
Abstract
Anthropogenic landscape transformation has an important effect on vector-borne pathogen transmission. However, the effects of urbanization on mosquito communities are still only poorly known. Here, we evaluate how land-use characteristics are related to the abundance and community composition of mosquitoes in an area with endemic circulation of numerous mosquito-borne pathogens. We collected 340 829 female mosquitoes belonging to 13 species at 45 localities spatially grouped in 15 trios formed by 1 urban, 1 rural and 1 natural area. Mosquito abundance and species richness were greater in natural and rural areas than in urban areas. Environmental factors including land use, vegetation and hydrological characteristics were related to mosquito abundance and community composition. Given the differing competences of each species in pathogen transmission, these results provide valuable information on the transmission potential of mosquito-borne pathogens that will be of great use in public and animal health management by allowing, for instance, the identification of the priority areas for pathogen surveillance and vector control.
Similar content being viewed by others


Introduction
Vector-borne diseases are an important public health concern1 since both the distribution and incidence of many diseases have increased in recent decades2. With over 3,500 species worldwide, mosquitoes are by far the most relevant vectors of pathogens3 such as metazoan (e.g. filarial nematodes), protozoan (e.g. malaria parasites) and numerous viruses (e.g. Dengue virus, Rift Valley virus and West Nile virus).
The transmission dynamics of vector-borne pathogens are determined by the interactions between the pathogens, vectors and vertebrate hosts, and are influenced by environmental and socio-ecological drivers4. Factors such as vector abundance and species distribution are crucial for determining the distribution and incidence of vector-borne diseases5. Vector community composition may also affect pathogen transmission given that not all species are competent vectors for a pathogen6 and individual vector species play different roles in the transmission cycles of the pathogen. Some species may facilitate the transmission between reservoir vertebrate hosts, while others may act as bridges between infected competent vertebrates and humans or other susceptible species7. Moreover, a published model of pathogen transmission dynamic in relation to host and vector biodiversity concluded that heterogeneity in the susceptibility of the reservoir species could dilute the pathogen transmission, while greater vector species richness may amplify its circulation8. In this work, Roche et al. suggested, using mathematical models, that both mosquito and bird species diversity are important for the transmission dynamics of vector-borne pathogens (e.g. West Nile virus (WNV))8.
Understanding how environmental variables influence the distribution of mosquitoes is a key issue in disease ecology, especially since anthropogenic changes in climate and landscape (global change) are now affecting the distribution and incidence of vector-borne pathogens9. It is widely accepted that environmental characteristics affect mosquito distribution10,11 since each mosquito species has certain habitat requirements, which can vary greatly even between closely related species12.
Human activities produce substantial ecological disturbances that affect communities of both vertebrate and invertebrate organisms13,14, and often lead to an increase in the abundance of a few species and a general loss of biodiversity15. Therefore, the increased dominance of a few key species, both vector and/or host species, and the ecological conditions (e.g. anthropized environments) could favour their interspecific contact rates (i.e. increased biting frequency) and pathogen transmission rates16. Factors including the simplification of habitat structures17 and the alteration of trophic interactions18 may govern anthropogenic-mediated loss of biodiversity in urban areas. Changes in resource availability, vegetation coverage, the characteristics of water bodies, and both temperature and rainfall patterns may all have a severe impact on mosquito populations12,19,20 and thus directly and/or indirectly affect their community ecology14 and the pathogen transmission risk (reviewed in LaDeau et al.21,22).
Mosquito communities in urban and rural landscapes are generally characterized by a lower mosquito diversity and/or abundance than in natural areas12,23,24. Although marshlands and other temporary flooded areas are generally absent from urban areas, human activity creates artificial habitats such as water deposits, swimming pools, urban sewage systems, gardens, and subterranean and storm water systems that act as alternative breeding sites for mosquitoes. These habitats appear to be suitable for Anopheles25, Culex26 and Aedes mosquitoes27, especially during the dry season when surface water is otherwise scarce. Therefore, the effects of urbanization may vary depending on the mosquito species as some species may be favoured by anthropogenic environmental changes13. By contrast, other species may react differently to such change and their abundances may decrease on an urban-to-rural gradient. For instance, Culex mosquitoes were more common in urban sites while mosquitoes of the Mansonia genus predominated in rural habitats10. In addition to these species-specific responses, mosquito community composition and abundance may also play an important role in pathogen transmission7,28. The varying degrees of implication of each species in pathogen transmission depend on their vectorial competence, blood-feeding behaviour and life history traits. Therefore, it is vital to identify the ecological factors that drive the abundance of mosquito vectors of pathogens and thus ultimately may affect pathogen transmission.
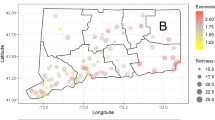
Here, we integrate field data collected at 45 sites along an urbanization gradient to assess the impact of land use on mosquito abundance and community composition (Fig. 1). The study was conducted in areas with differing degrees of anthropization ranging from natural areas with little human-driven landscape transformation, to rural areas characterized by the presence of livestock and anthropized urban areas with dominance of human presence. We expect that results from this study allow to better understand how landscape characteristics are related to the amplification and transmission dynamics of different mosquito-borne pathogens such as heartworms29, West Nile and Usutu viruses30,31, and avian malaria parasites32 that are endemic in southwest Spain.

