D analysis of glomerular structure in aging Col4a3−/− mice.
(a) Transverse view of reconstructed GBMs (blue) and podocyte FPs (green and magenta) reveal focal areas of podocyte FP effacement that coincide with thickened irregular regions of GBM. (b) Quantification of podocyte FP density. The violin plots demonstrate the distributions for the number of FPs per length of GBM along the y-axis, the black lines represent the mean number of FPs per length of GBM in each group. (C) Quantification of GBM thickness. The violin plots describe the distributions for the thickness of GBM along the y-axis, the black lines represent the mean thickness of the GBM in each group. GBM, glomerular basement membrane; FPs, foot processes; *P < 0.05; ****P < 0.0001.
Podocytes invade into the GBM in Alport nephropathy
The appearance of cellular material within the GBM has been previously described. Erythrocytes have occasionally been captured traversing the GBM in thin basement membrane nephropathy22. Podocyte infolding in the GBM has also been described in a range of glomerular pathologies23 and more recent investigation of Col4a3−/− mice, suggested that cellular interposition in the GBM is of mesangial cell origin24. Mesangial cells are located between adjacent capillary loops and have contact with the GBM at the bases of the capillary loops (Supplementary Fig. S1a). In contrast, podocytes directly adhere to the GBM of the capillary walls (Fig. 1a,b). Using SBF-SEM we were able to identify cellular invasions within the GBM that connected to podocyte FPs (Fig. 4a,b and Supplementary Movies S4 and S5). These invasions were rarely observed in young adult Col4a3−/− mice (Fig. 5a) and were never observed in Col4a3+/− or wild type mice. In contrast, older adult and aged Col4a3−/− mice had frequent podocyte invasions (Figs 4c and 5b–d). The lengths of podocyte invasions were variable, but the mean length increased with age (Fig. 5e). Moreover, they occurred more frequently in thickened and abnormal areas of GBM in Col4a3−/− mice (Figs 4c and 5f). In aged Col4a3−/− mice the entire GBM had irregular thickness and all regions of the GBM contained podocyte invasions (Fig. 5d,f). Since it is known that the genetic background of Alport mice has an effect on the rate of disease progression25, we also investigated Col4a3−/− mice on a 129S1/Svlmj background with SBF-SEM. Here, similarly, we found evidence of podocytes invading into the GBM (Supplementary Fig. S2a–e).

Identification of podocyte FPs invading the GBM in Alport syndrome.
(a) Tracking of cellular material embedded within the GBM through 50 nm sections revealed that this material originates from podocyte FPs. (b) Tilted view of the model generated in A. showing the interaction of the invading FP with the BM. (c) Distribution of GBM invasions of podocyte origin (magenta) recorded in 10,000 μm3 glomerular volume. Podocyte FP invasions were not present in wild type adult or Col4a3+/− mice, they were rarely observed in young Col4a3−/mice, but were abundant in adult and aged Col4a3−/− mice. GBM, glomerular basement membrane.

Analysis of podocyte FP invasions in Alport mice.
(a) Reconstructed podocyte GBM interface in a young adult (6 week) Col4a3−/− mouse. Podocyte cell body (green) and FPs from a neighbouring podocyte (magenta) are shown. A rare example of two short podocyte FP invasions inside the GBM (blue) is shown in young Col4a3−/− mice (right hand panel). The GBM in this region in thickened relative to the rest of the GBM. (b) SBF-SEM reconstruction reveals two FPs invading an abnormally thickened area of GBM in an adult (16 week) Col4a3−/− mouse. (c) The number of FP invasions increase with age in Col4a3−/− mice. (d) FP invasions are frequent in aged (28 week) Col4a3−/− mice. The GBM is irregular in thickness, both thinned and thickened compared to young GBM. (e) Quantification of podocyte FP invasions. The violin plots describe the distributions for the length of podocyte invasion into the GBM along the y-axis, the black lines represent the mean length of podocyte GBM invasions in each group. (f) Quantification of GBM thickness at sites of podocyte invasion. The violin plots describe the distributions for the thickness of GBM at sites of podocyte invasion and sites where there is no podocyte invasion. The black lines represent the mean GBM thickness of each group. GBM, glomerular basement membrane; FPs, foot processes; *P < 0.05; ****P < 0.0001.
