
Abstract
TMPyP4 is widely considered as a potential photosensitizer in photodynamic therapy and a G-quadruplex stabilizer for telomerase-based cancer therapeutics. However, its biological effects including a possible adverse-effect are poorly understood. In this study, whole genome RNA-seq analysis was used to explore the alteration in gene expression induced by TMPyP4. Unexpectedly, we find that 27.67% of changed genes were functionally related to cell adhesion. Experimental evidences from cell adhesion assay, scratch-wound and transwell assay indicate that TMPyP4 at conventional doses (≤0.5 μM) increases cell-matrix adhesion and promotes the migration of tumor cells. In contrast, a high dose of TMPyP4 (≥2 μM) inhibits cell proliferation and induces cell death. The unintended “side-effect” of TMPyP4 on promoting cell migration suggests that a relative high dose of TMPyP4 is preferred for therapeutic purpose. These findings contribute to better understanding of biological effects induced by TMPyP4 and provide a new insight into the complexity and implication for TMPyP4 based cancer therapy.
Similar content being viewed by others


Introduction
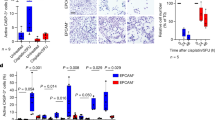
Photodynamic therapy (PDT) induces cancer cell death (necrosis or apoptosis) mainly by reactive oxygen species (ROS), which are produced by irradiated photosensitizers1. Compared to the conventional anticancer therapy, PDT is less invasive with better tolerance and outcome. In addition, PDT has obvious advantages over other cancer therapeutics such as surgery, radiation and chemotherapy: a minimal functional disturbance, being repetitively applicable on the same site and a low recurrence2. PDT has been rapidly developed over past decades with a great potential to treat multiple types of cancers including esophageal cancer and non-small cell lung cancer3,4. The photosensitizer is crucial for PDT treatment5. However, it has been challenging to obtain an optimal photosensitizer with a high yield of singlet oxygen (1O2) and high precision targeting cancer cells5. TMPyP4 (Fig. 1A), a porphyrins derivative, has been considered as a promising photosensitizer due to its high water solubility, high permeability through cell membrane and preferential accumulation in tumor cells6,7,8.

(A) The molecule of TMPyP4 used in this study. Counter ions are p-toluenesulfonate. Statistics of RNA-seq data comparing gene expression in TMPyP4 treated and untreated A549 cells. “GO” analysis showed the functional group of genes changed in expression. (B) As in (A) except that TPyP4-Pt was used to treat cells. Counter ions are NO3− anions.
Besides potentially serving as a photosensitizer in PDT, TMPyP4 has been recently developed as a chemotherapeutics drug to inhibit telomerase activity in cancer cells9,10,11. About 85% of cancer cells overcome the proliferative limit by activating telomerase, a ribonucleoprotein with reverse transcriptase activity that adds telomeric DNA repeats to the 3′-overhang of telomeres, thus maintaining telomere length and chromosome integrality12. Accumulated evidences show that single-stranded 3′-overhang of telomeres can stack via Hoogsteen hydrogen bonding into a structure referred as G-quadruplex13. TMPyP4 is able to associate and stabilize G-quadruplex, thereby blocking telomerase action. TMPyP4 treatment leads to progressive telomere shortening that eventually results in cancer cell death by apoptosis or senescence14.
Because DNA sequence with a potential to form G-quadruplex is widely present on genome, it has been reported that TMPyP4 treatment may lead to multiple consequences including the alteration of expression of particular genes15,16,17,18,19,20 and/or the interference with DNA replication21,22. Therefore, it is important to comprehensively understand biological effects induced by TMPyP4 before it can be used for anti-cancer therapeutics. Moreover, a possible adverse effect is worth investigating.
In this report, human A549 cancer cells were treated with TMPyP4 or its derivative TPyP4-Pt (Fig. 1B), and gene expression profile for treated and untreated cells was obtained by RNA-seq. Unexpectedly, we found that among the genes changed by TMPyP4 or TPyP4-Pt, ~27% are involved in cell adhesion and migration, implying that TMPyP4 treatment might affect cancer metastasis. The experiments including cell adhesion assay, scratch-wound healing assay and transwell assay demonstrate that TMPyP4 at commonly used dose (≤0.5 μM, close to its light IC50 values) promotes cancer cell migration. In strikingly contrast, the high-dose of TMPyP4 ( ) inhibits cell proliferation and induces cell death. These findings provide new insights into the complexity of TMPyP4 as a possible anticancer drug.