This map was created using ArcGIS v10.2.1 (ESRI, Redland).
Results
Mosquito abundance and species composition
A total of 340 829 female mosquitoes belonging to 13 species and five genera were collected. The most trapped species were Culex theileri Theobald (282 891 ind.), Ochlerotatus caspius Pallas (21 155), and Culex pipiens Linnaeus (19 268), followed by Culex perexiguus Theobald (5,939), Anopheles atroparvus Van Thiel (5,387), Culiseta annulata Schrank (2,514), Ochlerotatus detritus Haliday (1,495), Culex modestus Ficalbi (1,237) and Culiseta longiareolata Marcquart (476). Most Anopheles atroparvus were captured at a single study site, the Doñana National Park, where 3,207 individuals were collected. We also captured a number of additional species in very low number: Anopheles algeriensis Theobald (41), Ochlerotatus berlandi Seguy (22), Culiseta subochrea Edwards (13) and Urotaenia unguiculata Edwards (6). Finally, 216 Anopheles spp., 167 Culex spp. and two Culiseta spp. could only be identified to genus level. Mosquitoes not identified to species level were excluded from species richness and diversity index calculations.
The average number of mosquito captured per trap night was 424.33 (range 4.28–5,780.89), the average species richness per locality was 6.75 (range 2–10) and the average mosquito diversity per locality was 0.41 (range 0.06–0.71), see data for each locality in Supplementary Table S1.
Environmental effects on mosquito community
General Linear Mixed Models (GLMM) indicate that mosquito abundance and species richness were similar in natural and in rural areas but were greater than in urban areas (Table 1). The captures of each of the commonest mosquito species were lower in urban areas than in natural ones. However, the abundance of Cx. modestus and Cx. perexiguus were similar in rural and both urban and natural areas. A significantly lower abundance of An. atroparvus, Cx. theileri and Oc. caspius was found in urban than both rural and natural areas (Table 1). The abundance of Cx. pipiens was similar in urban and rural areas but lower than in natural areas (Table 1). Cx. pipiens was by far the most abundant mosquito species in urban areas, while Cx. theileri was the most abundant species in rural areas (Table 1).
Random Forest (RF) results support a close association between landscape and hydrological characteristics, Normalized Difference Vegetation Indices (NDVI), and mosquito community variables (Table 2). The best models for total mosquito abundance and species richness were those that included environmental variables measured at the 1000 m and 250 m radii buffers, respectively. The total abundance of mosquitoes was positively associated with the area occupied by wetlands (Fig. 2a) but negatively associated with both the area of urban land (Fig. 2b) and the human population (Fig. 2c, Table 2). Similarly, mosquito species richness was negatively associated with the area occupied by urban land (Fig. 2d), the human population (Fig. 2e) and the distance to marshlands (Fig. 2f, Table 2). Table 2 summarizes the most important variables explaining the abundance of the commonest mosquito species sampled. Cx. theileri was the most abundant species in the studied areas and its abundance was negatively related with the area occupied by urban land (Fig. 2g) but positively related to the area occupied by wetlands (Fig. 2h, Table 2). The abundance of the majority of mosquito species was positively related to summer NDVI. Positive associations were found between the abundance of An. atroparvus, Cx. modestus and Cx. perexiguus, Cx. theileri and the summer NDVI (Fig. 2i), and the abundance of Cx. perexiguus and autumn NDVI (see Supplementary Fig. S1 for further details). Cx. modestus abundance was negatively related to winter NDVI (Supplementary Fig. S1). Additionally, the abundance of Cx. modestus and Oc. caspius was negatively related to the distance from marshlands (Supplementary Fig. S1). We did not find any association between the diversity of mosquitoes and the abundance of Cx. pipiens, and the independent variables included in the RF.

Partial dependence plot for mosquito log-transformed captures and: (a) the percentage of land area covered by wetlands (log ratio transformed); (b) the percentage of land area covered by urban areas (log ratio transformed); (c) human population density (log-transformed). Partial dependence plot for species richness (number of different species) and: (d) the percentage of land area covered by urban areas (log ratio transformed); (e) human population density (log-transformed); (f) the distance to the nearest marshland (m). Partial dependence plot for Cx. theileri captures and: (g) the percentage of land area covered by urban areas (log ratio transformed); (h) the percentage of land area covered by wetlands (log ratio transformed); (i) the summer NDVI index. Partial dependence is the dependence of the probability of presence of one predictor variable after averaging out the effects of the other predictor variables in the model.
Discussion
To understand the risk of transmission of vector-borne pathogens it is essential to identify the environmental and biological factors determining the abundance and species composition of mosquito communities33. Measuring mosquito abundance in the field is time consuming and financially expensive and so remote sensing variables provide an effective way for identifying which areas are most prone to harbouring significant populations of certain mosquito species34. In this study, we found associations between mosquito abundance and richness, and land use in an area of Mediterranean climate, in which several pathogens affecting humans, wildlife and livestock are circulating (e.g. Figuerola et al.30).