Podocyte-GBM invasion is a common feature in glomerular disease
To investigate whether podocyte invasion into the GBM is a unique feature of Alport Syndrome, or shared with other genetic glomerular diseases, we investigated two additional mouse models of human disease. MYO1E mutations in humans cause steroid resistant focal segmental glomerulosclerosis (FSGS)26. MYO1E encodes myosin1e, a class I myosin which is expressed by mouse and human podocytes26,27. Myo1e−/− mice develop FSGS and renal failure associated with morphological abnormalities in both podocytes and the GBM; we therefore investigated adult Myo1e−/− mouse glomeruli with SBF-SEM. Our analysis revealed that podocyte FPs were flattened, broadened and in many regions completely effaced (Fig. 6a–c). There were expanded areas of GBM that coincided with loss of podocyte FP morphology (Fig. 6d). Overall, there was a loss of podocyte FP density and a thickening of the GBM (Fig. 6f,g). Along with these classical pathological changes in podocyte and GBM ultrastructure, we also discovered prevalent podocyte FP invasion into the GBM (Fig. 7a, Supplementary Fig. S3a and Supplementary Movie S6).

3D analysis of Myo1e−/− and Ptpro−/− mouse glomerular structure.
(a) Transverse view of reconstructed GBMs (blue) and podocyte FPs (green and magenta) reveal broadened and flattened FPs in Myo1e−/− and Ptpro−/− mice. (b) Large expansions of GBM occur in Myo1e−/− and Ptpro−/− mice. (c) Focal areas of podocyte FP effacement coincide with thickened and irregular regions of GBM in Myo1e−/− and Ptpro−/− mice. (d) In many regions there is complete effacement of podocyte FPs in in Myo1e−/− and Ptpro−/− mice. (e) Quantification of podocyte FP density. The violin plots describe the distributions for the number of FPs per length of GBM along the y-axis, the black lines represent the mean number of FPs per length of GBM in each group. (f) Quantification of GBM thickness. The violin plots describe the distributions for the thickness of GBM along the y-axis, the black lines represent the mean thickness of the GBM in each group. GBM, glomerular basement membrane; FPs, foot processes; ****P < 0.0001.

Podocyte-GBM invasion is a common feature in glomerular disease.
(a) Reconstructed podocyte GBM invasion in Myo1e−/− mouse. Podocyte invasion (magenta) and GBM (blue) are shown. (b) Reconstructed podocyte GBM invasion in Ptpro−/− mouse. Podocyte invasion (green) and GBM (blue) are shown. (c) Quantification of podocyte FP invasions. The violin plots describe the distributions for the length of podocyte invasion into the GBM along the y-axis, the black lines represent the mean length of podocyte GBM invasions in each group. (d) Quantification of GBM thickness at sites of podocyte invasion. The violin plots describe the distributions for the thickness of GBM at sites of podocyte invasion and sites where there is no podocyte invasion. The black lines represent the mean GBM thickness of each group. GBM, glomerular basement membrane; FPs, foot processes; ****P < 0.0001.
Next we investigated a third mouse model of human glomerular disease. Glomerular epithelial protein 1 (GLEPP1) is a receptor tyrosine phosphatase expressed by podocytes. Mutations in PTPRO the gene that encodes GLEPP1 protein causes childhood-onset nephrotic syndrome in humans28. Ptpro−/− mice have impaired renal function and SEM has demonstrated that these mice have shortened podocyte FPs29. We investigated adult Ptpro−/− mice with SBF-SEM and found dramatic GBM expansions, overall GBM thickening and focal areas of reduced podocyte FP density (Fig. 6a–d,f,g). Furthermore we identified podocyte invasions into the GBM similar to those observed in Col4a3−/− mice and Myo1e−/− mice (Supplementary Movie S7). In contrast to Col4a3−/− and Myo1e mice, podocyte invasions into the GBM in Ptpro−/− mice were more slender and short (Fig. 7b and Supplementary Fig. S3b). In addition, podocyte invasions in Ptpro−/− mice were less variable in length than those in adult and aged Col4a3−/− mice (Fig. 7c). Similar to Col4a3−/− mice, podocyte invasions were associated with thickened regions of GBM in both Myo1e−/− and Ptpro−/− mice (Fig. 7d).