) inhibits cell proliferation and induces cell death. These findings provide new insights into the complexity of TMPyP4 as a possible anticancer drug.
Results
TMPyP4 changes the expression of adhesion-related genes in human lung cancer cells A549
The effect of TMPyP4 on global gene expression in cancer cells was evaluated using RNA-seq, a whole transcriptome sequencing (mRNA, Hiseq2000-PE125). Human A549 lung cancer cells were cultured in the presence or absence of 0.5 μM TMPyP4 for 2 days; their mRNA was isolated and subjected to RNA-seq. The top 100 changed mRNA transcripts and their abundance are listed in Table S1 and full sequence data from these experiments were uploaded to GEO database under accession number of GSE72983. Changed genes were functionally grouped by GO-biology analysis. Our results showed that the expression of 1.73% genes was changed upon TMPyP4 treatment. 27.67% genes of them were functionally related to cell adhesion and migration (Fig. 1A, Table S4). To further verify these data, we treated A549 cells with TPyP4-Pt (Fig. 1B), a derivative of TMPyP4 with similar characteristics to TMPyP415. Consistent with the results from TMPyP4, TPyP4-Pt treated cells displayed the increasement of cell adhesion and migration related genes (28.36%) (Fig. 1B, Table S2 and Table S5).
TMPyP4 increases cell-matrix adhesion
The commonly used concentration for TMPyP4 study is near its light IC50 values, i.e. 0.25 μM in vitro23. RNA-seq results implied that cell adhesion to extracellular matrix may be changed upon TMPyP4 or TPyP4-Pt treatment. To test this, cell adhesion assay that determines the adhesion between cell and attached matrix was performed in multiple cancer cell lines including human A549, HeLa, osteosarcoma U2OS and SAOS2. As shown in Fig. 2, all four cell lines displayed significant increase in cell adhesion to matrix after treatment with TMPyP4 (0.125 μM, 0.25 μM and 0.5 μM). Similar phenomena were observed when cells were treated with TPyP4-Pt (Figure S1). These results suggested that TMPyP4 and its derivate increases cell-matrix adhesion by changing the expression of adhesion-related genes. To validate this, MUC5B, a top hit of RNA-seq that has been previously reported to promote cell adhesion24, was knocked down by siRNA in A549 cells (Fig. 2E). Indeed, the knockdown of MUC5B resulted in the decrease of cell attachment (Fig. 2F). However, TMPyP4 failed in promoting the attachment of MUC5B deficient cells (Fig. 2F), demonstrating that the change of gene expression such as MUC5B is responsible for increased cell adhesion upon TMPyP4 treatment.

(A–D) TMPyP4 increase cell adhesion to extracellular matrix. Indicated cell lines were used. (E) qRT-PCR showed the abundance of MUC5B mRNA in A549 cancer cell line (NC) and A549 cells transfected with siRNA targeting MUC5B (siRNA_1 and siRNA_2). (F) MUC5B knock-down suppress the adhesion of A549 cancer cells to extracellular matrix. MUC5B deficient cells showed no increase of cell adhesion upon TMPyP4 treatment. Values are average ± SD of three independent experiments.
TMPyP4 promotes cancer cell migration
Cell adhesion is often associated with cell migration25. To this end, we performed scratch-wound healing assay to determine the migration rate of A549 and U2OS cells in the presence or absence of TMPyP4. Our results showed that TMPyP4 significantly increased cancer cell migration (Fig. 3). After 72h treatment, 51% scratch was covered by migrated A549 cells, while 90% scratch was covered when cells were treated with 0.25 μM TMPyP4 (Fig. 3A,B). Similarly, 0.25 μM TMPyP4 treated U2OS cells showed significantly increased migration rate (Fig. 3C,D). TPyP4-Pt treated A549 cells also displayed increased cell migration compared to untreated cells (Figure S2). These results demonstrated that TMPyP4 and its derivate enhance the migration of cancer cells.

(A) TMPyP4 promotes A549 cells migration in scratch-wound healing assay. (B) Quantification of (A). (C) as in (A) except U2OS cells were used. (D) Quantification of (C). (E) TMPyP4 promoted A549 cells migration in a transwell assay. (F) as in (E) except U2OS cells were used. Values are average ± SD of three independent experiments.