We found strong support for the impact of urbanization on mosquito abundance and community composition and identified some key environmental variables that potentially affect these associations. Mosquito abundance and species richness were higher in natural and rural areas than in urban ones. These results agree with those from previous studies conducted in Europe24 and elsewhere35 where anthropogenic habitats usually show the lowest abundance of mosquitoes. Natural areas, with freshwater and brackish water wetlands, are more favourable breeding environments, in spite those urban areas may provide suitable habitats for some particular mosquito species (e.g. a wide range of breeding sites in artificial containers and recipients)9,36. According to our results, mosquito abundance rose as the area occupied by wetlands increased, but fell as the area occupied by urban areas and populated areas increased. Similarly, in South Australia the abundance of different mosquito species increased in areas further from the city centre and closer to saltmarshes35. A similar pattern was found for mosquito species richness, which was negatively correlated to factors such as the area occupied by urban land, human density and the distance between urban areas and marshlands. In addition, larvicide treatments with Bacillus thurigiensis are carried out in urban areas of Huelva to reduce the nuisance to human populations caused by mosquitoes (S. Ruiz pers. com.) and may have potentially reduced the mosquito populations in some of the studied urban areas. Salt marshes may provide a suitable environment for halophytic species of mosquito35,37 and the presence of this habitat strongly affected the abundance of species such as Oc. caspius in this study. Interestingly, the relationship between mosquito abundance and distance to marshlands was not lineal and had marked thresholds at distances of about 2.5 km for Cx. modestus and 10 km for Oc. caspius. These differences fit well with the estimated flying distances of mosquitoes, which range between 0.16 km and 1.98 km for Culex (e.g. Cx. pipiens38) and up to 12 km for Oc. caspius39. According to our results, mosquito abundance and species richness decreased as human population density rose. In this case, the relationship between mosquito abundance and human density had a threshold at areas with approximately 50 habitants/250 m2. Previous studies have found strong support for the effect of human population on the transmission of vector-borne pathogens40. This may be, at least in part, due to the impact that the reduction in the availability/suitability of breeding areas and the implementation of mosquito control in more densely populated areas have on native mosquito communities. On the other hand, certain invasive species such as Aedes albopictus seem to prefer anthropically altered areas to natural landscapes41, but fortunately this invasive species, that is colonizing southern and eastern Spain42, was not detected during this study.
Close associations were found between the abundance of the commonest mosquito species and environmental characteristics, although these associations varied between species. In particular, we found a general positive relationship between the abundance of the commonest mosquito species and NDVI (an index of photosynthetic activity), especially during the summer season. Higher values of NDVI reflect a higher vegetation covertures and potentially a higher availability of breeding and, especially, resting habitats for mosquitoes and other insect vectors43. Previous studies both in USA and Europe have also reported positive associations between NDVI values and mosquito presence, abundance and diversity44,45,46.
Culex theileri, the most abundant species trapped in the study area, was extremely abundant in natural and rural areas. Previous studies have found that this species typically feed on blood from livestock and wild and feral mammals28. For instance, Cx. theileri represent an important vector for Dirofilaria sp.47 and its great abundance in rural areas may explain the high prevalence of dirofilariasis in the study area29. Moreover, the abundance of Cx. theileri rose as the area occupied by wetlands increased but fell where urban areas occupied more land. This fact may be due to the breeding requirements of this species. Although it is able to use artificial habitats and heavily polluted water, Cx. theileri usually occurs in flooded meadows, stagnant or slowly moving streams, rock pools, swamps and rice paddies, which probably explains its greater abundance in wetland natural areas12. Furthermore, Cx. theileri is the more abundant mosquito in May–July in the study area32,44, when the environmental conditions reflected by the summer NDVI index (higher vegetation cover during the summer season) are higher. In addition to the higher abundance of this species close to wetlands due to the suitability of breeding areas, rural and natural areas may also provide a higher abundance of potential hosts for this species.
Anopheles atroparvus is the only species of mosquito competent for malaria transmission to humans that is present in Spain48. It was the most abundant species in rural areas and, similarly to Cx. theileri, this species was positively associated with both the area occupied by wetland habitats and greater summer NDVI. Roiz et al.44 reported a positive relationship between NDVI and inundation area and An. atroparvus abundance in the Doñana National Park. In fact, An. atroparvus prefers to breed in not polluted sites, with a slight preference for brackish water. This species is commonly found in canals, river margins and paddy fields12, which may explain the higher abundance of this species close to wetlands. Moreover, the abundance of An. atroparvus was positively related to summer NDVI and its abundance also peaks in summer44.
