
Abstract
Lymphatic filariasis and onchocerciasis are parasitic helminth diseases, which cause severe morbidities such as elephantiasis, skin disease and blindness, presenting a major public health burden in endemic communities. The anti-Wolbachia consortium (A·WOL: http://www.a-wol.com/) has identified a number of registered antibiotics that target the endosymbiotic bacterium, Wolbachia, delivering macrofilaricidal activity. Here we use pharmacokinetics/pharmacodynamics (PK/PD) analysis to rationally develop an anti-Wolbachia chemotherapy by linking drug exposure to pharmacological effect. We compare the pharmacokinetics and anti-Wolbachia efficacy in a murine Brugia malayi model of minocycline versus doxycycline. Doxycycline exhibits superior PK in comparison to minocycline resulting in a 3-fold greater exposure in SCID mice. Monte-Carlo simulations confirmed that a bi-daily 25–40 mg/Kg regimen is bioequivalent to a clinically effective 100–200 mg/day dose for these tetracyclines. Pharmacodynamic studies showed that minocycline depletes Wolbachia more effectively than doxycycline (99.51% vs. 90.35%) after 28 day 25 mg/Kg bid regimens with a more potent block in microfilarial production. PK/PD analysis predicts that minocycline would be expected to be 1.7 fold more effective than doxycycline in man despite lower exposure in our infection models. Our findings warrant onward clinical investigations to examine the clinical efficacy of minocycline treatment regimens against lymphatic filariasis and onchocerciasis.
Similar content being viewed by others

Introduction
Lymphatic filariasis (LF) and onchocerciasis are parasitic helminth diseases affecting an estimated 158 million people globally, causing a substantial socioeconomic and health burden in endemic communities1,2,3. Lymphatic filariasis, caused by the filarial nematodes Wuchereria bancrofti, Brugia malayi and B. timori affects an estimated 120 million people. Morbidity associated with LF includes lymphoedema, elephantiasis and hydrocele1,3. Onchocerciasis is caused by Onchocerca volvulus and affects approximately 38 million people. Symptomatically, onchocerciasis presents as skin disease and can cause loss of sight in advanced untreated cases due to the immune response to migration and death of microfilariae in the eye2,3.
Current filariasis treatment and control programs rely on the use of the microfilaricides diethylcarbamazine, ivermectin and albendazole. In the context of mass drug administration (MDA), a combination of ivermectin and albendazole is used to control LF in Africa whereas a diethylcarbamazine/albendazole combination treatment is mainly used in the rest of the world3. In the case of onchocerciasis, ivermectin is administered as a monotherapy during MDA in all endemic areas. Targeting the microfilariae has proved effective in many cases, however, the length of treatment required (5 years for LF and up to 17 years or more for onchocerciasis) combined with long life span of an adult worm means that transmission will not be interrupted in some cases, especially where MDA is disrupted3,4. Treatment in geographical regions in which Loa loa is co-endemic is further complicated as ivermectin can cause severe adverse reaction events (SAEs), which can result in encephalophathy, coma and death5. These SAEs are probably caused by the drug-induced paralysis and death of the Loa loa microfilariae in the brain6. There is also increasing evidence of ivermectin resistance emerging in some endemic regions7. Epidemiological modelling also suggests MDA based on a microfilaricide mode of action may not be effective for elimination in all scenarios8,9, therefore alternative treatment strategies that include the use of macrofilaricidal (curative) drugs require development to facilitate the elimination of filarial diseases10.