To further confirm the strengthening migration ability of cancer cells induced by TMPyP4, transwell migration assay was performed, in which cells are initially placed on the upper layer of permeable cell membrane and the cells that have migrated through the membrane are stained and counted after certain period of times. The quantitative data demonstrated that A549 and U2OS cells treated with TMPyP4 exhibited increased transferability compared to untreated cells (Fig. 3E,F).
TMPyP4 at high dose induce cancer cell death
While TMPyP4 at commonly used concentration (0.25 or 0.5 μM) may have undesired “side-effect” on promoting cancer cell migration, we then explored a possible strategy to eliminate this effect. It has come to our attention that A549 cells treated with higher concentration of TMPyP4 (2 μM) exhibited remarkably slower proliferation compared with control cells or cells treated with lower dose of TMPyP4 (Fig. 4A). This phenomenon was also observed in HeLa, U2OS and SAOS2 cells (Figure S3). FACS analysis indicated that 2.0 μM TMPyP4 induced apoptosis of ~23% A549 cells after only 3 days (Fig. 4B,C). Likewise, 33% U2OS cells underwent apoptosis after 3 days treatment with 2.0 μM of TMPyP4 (Figure S4). Moreover, scratch-wound healing assay demonstrated that the treatment with 2.0 μM TMPyP4 slowed down cell migration rate (Fig. 4D,E and Figure S5).

(A) Proliferation curve of A549 cells in the presence of low and high dose of TMPyP4. (B) 1.0 or 2.0 μM TMPyP4 treatment induces apoptosis of A549 cells. Apoptotic cells were assayed by Annexin V/PI staining and FACS analysis. (C) Quantification of (B). (D) Scratch-wound healing assay showing decreased migration rate of A549 cells when treated with high-dose TMPyP4 (2.0 μM). (E) Quantification of (D). (F) Statistics of RNA-seq data comparing gene expression in 2.0 μM TMPyP4 treated and untreated A549 cells. Values are average ± SD of three independent experiments.
As expected, TMPyP4 inhibited telomerase activity of A549 (Figure S6), however, it is unlikely that rapid cell apoptosis (in 3 days) induced by TMPyP4 is due to the inhibition of telomerase. We thus performed RNA-seq analysis to determine the change of gene expression in response to high dose of TMPyP4 (2 μM). The results showed that expression of 2.11% genes was changed upon TMPyP4 (2 μM) treatment (Fig. 4F and Table S3), of which 26.52% genes were functionally related to cell proliferation and apoptosis, and 12.55% genes were related to cell adhesion and migration (Fig. 4F, Table S3 and Table S6). Strikingly, a substantial number of cell adhesion and migration related genes are found in both 0.5 and 2 μM treated cells, indicating the same mechanism underlying gene regulation by TMPyP4.
Discussion
TMPyP4 treatment changes the expression of a significant number of genes (1.73% of whole genome). TMPyP4 and its derivate may affect gene transcription in multiple manners. First, TMPyP4 is able to incorporate into duplex DNA26, thus may facilitate or block gene transcription; second, given the fact that G-rich sequences that can fold into G-quadruplex are widely present on genome27 and highly enriched in promoter region of genes28, it is conceivable that G-quadruplex stabilized by TMPyP4 interferes the transcription of those genes; third, TMPyP4 may increase the level of ROS in cells (even cells are cultured in dark, the light can’t be absolutely excluded), thereby resulting in a global cell response including the change in expression of particular genes. Intriguing finding in this study is that among all genes affected by TMPyP4 (~1.7% of whole genome), ~27% of them are involved in cell adhesion and migration (Fig. 1A). While molecular mechanisms underlying the regulation of these genes are remained to be elucidated, the consequence is that TMPyP4 treated cells exhibit increased ability to adhere to extracellular matrix (Fig. 2). Accordingly, increased migration rate (Fig. 3A–D) and transferability (Fig. 3E,F) was observed for TMPyP4 treated cells. It has been widely accepted that increased cell migration and transferability are closely associated with cancer invasion and metastasis, a major cause of patient death for variety of cancers29. Thus, our study implies that TMPyP4 may increase the risk of cancer metastasis. This raises a serious question regarding how to prevent this “side-effect” of TMPyP4 in cancer therapeutics.