Mesangial cells invade the GBM at the mesangial aspect
Using 3D modelling of glomerular structure we also analysed the mesangial cells since they have been reported to invade the GBM in Alport syndrome24. Indeed, analysing SBF-SEM from Alport mice we identified mesangial invasions of varying length inside the GBM along the mesangial aspect of the glomerular capillary loop (Supplementary Fig. S1b and Supplementary Movies S9 and S10). These mesangial processes were distinct from podocyte invasions; they were only observed close to the mesangial angle and not in the peripheral capillary loops. We also detected mesangial invasions in both Myo1e−/− and Ptpro−/− mice (Supplementary Fig. S1c,d and Supplementary Movies S10 and S11). Mesangial invasions were rarer than podocyte invasions and in contrast to podocyte invasions, mesangial invasions were also present in wild type animals (Supplementary Movie S8). However, mesangial invasions were longer in mice with Col4a3, Myo1e and Ptpro mutations compared with wild type mice (Fig. S1E, Movie S9–11).
GBM defects resembling podocyte invasion in human Alport syndrome
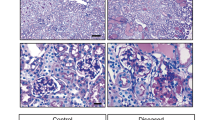
We interrogated TEM sections from the renal biopsies of two siblings that we have previously reported30. The proband was investigated for persistent microscopic haematuria and mild proteinuria and he had a renal biopsy at 3 years of age. This demonstrated abnormal GBM thickness, which prompted COL4 genetic screening. He was found to be hemizygous for two COL4A5 variants: c.2858G > T; p.(Gly953Val) and c.3097G > C; p.(Gly1033 Arg). His younger brother presented at the age of 6 months with steroid resistant nephrotic syndrome and was also hemizygous for the same COL4A5 variants. In addition he carried two homozygous missense variants in MYO1E: c.352A > G; p.(Lys118Glu) and c.2627C > G; p.(Thr876Arg). In both renal biopsies there was evidence of electron lucent regions within the GBM (Fig. 8). In addition, there was evidence of cellular material within the GBM and these resembled the podocyte invasions that we observed in Col4a3−/−, Myo1e−/− and Ptpro−/− mice. Future SBF-SEM studies of human biopsies are needed to confirm that the cellular material is indeed podocyte protrusions as we have shown in the mouse.

Evidence for podocyte invasion in patients with Alport syndrome.
Transmission electron microscopy of an Alport patient with mutant COL4A5 alleles c.2858G > T; p.(Gly953Val) and c.3097G > C; p.(Gly1033 Arg) (a) and a sibling who in addition to carrying COL4A5 variants had mutations in MYO1E: c.352A > G; p.(Lys118Glu) and c.2627C > G; p.(Thr876Arg) (b) These TEMs reveal features that resemble podocyte invasions identified in mouse models of glomerular disease.
Discussion
In this study we used SBF-SEM to analyze spatial relationships between cellular and extracellular glomerular components in health and disease. We identified podocyte invasions into the GBM as a novel morphological feature of glomerular disease. Irregular expansions of the urinary surface of the GBM were also a dominant feature in all disease models and these irregular expansions were more likely to harbor podocyte invasions. Podocyte invasions occurred in Alport (Col4a3−/−), Myo1e−/− and Ptpro−/− mice and lucent regions of GBM, resembling sites of podocyte invasion, were seen in patients with Alport syndrome. Finally, we demonstrate that GBM thickness and irregularity markedly increases and podocyte FP density decreases with glomerular disease progression.
The GBM is a critical component of the glomerular filtration barrier being exquisitely water permeable yet prohibiting the passage of macromolecules. Super resolution microscopy was recently used to describe the precise molecular topology of the GBM31. In this nanoscale investigation of the GBM, type IV collagen was shown to be restricted to the centre of the GBM. This localization indicates that the collagen IV network is too distant from podocytes to interact with integrins at the cell surface and therefore initiate outside-in signaling. This paper also demonstrated that type IV collagen α1α1α2 mislocalizes during Alport syndrome and is found adjacent to podocytes. Type IV collagen α1α1α2 that is proximal to podocytes could conceivably perturb cell-ECM interactions or alter the organization of other components within the GBM. In our investigation, we identified novel podocyte protrusions into the GBM and one explanation for podocyte invasion is that increased GBM degradation and subsequent podocyte migration occur in order to improve podocyte anchorage and to prevent detachment. A second, attractive explanation is that podocytes use invasions to remodel GBM damage that is caused by altered filtration forces. Potentially, podocyte invasions are extensions of plasma membrane used to remodel ECM, similar to fibropositors that regulate the direction of collagen fibril assembly during tendon repair24.