The abundance of Cx. modestus, Cx. perexiguus and Oc. caspius was greater in natural than in urban environments. Moreover, Cx. modestus, was more abundant both in wetlands and in areas with higher summer NDVI (but lower winter NDVI), and its abundance fell as the distance to marshlands increased. This positive relationship with summer NDVI and negative relationship with winter NDVI probably is due to the preference of this species to breed in rice-fields that remain unvegetated during the winter and had very high vegetation cover during the summer season. This species is mainly found in brackish marshes and flooded paddy fields12, which are especially abundant in the study area49. According to Ponçon et al.50, this species prefers shallow sunlit habitats and is frequent in meadows and irrigation channels. Analogous results were also found in marshlands from southern Spain44, mostly during summer, which may explain, at least in part, the negative association between its abundance and the distance to marshlands. Thus, these results support the potential role of rice paddies as both major sources of mosquito-induced nuisance and an important factor in disease transmission44. Furthermore, the abundance of Cx. perexiguus, a key link in the epizootic transmission of WNV to horses in southern Spain28, was positively related to zones with greater summer and autumn NDVIs. In our study, Cx. perexiguus abundance was negatively related to the area occupied by urban land, explaining the higher abundance of this species in natural than urban and rural areas. Cx. perexiguus use to breed in stagnant and ephemeral ponds such as swamps, streams, and paddy fields, usually with emergent vegetation especially during later summer and autumn when the abundance of this species peak44 while it is usually less abundant during spring32. Also, we found strong negative relationship between the abundance of the salt marsh species Oc. caspius and the distance to the marshlands. This result supports the finding of Leisnham et al.37 who reported the influence of tide heights on the abundance of this species. Oc. caspius prefers to breed in natural environments12,24 and usually use salt marshes as breeding sites and areas for larval development44. This fact may also explain the higher abundance of this species found in natural habitats in our study. This species is considered a secondary epizootic vector for different and novel flaviviruses of potential medical concern isolated in Spain51. Finally, Cx. pipiens, an important vector of a large number of viruses and other pathogens including WNV52, was the most abundant of all mosquito species in urban areas where it has been suggested to play a key role in the epizootic transmission of pathogens to humans53. This mosquito species is commonly found in urbanized areas52,54,55,56 where it uses water bodies like vases in cemeteries, small clay pots or water outlets for oviposition and larvae are frequently found in human-made water bodies55. In fact, among the six mosquito species analysed, Cx. pipiens was the most abundant in urban areas, although its abundance was significantly higher in natural habitats. In The Netherlands, Cx. pipiens was a widespread species being present different habitats including wetlands, agricultural and urban sites24. In the study area, natural areas provide more suitable conditions for this species. Unfortunately, we failed to identify any significant relationships with none of the analysed variables and the Cx. pipiens abundance. This fact is probably due to the incapacity of the spatial resolution of the cartography used here to identify the relevant variables for the ecology of this species. Additionally, the abundance of this species may be strongly influenced by distribution of human infrastructures (i.e. the design of water outlets in the streets) and activities not measured in this analysis.
In conclusion, in this study we provide strong support for the assertion that the anthropization of the landscape is an important factor determining the abundance and community composition of native mosquito species in an area with a Mediterranean climate. In particular, the mosquito species studied here may be implicated in the transmission of diseases affecting humans, wildlife and livestock. Therefore, results reported here on the effect of environmental factors affecting the abundance of particular mosquito species and, given the differing competence of these species in the transmission of pathogens, may provide valuable information for public health management and mosquito control by allowing the identification of priority areas for pathogen surveillance and/or vector control.
Materials and Methods
Study areas
Our field sites were located in Andalusia (S Spain) (Fig. 1), an area characterized by a Mediterranean climate, with a long dry summer season and greatest precipitation levels in winter. Mosquitoes were captured at 45 different localities in Cádiz, Huelva and Sevilla provinces (15 localities in each province). The sampling sites were grouped in three geographically close localities (named trios). Each trio included one locality in a natural habitat, one locality in a rural habitat and one locality in an urban habitat according to their environmental characteristics. Urban habitats contained more densely populated areas than the other two habitat types; rural habitats had more heads of livestock than the other two habitat types; natural habitats were selected on the basis of both lower human and livestock densities than in the other two habitat types, and an overall better conserved landscape. The three habitat categories were selected based upon visual inspection of the locations. In order to enhance the statistical power of our analyses, we compared the three habitat categories (natural, rural and urban areas) within the same trios to control for any geographically structured factor that may influence mosquito distribution or abundance.
Mosquito sampling and identification
From April to December 2013, mosquitoes were captured using BG-sentinel traps baited with BG-lure and dry ice as a source of CO2, which are known to be very effective for characterizing mosquito diversity and abundance56. This period covers the maximum mosquito activity in southern Spain32,44. Three traps were operated for 24 hours at each sampling site. Overall, 135 traps (3 traps x 45 localities), with a mean distance between traps of 119 m (range 20–636 m), were employed during each mosquito trapping session. Mosquito sampling was conducted once every 45 days and the 5–6 trapping sessions conducted at each site gave an overall trapping effort of 810 trap nights.