TMPyP4 is first developed as a photosensitizer in PDT6,7,8. The treatment process of PDT involves the injection of photosensitizer to patient’s blood7, followed by irradiation of diseased tissue (tumors) with a high intensity light7. However, a number of tumors may escape from irradiation during treatment of metastatic cancers due to the fact that PDT cannot irradiate all the tumors at a time30,31. In addition, the light used in PDT cannot penetrate more than 1cm tumor tissue30,31. In these scenarios, escaped cancer cells may become more metastatic aggressive. Our results thus provide a strong rational to pay a great attention to the “side-effect” of TMPyP4 in PDT.
TMPyP4 has also been proposed to serve as a potential G-quadruplex stabilizer to inhibit telomerase-mediated telomere lengthening9,10,32. Because telomerase is exclusively present in cancer and stem cells, the inhibition of telomerase action has been considered as a promising approach for specifically targeting cancer cells. As telomeres shorten at rate of ~60 bp/PD in absence of telomerase, it takes a long time before cancer cells undergo crisis due to extremely short telomeres33. In this context, TMPyP4 may not be a good candidate for telomerase-based therapeutics because it increases the risk of metastasis.
The treatment with a high dose of TMPyP4 (≥2 μM) results in cell apoptosis, which may be responsible for slower cell proliferation and decreased migration rate (Fig. 4). High doses may be toxic to normal cells in body; it nevertheless provides a possible option for application of TMPyP4 in oncotheropeutics. Alternatively, TMPyP4 needs to be used in combination with other therapeutic drug to minimize its side-effect. Altogether, our study reveals dose-dependent effect of TMPyP4 in cancer treatment and provides a strong rational to re-consider the strategy for application of TMPyP4 in cancer therapeutics.
Methods
Cell Culture
A549, HeLa, U2OS and SAOS2 were obtained from Cell Resource Center of Peking Union Medical College and cultured at 37 °C under 5% CO2 in DMEM (gibco) supplemented with 10% fetal calf serum (gibco) and 100 U/mL penicillin and streptomycin (HyClone). TMPyP4 was purchased from Sigma.
RNA-seq whole transcriptome sequencing
8.0 × 105 A549 cells were seeded to 10 cm2 dish, after incubating for 6 h, the TMPyP4 or TPyP4-Pt was added to medium (a final concentration of 0.5 μM). After incubating for 2 days, cells were harvested and total RNA were isolated by Trizol (Takara) extraction. 3 μg RNA per sample was used as input material for the RNA sample preparations. Sequencing libraries were generated using a NEBNext Ultra RNA Library Prep Kit for Illumina (NEB, USA) according to the manufacturer’s recommendations and sequencing was performed on an Illumina Hiseq 2000 platform. Raw data were extracted by Basecalls of CASAVA (version 1.8). Sequenced reads were mapped to whole genome (hg19) using Tophat2.0.2. Unique reads were used for further analysis. The abundance of reads for genes was determined by htseq-count script (Version 0.6.1) with parameters -s no. The expression level of genes was normalized by Fragment Per Kilobase of exon per Megabase of library size (FPKM)(50). The correlation between different samples was measured by Spearman’s rank correlation test and visualized with R package.
Long-term cell proliferation
Long-term cell proliferation experiments were carried out using A549, HeLa, U2OS and SAOS2 cancer cell lines. 5.0 × 105 cells were seeded to 10 cm2 dish and incubated for 6 h, TMPyP4 or TPyP4-Pt was added to medium to a final concentration of 0.125, 0.25, 0.5, 1.0 or 2.0 μM, respectively. The medium was changed every three days until confluence was reached.
Annexin V/PI apoptosis assay
Cells were seeded in 10 cm2 dish at a density of 5.0 × 105cells per dish and incubated at 37 °C for ~6 h until cells attached to the dish. TMPyP4 with final concentration of 1.0 or 2.0 μM were then added into medium. After 3 days, cells were harvested for anexin V/PI apoptosis assay. The assay was performed following the protocol provided by the Annexin V/PI apoptosis Kit (Sigma).
Cell adhesion assay
The 96-well plate was coated with 2.5 μg/ml human fibronectin in PBS (Millipore, CA) for 2 h at room temperature. Cells were seeded into the 96-well plate at a density of 4 × 104cells/well and cultured for 1h at 37 °C in a CO2 incubator. Cells were then rinsed three times with 10% formalin and stained with crystal violet for 5 min at room temperature. After three times washing with ddH2O, stained cells were dissolved in 100 μL 33% acetic acid. The absorbance at 560 nm was detected by Synergy H1 Multi-Mode Reader (BioTek). Relative number of cells attaching to extracellular matrix was calculated using the following equation: mean OD of treated cells/mean OD of control cells. Cells treated with vehicle (0.1% DMSO) were used as a control.