Our investigation of human GBM was limited to TEM sections since archival EM blocks from human biopsies are not routinely stained with heavy metals prior to embedding. For SBF-SEM staining is required prior to embedding since sectioning is automated within the microscope. In the human TEM we observed lucent regions in the GBM, resembling the regions in the mouse GBM that were tracked to podocyte protrusions and we therefore propose that the same protrusions occur in human glomerular disease. These observations are likely to overlap with the previously reported podocyte infolding in the GBM, seen across a range of glomerular pathologies23.
Defining the molecular composition and nature of podocyte invasions into the GBM would determine whether these could be therapeutically blocked or activated in order to protect glomerular function. There are a number of candidate pathways that could drive podocyte invasion into the GBM. One such pathway is integrin mediated adhesion signalling. Focal adhesion kinase (FAK), a key integrin signalling protein, has been shown to be activated in Alport mouse podocytes by ectopic laminin α232. This same study revealed that FAK induces matrix metalloproteinase (MMP) release from podocytes. This proposes an explanation for podocyte protrusion into the GBM following podocyte-mediated ECM degradation. Indeed TEM imaging provides evidence of an electro-lucent halo around FP invasions (Figure S4), which could be the result of active ECM degradation by podocytes.
In conclusion, SBF-SEM has provided enhanced visualization of the glomerulus and this has led to the identification of previously unseen pathological features of glomerular disease. 3D ultrastructural imaging revealed expected GBM abnormalities in addition to podocyte protrusions, resembling invadosomes, in the GBM. This novel finding provides new insight into disease mechanisms and suggests that a common matrix-adhesion pathway is activated regardless of the primary molecular insult. What remains unknown is whether podocytes actively migrate into the GBM or if these protrusions occur passively. Further study is necessary to understand how and why podocytes protrude into the GBM. Answers to these questions could lead to the development of targeted therapies that would offer new therapeutic avenues for glomerular disease.
Methods
Mouse models
All animal experiments were performed in accordance with the relevant institutional approvals. The Col4a3−/− mice have been reported previously33 and experiments on these mice conformed to the National Institutes of Health Guide for the Care and Use of Laboratory Animals and were approved by the Washington University Animal Studies Committee. For Myo1e−/− mice27 experiments were performed according to the protocol approved by the SUNY Upstate Animal Committee. Ptpro−/− mice29 were backcrossed onto a 129P3/J background34 and the experiments were approved by Heinrich Heine University, Düsseldorf. C57BL/6JOIaHsd male mice were used as WT controls. A minimum of 3 animals per group were analysed in this study.
Serial block face-scanning electron microscopy
Kidney samples were prepared for SBF-SEM as described previously17. In brief, mouse kidneys were cut into 1 mm cubes and fixed in situ by using 2% (wt/vol) glutaraldehyde (Agar Scientific, UK) in 0.1 M cacodylate buffer (pH 7.2); stained in 1% (wt/vol) osmium tetroxide, 1.5% (wt/vol) potassium ferrocyanide in 0.1 M cacodylate buffer, followed by 1% (wt/vol) thiocarbohydrazide. After washing, more osmium was added by staining in 1% (wt/vol) osmium tetroxide, and soaked in 1% (wt/vol) uranyl acetate overnight. The final staining step incubation was performed at 60 °C with lead aspartate pH 5.5 for one hour. Samples were dehydrated in ethanol and infiltrated in TAAB 812 hard resin. Tissue was mounted onto an aluminium cryo pin (Electron Microscopy Sciences, cat. no. 70446) using cyanoacrylate glue and all block surfaces trimmed at 90° using a glass knife or diamond trimming tool (Diatome). A gold coating was applied to the block to create a conductive surface. The block was placed in the Quanta 250 FEG (FEI Company) + Gatan 3view system and a 41 μm × 41 μm field of view was chosen and imaged by using a 4096 × 4096 scan, which gave an approximate pixel size of 10 nm. The section thickness was set to 50 nm in the Z (cutting) direction.