Adult mosquitoes were preserved in dry ice and stored frozen until identification. Mosquitoes were separated over a filter paper on a Petri plate on a chill table under a stereomicroscope. Mosquitoes were sorted by gender and date of collection, and then morphologically identified to species level following Becker et al.12 and Schaffner et al.57. Specimens belonging to the univittatus complex were identified as Culex perexiguus based on male genitalia as per Harbach58. When several thousands of mosquitoes were captured per trap per night, once we had identified 500 individuals, five groups of 100 individuals were weighted to the nearest 0.001 g, the total number of mosquitoes was estimated from sample weight and the proportion of identified individuals of each species was extrapolated for the rest of the sample.
Analyses were conducted using the following dependent variables calculated for each sampling site: i) total abundance of female mosquitoes, measured as the mean over the whole study period of the mean number of mosquito females of each species captured at each locality on each night, ii) mosquito species richness, measured as the total number of species captured, iii) mosquito diversity estimated using the Simpson index, and iv) the abundance of each of the five most abundant mosquito species – in addition to Cx. modestus – that plays an important role in the transmission of pathogens such as WNV59.
Remote sensing variables
Normalized difference vegetation indices (NDVI) were estimated from MODIS sensor images at a spatial resolution of 250 m and a temporal resolution (MOD13Q1 product) of 16 days. Overall, 23 images were obtained to estimate the mean NDVI for each season: winter (images from 03/12/2012 to 18/02/2013), spring (06/03/2013 to 25/05/2013), summer (10/06/2013 to 29/08/2013) and autumn (14/09/2013 to 17/11/2013).
Subsequently, the hydrological and land use information was obtained from cartography accessible at http://www.juntadeandalucia.es/institutodeestadisticaycartografia/DERA/ using ArcGIS v10.2.1 (ESRI, Redland). For the hydrological variables, we measured the distance from each mosquito trap to different water sources (e.g. the distance to the nearest river, marshland patch, stretch of freshwater, the coast and reservoirs), and then calculated the mean distances of the three traps at each of the 45 localities. As well, we calculated the shortest distance to any kind of water source according to the information obtained from the previously described variables. In addition, we quantified land use for different buffers around each mosquito trap using spatial analyses and zonal statistical tools for raster files and the geoprocessing intersect tool for vector variables. The mean value from the three traps at each of the 45 localities was quantified. We considered five different buffers around each trap point with radii of 100, 250, 500, 1000 and 2000 m, which embraced most of the daily flight range of Culex mosquitoes (e.g. Cx. pipiens38). The 33 different land use categories were grouped into four classes: agricultural, forests, wetlands and urban lands (see Supplementary Table S2), and the percentage of the total area occupied by each category was recorded. These land use variables add to 100% and for this reason log-transformed ratios were used for the statistical analyses60.
Human density was estimated as the number of people living in a grid of 250 × 250 m developed by the Institute of Statistics and Cartography of Andalusia. The population of each grid cell was taken to be the number of residents registered there on 1 January 2013 according to the local census Base de Datos Longitudinal de Población de Andalucía. See Supplementary Table S3 for further details about the variables included in this study.
Statistical analyses
Firstly, the differences in mosquito abundance, species richness and diversity and the abundance of the six commonest mosquito species between urban, rural and natural areas were tested with General Linear Mixed Models (GLMM). We performed a GLMM for each of the dependent variables, fitted by maximum likelihood functions with Gaussian distribution (package lme4 in R software), including habitat category as a fixed factor and ‘province’ and ‘trio’ as random factors to account for the geographic stratification present in our sampling design.
Secondly, to identify the relationship between environmental variables and mosquito abundance, richness and diversity and the abundance of the six commonest mosquito species we used Random Forest (RF) regression analyses based on 5,000 trees61. Random Forest represent iterations of regression trees, whereby both records and predictor variables are randomly permuted to assess the robustness of the derived classifications. This non-parametric algorithm method was used because RF procedures do not require the use of any particular model, which might be difficult to assign given the high number of independent variables. The advantage of RF models is their ability to predict a continuous (as in our case) rather than categorical (presence/absence) variable across a landscape, as well as their ability to assess the relative importance of each variable by predicting a complex model of interactions62. No a priori assumptions are made about the relationship between predictor and response variables, thereby allowing for the possibility of non-linear relationships.
In the RF analyses, we first included environmental data from the five buffers considered in each model in order to identify the best buffer selected for each variable (see the Supplementary Table S3 for further information about the predictors included in the models). After that, we conducted a second RF analysis including only data from the best-selected buffer using %IncMSE (percentage of increase in Mean Square Error) splitting criterion to find the optimal predictors. Finally, we ran a model selection procedure using the VSURF function to identify and plot the set of variables that most influenced our models. Trio was included as a stratification factor in all of these analyses. For consistence between the GLMM and the RF analyses, we have used the mean number of mosquito captured at each locality instead of data at the lowest level (considering each mosquito sampling as an independent sample). GLMM’s build using each mosquito trap/night instead of mean values did not differ qualitatively of the results presented here.