Transwell assay
Transwell assay was performed using Transwell Kit (Corning Costar, NY) following manufacturer’s instructions. Briefly, cells were pretreated with different concentrations (0, 0.125 or 0.25 μM) of TMPyP4 for 3 days, trypsinized and seeded into Transwell Permeable Support (insert) pre-equilibrated with serum-free DMEM medium. For each group, 1 × 105cells/insert were seeded and incubated in 100 μl serum-free DMEM medium. The insert was placed on 24-well plate containing 600 μl of DMEM medium with 10% FBS. After 24 h of culturing, cells on the upper surface of insert were removed with cotton-tipped swabs. And the cells on backside surface of insert were fixed with 10% formalin, stained with crystal violet for 5 min at room temperature, and washed three times with ddH2O. Stained cells were dissolved in 500 μL 33% acetic acid and their absorbance was detected at 560 nm by Synergy H1 Multi-Mode Reader (BioTek).
The scratch-wound assay
The cells were seeded in a 6-well plate at a density of 2 × 105cells/well and cultured in medium containing TMPyP4 or TPyP4-Pt for 3 days. A denuded area was created across the diameter of dish by a yellow tip. The cells were washed with PBS and incubated in a serum free medium. Phase-contrast images were taken at a time point of 0, 24, 48 and 72 h of incubation. Images were analyzed with Axiovision Rel.4.8 software. The percentage of areas covered by migrated cells (wound recovery) was calculated. Three independent experiments were carried out for quantification.
Knockdown of MUC5B by siRNA
MUC5B or PCDH17 was knocked down in A549 cells by siRNA. siRNA sequence are: siRNA_1-GCAGCTACGTTCTGTCCAA; siRNA_2 GCGTGTTCCTCAACTCCAT; siRNA were transfected into cells using Lipofectamine RNAiMAX (Life Technologies) following manufacturer’s instruments. SiRNA with scramble sequence was used as a negative control (NC). Cells were assayed 60 h after transfection.
Quantitative real-time PCR (qRT-PCR)
Total RNA of A549 cells treated with indicated siRNA was extracted by RNAiso Plus Reagent (Takara). cDNA was produced using Superscript III Reverse Transcriptase (Invitrogen) and random primers. qRT-PCR were performed using Fast SYBR Green PCR mastermix (Invitrogen) and specific primers (MUC5B: 5′-GCCTACGAGGACTTCAACGTC-3′, 5′-CCTTGATGACAACACGGGTGA-3′; β-actin: 5′-CATGTACGTTGCTATCCAGGC-3′, 5′-CTCCTTAATGTCACGCACGAT-3′). The relative abundance of MUC5B mRNA were calculated using the formula of 2−ΔΔCT and normalized to an endogenous housekeeping gene (β-actin). Values are means 6 standard deviation from four to six independent samples.
TRAP assay
A telomerase extract (1.0 μl, 200 cells) was prepared from A549 cells with NP-40 lysis buffer. Each reaction was performed in a final volume of 20 μl in a reaction mixture containing of 2.0 μL of 10× TRAP buffer, 1.6 μL of dNTP mix, 0.4 μL of TS primer (100 ng/mL), 0.8 μL Primer mix (100 ng/mL), 2.0 μL TSNT internal control primer (4.0 × 10−11M), 0.4 μL of RNase inhibitor (2 U/mL), 0.16 μL of Taq polymerase (5 U/mL), 5.0 μL of different concentrations of TMPyP4 and 6.64 μL of DEPC-treated water. The experiment was performed as previously reported34,35. Relative telomerase activity was calculated as a ratio of signal in TRAP ladder to signal of an internal control.
Statistical analysis
The student’s 2-tailed unpaired t-test was used to determine statistical significance and the resulting P-values are indicated in figures (*P < 0.05; **P < 0.01; ***P < 0.001).
Additional Information
How to cite this article: Zheng, X.-H. et al. TMPyP4 promotes cancer cell migration at low doses, but induces cell death at high doses. Sci. Rep. 6, 26592; doi: 10.1038/srep26592 (2016).
References
Pervaiz, S. & Olivo, M. Art and science of photodynamic therapy. Clin. Exp. Pharmacol. Physiol. 33, 551–556 (2006).