Transmission electron microscopy and quantification
Sample fixation and staining was performed as for SBF-SEM. Different blocks from the same sample were used for TEM and SBF-SEM. Tissue blocks were removed from their moulds and sectioned (70–80 nm thickness). Sections were examined using a FEI Tecnai 12G2 Biotwin transmission electron microscope. Magnification varied from 145X to 6800X. n = 6 glomeruli were imaged for each mouse model. Images were saved as original.dm3 files and.tiff files. The dm3 files were opened using Fiji/ImageJ software version 2.0.0-rc-29/1.49r. Collectively 60 randomly selected points of the GBM across 50.dm3 images were measured to assess GBM thickness. The straight-line option was used to draw across the width of the GBM at the chosen point > Analyse > Measure. To quantify podocyte FP number and FP protrusions 50 randomly selected areas of GBM 5 μm in length were measured as above and FP or protrusion number counted manually. Where protrusions were identified the thickness of the infiltrated GBM was also measured. All measurements were imported into GraphPad Prism software version 5.0 for interpretation and graphical representation.
3D modeling
Typically, Z volumes datasets comprised 1000 images (50 μm z depth). The IMOD beta version 4.7.11 suite of image analysis software was used to build image stacks, reduce imaging noise, and generate 3D reconstructions35. A shell script was run to float images to a common density range, following which they were converted to a size 8 bit smoothed data MRC file as reported previously17. A 24″ Wacom tablet with interactive pen tool was used to construct 3D models from the MRC file open in IMOD. The image stack was scrolled through to locate a feature of interest e.g. the GBM. The pen tool was positioned over this feature and a contour drawn around the entire structure. The image stack was moved up or down by one section and a second contour was drawn around the same feature. This process was repeated on every image for the desired number of sections.
Statistical analysis
170 measurements of podocyte FP density and 70 measurements of GBM thickness were taken per group. All identified invasions were quantified and invasion number was normalised to the total volume of glomerulus analysed. Comparisons between individual disease models and wild type were performed in GraphPad Prism using one way ANOVA with a Tukey’s post-test, p values less than 0.05 were considered to be significant.
Additional Information
How to cite this article: Randles, M. J. et al. Three-dimensional electron microscopy reveals the evolution of glomerular barrier injury. Sci. Rep. 6, 35068; doi: 10.1038/srep35068 (2016).
References
Hudson, B. G., Tryggvason, K., Sundaramoorthy, M. & Neilson, E. G. Alport’s syndrome, Goodpasture’s syndrome, and type IV collagen. The New England journal of medicine 348, 2543–2556, doi: 10.1056/NEJMra022296 (2003).
Gross, O. et al. Early angiotensin-converting enzyme inhibition in Alport syndrome delays renal failure and improves life expectancy. Kidney Int 81, 494–501, doi: 10.1038/ki.2011.407 (2012).
Barker, D. F. et al. Identification of mutations in the COL4A5 collagen gene in Alport syndrome. Science 248, 1224–1227 (1990).
Heiskari, N. et al. Identification of 17 mutations in ten exons in the COL4A5 collagen gene, but no mutations found in four exons in COL4A6: a study of 250 patients with hematuria and suspected of having Alport syndrome. Journal of the American Society of Nephrology: JASN 7, 702–709 (1996).
Kalluri, R. et al. A COL4A3 gene mutation and post-transplant anti-alpha 3(IV) collagen alloantibodies in Alport syndrome. Kidney international 47, 1199–1204 (1995).
Boye, E. et al. Determination of the genomic structure of the COL4A4 gene and of novel mutations causing autosomal recessive Alport syndrome. American journal of human genetics 63, 1329–1340, doi: 10.1086/302106 (1998).
Storey, H., Savige, J., Sivakumar, V., Abbs, S. & Flinter, F. A. COL4A3/COL4A4 mutations and features in individuals with autosomal recessive Alport syndrome. J Am Soc Nephrol 24, 1945–1954, doi: 10.1681/ASN.2012100985 (2013).