The normal distribution of all dependent and independent variables were checked. Moreover, the normal distribution of model residuals was also tested by using qq plots in R software. In all cases, residuals followed a normal distribution. Variables were transformed when necessary to reduce the influence of extreme values. Total counts of mosquito, counts of the six mosquito species and human density were log-transformed to normalize its distribution and stabilize the variance and to deal with differences of several orders of magnitude between sampling sites. Statistical analyses were conducted with R version 2.14.2 (R Development Core Team 2005) using the vegan, lme4, car, arm, MuMIn, randomForest and VSURF packages.
Ethics statement
Mosquito trapping was carried out with all the necessary permits issued by the regional Department of the Environment (Consejería de Medio Ambiente, Junta de Andalucía). Entomological surveys and sampling on private land and in private residential areas were conducted with all the necessary permits and consent, and in the presence of owners. This study did not affect any endangered or protected species.
Additional Information
How to cite this article: Ferraguti, M. et al. Effects of landscape anthropization on mosquito community composition and abundance. Sci. Rep. 6, 29002; doi: 10.1038/srep29002 (2016).
References
Jones, K. E. et al. Global trends in emerging infectious diseases. Nature 451, 990–993 (2008).
Gubler, D. J. The 20th century re-emergence of arboviral diseases: lessons learned and prospects for the future, in Proceedings of the Eighth Sir Dorabji Tata Symposium on Arthropod Borne Viral Infections, Bangalore, India (eds Raghunath, D. & Durga Rao, C.) Sir Dorabji Tata Centre for Research in Tropical Diseases (Tata McGraw-Hill Publishing Company Limited, Bangalore, India, 2008).
Tolle, M. A. Mosquito-borne diseases. Curr. Probl. Pediatr. Adolesc. Health Care 39, 97–140 (2009).
Higgs, S. & Beaty, B. J. Natural cycles of vector-borne pathogens. Biology of disease vectors, In Biology of disease vectors 2nd edn (eds Marquardt, W. C. et al.) Elsevier Academic Press (New York, 2005).
Manguin, S. & Boëte C. Global impact of mosquito biodiversity, human vector-borne diseases and environmental change, In The importance of biological interactions in the study of biodiversity (ed. Lopez-Pujol, J. ) 27–50 (Intech open access Publisher, Rijeka, Croatia, 2011).
Beerntsen, B. T., James, A. A. & Christensen, B. M. Genetics of mosquito vector competence. Microbiol. Mol. Biol. Rev. 64, 115–137 (2000).
Kilpatrick, A. M. et al. West Nile virus risk assessment and the bridge vector paradigm. Emerg. Infect. Dis. 11, 425–429 (2005).
Roche, B., Rohani, P., Dobson, A. P. & Guégan, J.-F. The impact of community organization on vector-borne pathogens. Am. Nat. 181, 1–11 (2013).
Norris, D. E. Mosquito-borne diseases as a consequence of land use change. EcoHealth 1, 19–24 (2004).
Johnson, M. F., Gomez, A. & Pinedo-Vasquez, M. Land use and mosquito diversity in the Peruvian Amazon. J. Med. Entomol. 45, 1023–1030 (2008).
Steiger, D. M. et al. Effects of landscape disturbance on mosquito community composition in tropical Australia. J. Vector Ecol. 37, 69–76 (2012).
Becker, N. D. et al. Mosquitoes and their control 2nd edn, Springer Verlag (Berlin Heidelberg, Germany 2010).
Frankie, G. W. & Ehler, L. E. Ecology of insects in urban environments. Annu. Rev. Entomol. 23, 367–387 (1978).
McKinney, M. L. Effects of urbanization on species richness: a review of plants and animals. Urban Ecosyst. 11, 161–176 (2008).
McKinney, M. L. Urbanization, biodiversity, and conservation. Bioscience 52, 883–890 (2002).
Woolhouse, M. E. J., Taylor, L. E. & Haydon, D. T. Population biology of multihost pathogens. Science 292, 1109–1112 (2001).
Shochat, E., Warren, P. S., Faeth, S. H., McIntyre, N. E. & Hope, D. From patterns to emerging processes in mechanistic urban ecology. Trends Ecol. Evol. 21, 186–191 (2006).
Faeth, S. H., Warren, P. S., Shochat, E. & Marussich, W. A. Trophic dynamics in urban communities. BioScience 55, 399–407 (2005).
Gilioli, G. & Mariani, L. Sensitivity of Anopheles gambiae population dynamics to meteo-hydrological variability: a mechanistic approach. Malaria J. 10, 294 (2011).
Li, Y. et al. Urbanization increases Aedes albopictus larval habitats and accelerates mosquito development and survivorship. PLoS Negl. Trop. Dis. 8, e3301 (2014).
LaDeau, S. L., Allan, B. F., Leisnham, P. T. & Levy, M. Z. The ecological foundations of transmission potential and vector-borne disease in urban landscapes. Func. Ecol. 29, 889–901 (2015).
Chuang, T. W., Hockett, C. W., Kightlinger, L. & Wimberly, M. C. Landscape-level spatial patterns of West Nile virus risk in the northern Great Plains. Am. J. Trop. Med. Hyg. 86, 724–731 (2012).
Versteirt, V. Nationwide inventory of mosquito biodiversity (Diptera: Culicidae) in Belgium, Europe. Bull. Entomol. Res. 103, 193–203 (2013).