Wilson, B. C. & Patterson, M. S. The physics, biophysics and technology of photodynamic therapy. Phys. Med. Biol. 53, R61–109 (2008).
Dolmans, D. E., Fukumura, D. & Jain, R. K. Photodynamic therapy for cancer. Nat. Rev. Cancer 3, 380–387 (2003).
Dabrowski, J. M. & Arnaut, L. G. Photodynamic therapy (PDT) of cancer: from local to systemic treatment. Photochem. Photobiol. Sci. 14, 1765–1780 (2015).
Benov, L. Photodynamic therapy: current status and future directions. Med. Princ. Pract. 24 Suppl 1, 14–28 (2015).
Villanueva, A., Caggiari, L., Jori, G. & Milanesi, C. Morphological aspects of an experimental tumour photosensitized with a meso-substituted cationic porphyrin. J. Photochem. Photobiol. B 23, 49–56 (1994).
Awan, M. A. & Tarin, S. A. Review of photodynamic therapy. Surgeon 4, 231–236 (2006).
Tada-Oikawa, S., Oikawa, S., Hirayama, J., Hirakawa, K. & Kawanishi, S. DNA damage and apoptosis induced by photosensitization of 5,10,15,20-tetrakis (N-methyl-4-pyridyl)-21H,23H-porphyrin via singlet oxygen generation. Photochem. Photobiol. 85, 1391–1399 (2009).
Rha, S. Y. et al. Effect of telomere and telomerase interactive agents on human tumor and normal cell lines. Clin. Cancer Res. 6, 987–993 (2000).
Shammas, M. A. et al. Telomerase inhibition and cell growth arrest by G-quadruplex interactive agent in multiple myeloma. Mol. Cancer Ther. 2, 825–833 (2003).
Grand, C. L. et al. The cationic porphyrin TMPyP4 down-regulates c-MYC and human telomerase reverse transcriptase expression and inhibits tumor growth in vivo . Mol. Cancer Ther. 1, 565–573 (2002).
Morin, G. B. The human telomere terminal transferase enzyme is a ribonucleoprotein that synthesizes TTAGGG repeats. Cell 59, 521–529 (1989).
Gellert, M., Lipsett, M. N. & Davies, D. R. Helix formation by guanylic acid. PNAS 48, 2013–2018 (1962).
Fujimori, J. et al. Antitumor Effects of Telomerase Inhibitor TMPyP4 in Osteosarcoma Cell Lines. J. Orthop. Res. 29, 1707–1711 (2011).
Zheng, X. H. et al. Platinum(II) clovers targeting G-quadruplexes and their anticancer activities. Dalton Trans. 44, 50–53 (2015).
Hurley, L. H., Von Hoff, D. D., Siddiqui-Jain, A. & Yang, D. Z. Drug targeting of the c-MYC promoter to repress gene expression via a G-quadruplex silencer element. Semin. Oncol. 33, 498–512 (2006).
Siddiqui-Jain, A., Grand, C. L., Bearss, D. J. & Hurley, L. H. Direct evidence for a G-quadruplex in a promoter region and its targeting with a small molecule to repress c-MYC transcription. PNAS 99, 11593–11598 (2002).
Bearss, D. J., Munoz, R. M., Han, H., Hurley, L. H. & Von Hoff, D. D. Specific inhibition of c-MYC expression by the cationic porphyrin TMPyP4 results in downregulation of hTERT expression and reduced telomerase activity. Clin. Cancer Res. 6, 4500s–4500s (2000).
Cogoi, S. & Xodo, L. E. G-quadruplex formation within the promoter of the KRAS proto-oncogene and its effect on transcription. Nucleic Acids Res. 34, 2536–2549 (2006).
Morris, M. J., Wingate, K. L., Silwal, J., Leeper, T. C. & Basu, S. The porphyrin TmPyP4 unfolds the extremely stable G-quadruplex in MT3-MMP mRNA and alleviates its repressive effect to enhance translation in eukaryotic cells. Nucleic Acids Res. 40, 4137–4145 (2012).
Vannier, J. B., Pavicic-Kaltenbrunner, V., Petalcorin, M. I., Ding, H. & Boulton, S. J. RTEL1 dismantles T loops and counteracts telomeric G4-DNA to maintain telomere integrity. Cell 149, 795–806 (2012).