Abrahamson, D. R., Hudson, B. G., Stroganova, L., Borza, D. B. & St John, P. L. Cellular origins of type IV collagen networks in developing glomeruli. Journal of the American Society of Nephrology: JASN 20, 1471–1479, doi: 10.1681/ASN.2008101086 (2009).
Suh, J. H. & Miner, J. H. The glomerular basement membrane as a barrier to albumin. Nat Rev Nephrol 9, 470–477, doi: 10.1038/nrneph.2013.109 (2013).
Lennon, R. et al. Global analysis reveals the complexity of the human glomerular extracellular matrix. J Am Soc Nephrol 25, 939–951, doi: 10.1681/ASN.2013030233 (2014).
Kalluri, R., Shield, C. F., Todd, P., Hudson, B. G. & Neilson, E. G. Isoform switching of type IV collagen is developmentally arrested in X-linked Alport syndrome leading to increased susceptibility of renal basement membranes to endoproteolysis. J Clin Invest 99, 2470–2478, doi: 10.1172/JCI119431 (1997).
Byron, A. et al. Glomerular Cell Cross-Talk Influences Composition and Assembly of Extracellular Matrix. Journal of the American Society of Nephrology: JASN, doi: 10.1681/ASN.2013070795 (2014).
Gunwar, S. et al. Glomerular basement membrane. Identification of a novel disulfide-cross-linked network of alpha3, alpha4, and alpha5 chains of type IV collagen and its implications for the pathogenesis of Alport syndrome. The Journal of biological chemistry 273, 8767–8775 (1998).
Kalluri, R., Shield, C. F., Todd, P., Hudson, B. G. & Neilson, E. G. Isoform switching of type IV collagen is developmentally arrested in X-linked Alport syndrome leading to increased susceptibility of renal basement membranes to endoproteolysis. The Journal of clinical investigation 99, 2470–2478, doi: 10.1172/JCI119431 (1997).
Savige, J. et al. Expert guidelines for the management of Alport syndrome and thin basement membrane nephropathy. J Am Soc Nephrol 24, 364–375, doi: 10.1681/ASN.2012020148 (2013).
Shankland, S. J. The podocyte’s response to injury: role in proteinuria and glomerulosclerosis. Kidney international 69, 2131–2147, doi: 10.1038/sj.ki.5000410 (2006).
Starborg, T. et al. Using transmission electron microscopy and 3View to determine collagen fibril size and three-dimensional organization. Nat Protoc 8, 1433–1448, doi: 10.1038/nprot.2013.086 (2013).
Yeung, C. Y. et al. Chick tendon fibroblast transcriptome and shape depend on whether the cell has made its own collagen matrix. Scientific reports 5, 13555, doi: 10.1038/srep13555 (2015).
Kalson, N. S. et al. A structure-based extracellular matrix expansion mechanism of fibrous tissue growth. eLife 4, doi: 10.7554/eLife.05958 (2015).
Taylor, S. H. et al. Matrix metalloproteinase 14 is required for fibrous tissue expansion. eLife 4, e09345, doi: 10.7554/eLife.09345 (2015).
Ichimura, K. et al. Three-dimensional architecture of podocytes revealed by block-face scanning electron microscopy. Scientific reports 5, 8993, doi: 10.1038/srep08993 (2015).
Collar, J. E., Ladva, S., Cairns, T. D. & Cattell, V. Red cell traverse through thin glomerular basement membranes. Kidney Int 59, 2069–2072, doi: 10.1046/j.1523-1755.2001.00721.x (2001).
Joh, K. et al. Proposal of podocytic infolding glomerulopathy as a new disease entity: a review of 25 cases from nationwide research in Japan. Clin Exp Nephrol 12, 421–431, doi: 10.1007/s10157-008-0104-z (2008).
Zallocchi, M., Johnson, B. M., Meehan, D. T., Delimont, D. & Cosgrove, D. alpha1beta1 integrin/Rac1-dependent mesangial invasion of glomerular capillaries in Alport syndrome. The American journal of pathology 183, 1269–1280, doi: 10.1016/j.ajpath.2013.06.015 (2013).
Cosgrove, D., Kalluri, R., Miner, J. H., Segal, Y. & Borza, D. B. Choosing a mouse model to study the molecular pathobiology of Alport glomerulonephritis. Kidney international 71, 615–618, doi: 10.1038/sj.ki.5002115 (2007).