Ibañez-Justicia, A., Stroo, A., Dik, M., Beeuwkes, J. & Scholte, E. J. National mosquito (Diptera: Culicidae) survey in The Netherlands 2010–2013. J. Med. Entomol. 52, 185–198 (2015).
Overgaard, H. J. et al. Effect of landscape structure on anopheline mosquito density and diversity in northern Thailand: implication on malaria transmission and control. Landscape Ecol. 18, 605–619 (2003).
Byrne, K. & Nichols, R. A. Culex pipiens in London underground tunnels: differentiation between surface and subterranean population. Heredity 82, 7–15 (1999).
Kay, B. H. et al. The importance of subterranean mosquito habitat to arbovirus vector control strategies in North Queensland, Australia. J. Med. Entomol. 37, 846–853 (2000).
Muñoz, J. et al. Feeding patterns of potential West Nile virus vectors in South-West Spain. PLoS One 7, e39549 (2012).
Morchón, R., Carretón, E., González-Miguel, J. & Mellado-Hernández, I. Heartworm disease (Dirofilaria immitis) and their vectors in Europe – new distribution trends. Front. Physiol. 3, 196 (2012).
Figuerola, J., Jiménez-Clavero, M. A., Rojo, G., Gómez-Tejedor, C. & Soriguer R. Prevalence of West Nile Virus neutralizing antibodies in colonial aquatic birds in southern Spain. Avian Pathol. 36, 209–212 (2007).
Vázquez, A. et al. West Nile and Usutu viruses in mosquitoes in Spain, 2008–2009. Am J. Trop. Med. Hyg. 85, 178–181 (2011).
Ferraguti, M. et al. Avian Plasmodium in Culex and Ochlerotatus mosquitoes from Southern Spain: effects of season and host-feeding source on parasite dynamics. PLoS One 8, e66237 (2013).
Burkett-Cadena, N. D., McClure, C. J., Estep, L. K. & Eubanks, M. D. Hosts or habitats: What drives the spatial distribution of mosquitoes? Ecosphere 4, art30 (2013).
Hay, S. I., Packer, M. J. & Rogers, D. J. The impact of remote sensing on the study and control of invertebrate intermediate hosts and vectors for disease. Int. J. Remote Sens. 18, 2899–2930 (1997).
Johnston, E. et al. Mosquito communities with trap height and urban-rural gradient in Adelaide, South Australia: implications for disease vector surveillance. J. Vector Ecol. 39, 48–55 (2014).
Cox, J., Grillet, M. E., Ramos, O. M., Amador, M. & Barrera, R. Habitat segregation of dengue vectors along an urban environmental gradient. Am. J. Trop. Med. Hyg. 76, 820–826 (2007).
Leisnham, P. T. & Sandoval-Mohapatra, S. Mosquitoes associated with ditch-plugged and control tidal salt marshes on the Delmarva Peninsula. Int. J. Environ. Res. Public Health 8, 3099–3113 (2011).
Ciota, A. T. et al. Dispersal of Culex mosquitoes (Diptera: Culicidae) from a wastewater treatment facility. J. Med. Entomol. 49, 35–42 (2012).
Bogojević, M. S., Merdić, E. & Bogdanović, T. The flight distances of floodwater mosquitoes (Aedes vexans, Ochlerotatus sticticus and Ochlerotatus caspius) in Osijek, Eastern Croatia. Biologia 66, 678–683 (2011).
Padmanabha, H., Durham, D., Correa, F., Diuk-Wasser, M. & Galvani, A. The interactive roles of Aedes aegypti super-production and human density in Dengue transmission. PLoS Negl. Trop. Dis. 6, e1799 (2012).
Roche, B. et al. The spread of Aedes albopictus in metropolitan France: contribution of environmental drivers and human activities and predictions for a near future. PLoS One 10, e012560 (2015).
Collantes, F. et al. Review of ten-years presence of Aedes albopictus in Spain 2004–2014: known distribution and public health concerns. Parasite. Vector. 8, 655 (2015).
Reisen, W. K., Meyer, R. P., Tempelis, C. H. & Spoehel, J. J. Mosquito abundance and bionomics in residential communities in Orange and Los Angeles counties, California. J. Med. Entomol. 27, 356–367 (1990).
Roiz, D., Ruiz, S., Soriguer, R. & Figuerola, J. Landscape effects on the presence, abundance and diversity of mosquitoes in Mediterranean wetlands. PLoS One 10, e0128112 (2015).
Lopes, P. et al. Modelling patterns of mosquito density based on remote sensing images. Estoril Congress Center (2005).
Diuk-Wasser, M. A., Brown, H. E., Andreadis, T. G. & Fish, D. Modeling the spatial distribution of mosquito vectors for West Nile virus in Connecticut, USA. Vector-Borne & Zoonotic Diseases, 6, 283–295 (2006).
Santa-Ana, M., Khadem, M. & Capela, R. Natural infection of Culex theileri (Diptera: Culicidae) with Dirofilaria immitis (Nematoda: Filarioidea) on Madeira Island, Portugal. J. Med. Entomol. 43, 104–106 (2006).