Vannier, J. B. et al. RTEL1 is a replisome-associated helicase that promotes telomere and genome-wide replication. Science 342, 239–242 (2013).
Liu, H. L. et al. Antitumor activity of G-quadruplex-interactive agent TMPyP4 with photodynamic therapy in ovarian carcinoma cells. Oncol. Lett. 8, 409–413 (2014).
Valque, H., Gouyer, V., Gottrand, F. & Desseyn, J. L. MUC5B leads to aggressive behavior of breast cancer MCF7 cells. PLoS One 7, e46699 (2012).
Boyd, D. Invasion and metastasis. Cancer Metastasis Rev. 15, 77–89 (1996).
Ohyama, T., Mita, H. & Yamamoto, Y. Binding of 5,10,15,20-tetrakis(N-methylpyridinium-4-yl)-21H,23H-porphyrin to an AT-rich region of a duplex DNA. Biophys. Chem. 113, 53–59 (2005).
Todd, A. K. Bioinformatics approaches to quadruplex sequence location. Methods 43, 246–251 (2007).
Huppert, J. L. & Balasubramanian, S. G-quadruplexes in promoters throughout the human genome. Nucleic Acids Res. 35, 406–413 (2007).
Weiss, L. Metastatic inefficiency. Adv. Cancer Res. 54, 159–211 (1990).
Filip, A. G. et al. Photodynamic therapy--indications and limits in malignant tumors treatment. Rom. J. Intern. Med. 46, 285–293 (2008).
Capella, M. A. & Capella, L. S. A light in multidrug resistance: photodynamic treatment of multidrug-resistant tumors. J. Biomed. Sci. 10, 361–366 (2003).
Grand, C. L. et al. The cationic porphyrin TMPyP4 down-regulates c-MYC and human telomerase reverse transcriptase expression and inhibits tumor growth in vivo . Mol. Cancer Ther. 2, 568–573 (2003).
Zhao, Y. et al. Telomere extension occurs at most chromosome ends and is uncoupled from fill-in in human cancer cells. Cell 138, 463–75 (2009).
Kim, N. W. et al. Specific association of human telomerase activity with immortal cells and cancer. Science 266, 2011–5 (1994).
Zheng, X. H., Chen, H. Y., Tong, M. L., Ji, L. N. & Mao, Z. W. Platinum squares with high selectivity and affinity for human telomeric G-quadruplexes. Chem. Commun. 48, 7607–9 (2012).
Acknowledgements
TPyP4-Pt is a gift from Prof. Zong-Wan Mao (SYSU, Guangdong, China). This work was supported by National Natural Science Foundation of China Grants [31271472, 31322033, 81273275], China’s Post-doctoral Science Fund (2014M562208), the Science and Technology Innovation Commission of Shenzhen under Grants JCYJ20150525092941055 for financial support.
Author information
Authors and Affiliations
Contributions
X.-H.Z., L.-X.X. and Y.Z. conceived and designed the experiments. X.-H.Z. conducted RNA-seq whole transcriptome sequencing, long-term cell proliferation experiment, cell adhesion assay and the scratch-wound assay X.N. conducted annexin V/PI apoptosis assay. H.-Y.L. helped in the RNA-seq whole transcriptome sequencing and data analysis. Y.-M.F. peformed the long-term cell proliferation and cell adhesion assay. X.-H.Z., L.-X.X. and Y.Z. wrote themanuscript. All authors discussed the results and commented on the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Zheng, XH., Nie, X., Liu, HY. et al. TMPyP4 promotes cancer cell migration at low doses, but induces cell death at high doses. Sci Rep 6, 26592 (2016). https://doi.org/10.1038/srep26592
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep26592
This article is cited by
-
Development and application of metallo-phthalocyanines as potent G-quadruplex DNA binders and photosensitizers
JBIC Journal of Biological Inorganic Chemistry (2023)
-
Teloxantron inhibits the processivity of telomerase with preferential DNA damage on telomeres
Cell Death & Disease (2022)
-
Multiple G-quadruplex binding ligand induced transcriptomic map of cancer cell lines
Journal of Cell Communication and Signaling (2022)
-
Piperlongumine inhibits migration and proliferation of castration-resistant prostate cancer cells via triggering persistent DNA damage
BMC Complementary Medicine and Therapies (2021)
-
H2O2 enhances the anticancer activity of TMPyP4 by ROS-mediated mitochondrial dysfunction and DNA damage
Medical Oncology (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