Mele, C. et al. MYO1E mutations and childhood familial focal segmental glomerulosclerosis. N Engl J Med 365, 295–306, doi: 10.1056/NEJMoa1101273 (2011).
Krendel, M. et al. Disruption of Myosin 1e promotes podocyte injury. J Am Soc Nephrol 20, 86–94, doi: 10.1681/ASN.2007111172 (2009).
Ozaltin, F. et al. Disruption of PTPRO causes childhood-onset nephrotic syndrome. American journal of human genetics 89, 139–147, doi: 10.1016/j.ajhg.2011.05.026 (2011).
Wharram, B. L. et al. Altered podocyte structure in GLEPP1 (Ptpro)-deficient mice associated with hypertension and low glomerular filtration rate. J Clin Invest 106, 1281–1290, doi: 10.1172/JCI7236 (2000).
Lennon, R. et al. Coinheritance of COL4A5 and MYO1E mutations accentuate the severity of kidney disease. Pediatric nephrology 30, 1459–1465, doi: 10.1007/s00467-015-3067-9 (2015).
Suleiman, H. et al. Nanoscale protein architecture of the kidney glomerular basement membrane. eLife 2, e01149, doi: 10.7554/eLife.01149 (2013).
Delimont, D. et al. Laminin alpha2-Mediated Focal Adhesion Kinase Activation Triggers Alport Glomerular Pathogenesis. Plos One 9, e99083, doi: 10.1371/journal.pone.0099083 (2014).
Lin, X., Suh, J. H., Go, G. & Miner, J. H. Feasibility of repairing glomerular basement membrane defects in Alport syndrome. J Am Soc Nephrol 25, 687–692, doi: 10.1681/ASN.2013070798 (2014).
Gonzalez-Brito, M. R. & Bixby, J. L. Protein tyrosine phosphatase receptor type O regulates development and function of the sensory nervous system. Mol Cell Neurosci 42, 458–465, doi: 10.1016/j.mcn.2009.09.009 (2009).
Kremer, J. R., Mastronarde, D. N. & McIntosh, J. R. Computer visualization of three-dimensional image data using IMOD. Journal of structural biology 116, 71–76, doi: 10.1006/jsbi.1996.0013 (1996).
Acknowledgements
MJR was supported by a project grant awarded to RL and JHM from Kidney Research UK (RP52/2014). SC was supported by an intercalation award from Kidney Research UK (ST/ID12/2014) and a Wolfson Foundation award (YLR/eac/19928). MK is supported by the National Institute of Health NIDDK award R01DK083345 and we acknowledge technical help from S.E. Chase at SUNY Upstate Medical University. JHM is supported by grants from the NIH (R01DK078314 and R56DK100593) and the American Heart Association (14GRNT20370035). RL is supported by a Wellcome Trust Intermediate Clinical Fellowship (090006). We also acknowledge core funding from the Wellcome Trust (088785/Z/09/Z) awarded to the Wellcome Trust Centre for Cell-Matrix Research, University of Manchester, UK.
Author information
Authors and Affiliations
Contributions
M.J.R., S.C. and R.L. designed the study. T.S., A.M. and K.E.K. assisted with electron microscopy imaging and analysis, M.K., E.K., L.S. and J.H.M. provided tissue samples. All authors contributed to academic discussion and preparation of the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Randles, M., Collinson, S., Starborg, T. et al. Three-dimensional electron microscopy reveals the evolution of glomerular barrier injury. Sci Rep 6, 35068 (2016). https://doi.org/10.1038/srep35068
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep35068
This article is cited by
-
Complexities of the glomerular basement membrane
Nature Reviews Nephrology (2021)
-
Podocyte penetration of the glomerular basement membrane to contact on the mesangial cell at the lesion of mesangial interposition in lupus nephritis: a three-dimensional analysis by serial block-face scanning electron microscopy
Clinical and Experimental Nephrology (2019)
-
Subcellular connectomic analyses of energy networks in striated muscle
Nature Communications (2018)
-
Ultrastructural Characterization of the Glomerulopathy in Alport Mice by Helium Ion Scanning Microscopy (HIM)
Scientific Reports (2017)
-
New structural insights into podocyte biology
Cell and Tissue Research (2017)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