Bueno Marí, R. & Jiménez Peydró, R. Malaria en España: aspectos entomológicos y perspectivas de futuro. Rev. Esp. Salud Publica 82, 467–479 (2008).
Toral, G. M., Aragones, D., Bustamante, J. & Figuerola, J. Using Landsat images to map habitat availability for waterbirds in rice fields. Ibis 153, 684–694 (2011).
Ponçon, N. et al. Effects of local anthropogenic changes on potential malaria vector Anopheles hyrcanus and West Nile virus vector Culex modestus, Camargue, France. Emerg. Infect. Dis. 13, 1810 (2007).
Vázquez, A. et al. Novel flaviviruses detected in different species of mosquitoes in Spain. Vector-Borne Zoonotic Dis. 12, 223–229 (2012).
Bisanzio, D. et al. Spatio-temporal patterns of distribution of West Nile virus vectors in eastern Piedmont Region, Italy. Parasite. Vector. 4, 230 (2011).
Farajollahi, A., Fonseca, D. M., Kramer, L. D. & Kilpatrick, A. M. “Bird biting” mosquitoes and human disease: a review of the role of Culex pipiens complex mosquitoes in epidemiology. Infect. Genet. Evol. 11, 1577–1585 (2011).
Rizzoli, A. et al. Understanding West Nile virus ecology in Europe: Culex pipiens host feeding preference in a hotspot of virus emergence. Parasite. Vector. 8, 213 (2015).
Vinogradova, E. B. Culex pipiens pipiens mosquitoes: taxonomy, distribution, ecology, physiology, genetics, applied importance and control, Vol. 2 (Pensoft Publishers, 2000).
Roiz, D. et al. Efficacy of mosquito traps for collecting potential West Nile mosquito vectors in a natural Mediterranean wetland. Am. J. Trop. Med. Hyg. 86, 642–648 (2012).
Schaffner, F. et al. The mosquitoes of Europe, an identification and training programme. CD-Rom. Montpellier, France (IRD Editions 2001).
Harbach, R. E. The identity of Culex perexiguus Theobald versus ex. univittatus Theobald in southern Europe. Eur. Mosq. Bull. 4, 7 (1999).
Balenghien, T. et al. Evidence of the laboratory vector competence of Culex modestus for West Nile virus. J. Am. Mosq. Control Assoc. 23, 233–236 (2007).
Aitchison, J. The statistical analysis of compositional data (ed. Chapman & Hall) 1–416 (London, UK, 1986).
Breiman, L. Random forests. Mach. Learn. 45, 5–32 (2001).
Cutler, D. R. et al. Random forests for classification in ecology. Ecology 88, 2783–2792 (2007).
Acknowledgements
This study was funded by project P11-RNM-7038 from the Junta de Andalucía, and project CGL2012-30759 from the Spanish Ministry of Science and Innovation and European Regional Development Fund (FEDER). Funds were also partially provided by EU grant HEALTH.2010.2.3.3-3 project 261391 EuroWestNile and by RNM118 and RNM153 from Junta de Andalucía. Information on remote sensing variables was obtained with the financial support of the project CGL2006-02247/BOS. M.F. and J.M.P. were supported by a FPU grant and a Juan de la Cierva contract, respectively. Laboratorio de SIG y Teledetección, Estación Biológica de Doñana, CSIC (LAST-EBD) provided logistical support. Cristina Pérez, Esmeralda Pérez, Juana Moreno Fernandez and Antonio Magallanes Martín de Oliva helped with insect capture and identification. Comments by two anonymous reviewers improved a previous version of the manuscript.
Author information
Authors and Affiliations
Contributions
M.F., J.M.-d.l.P., R.S. and J.F. conceived and designed the experiments; M.F., J.M.-d.l.P., D.R. and S.R. collected samples and performed the experiments; M.F., J.M.-d.l.P. and J.F. carried out the data analysis; J.F. and R.S. contributed to the reagents/materials/analysis tools. M.F., J.M.-d.l.P., D.R., S.R., R.S. and J.F. wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Ferraguti, M., Martínez-de la Puente, J., Roiz, D. et al. Effects of landscape anthropization on mosquito community composition and abundance. Sci Rep 6, 29002 (2016). https://doi.org/10.1038/srep29002
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep29002
This article is cited by
-
Distribution of Culex pipiens life stages across urban green and grey spaces in Leiden, The Netherlands
Parasites & Vectors (2024)
-
Environmental and geographical factors influence the occurrence and abundance of the southern house mosquito, Culex quinquefasciatus, in Hawai‘i
Scientific Reports (2024)
-
Aedes albopictus in a recently invaded area in Spain: effects of trap type, locality, and season on mosquito captures
Scientific Reports (2024)
-
Observations and modelling of mosquito prevalence within urban areas – A case study from Uppsala, Sweden
Urban Ecosystems (2024)
-
Modeling the effects of species associations and abiotic parameters on the abundance of mosquito species in a Mediterranean wetland
Wetlands Ecology and Management (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