Lymphatic filariae and O. volvulus are host to the obligate endosymbiotic bacterium, Wolbachia, which is essential for their development, growth and survival10. Wolbachia also contribute to filarial disease pathology11,12 and are associated with inflammatory adverse reactions to standard microfilaricidal treatments13,14. It has been shown that targeting Wolbachia with antibiotic drugs can induce growth retardation, embryotoxicity and death of adult filarial nematodes in preclinical models15,16 and in humans17. Doxycycline is the current gold standard anti-Wolbachia treatment with a growing body of clinical evidence that it has macrofilaricidal activity against LF18,19 and onchocerciasis20,21,22. Tetracyclines including doxycycline have been shown to suppress immune activity and therefore may offer the added benefit of alleviating symptomology23,24. This effect has been confirmed in clinical studies in which 200 mg/day doxycycline has been given for six weeks, showing reduced lymphangiogenic factors and filarial lymphedema25,26. Additionally, doxycycline is safe to use in Loa loa co-endemic areas as these parasites do not harbour Wolbachia22. The use of doxycycline in mass drug administration is however hampered by the duration of an optimally effective treatment (for onchocerciasis, recently defined as a minimum of 4 weeks, 100 mg, in a meta-analytical modelling framework) which could pose a hurdle to treatment adherence27.
The anti-Wolbachia consortium (A·WOL) program has identified a number of registered antibiotics that were effective against Wolbachia in in vitro and in vivo model systems28. The tetracycline, minocycline, caused a 1.6-fold log drop in Wolbachia in an in vitro C6/36 (wAlbB) cell based screen when administered at a concentration of 10 μM. A regimen of 25 mg/Kg/day of minocycline administered for 10 days elicited a log drop of 3.78 in Wolbachia ftsZ DNA in the Litomosoides sigmodontis murine filariasis model28. The data suggest superiority of minocycline over doxycycline. In this current work, we have investigated and compared the anti-Wolbachia efficacy of orally administered minocycline and doxycycline against the human filarial parasite B. malayi in a SCID mouse model29. Using quantitative assessment of Wolbachia load and PK/PD analysis we show that for a dosage that is equivalent to a previous clinically effective 100–200 mg/day doxycycline dose, minocycline is significantly more efficacious. The origin of these efficacy differences has been dissected through analysis of the relative contributions of drug pharmacokinetics and pharmacodynamics. Treatment duration and dosage have been investigated to identify optimal treatment regimens.
Results
In vitro anti-Wolbachia potency of minocycline vs doxycycline
The anti-Wolbachia potency of minocycline and doxycycline in in vitro Wolbachia infected A. albopictus cells C6/36 and in vitro male Brugia malayi worms harvested from untreated infected mice, is shown in Table 1. The data indicate that in vitro activities overlap for doxycycline and minocycline suggesting that both intrinsic antibiotic potency (in vitro data) and penetration of the filarial cuticle (in vitro worm data) are within the same range for both doxycycline and minocycline.
Pharmacokinetics of doxycycline and minocycline in BALB/c SCID mice
The pharmacokinetics of doxycycline and minocycline were determined for a 7 day 25 mg/Kg qd chronic dosing regimen (equivalent to the treatment regimens used in the murine infection studies). Table 2 shows the fitted pharmacokinetic profiles at steady state (day 7) for doxycycline and minocycline. Pharmacokinetic parameters for doxycycline day 1 were found to be not statistically different to those determined at steady state (Mann-Whitney test, n = 5, P-value > 0.05), this is consistent with there being no reports of time-dependent kinetics for tetracyclines in the literature. Figure 1 shows the predicted population PK profiles for doxycycline and minocycline. Minocycline was found to have a larger apparent clearance (CL/F: 3.2 vs. 1.2 L/h/Kg) and higher apparent volume of distribution than doxycycline (V/F: 13.2 vs. 6.9 L/hr). The elimination half-lives determined in BALB/c SCID mice were found to be comparable with those reported in the literature for doxycycline and minocycline in mice30,41. From Table 2 it can be seen that these differences lead to minocycline having approximately 3-fold less systemic exposure than doxycycline.

Pharmacokinetic profiles for doxycycline and minocycline at day 7.
Solid black line represents the median for a population of 1000 simulated individuals and the blue dashed lines depict the 5th and 95th percentile values for the population. Observed discreet concentrations are shown as grey circles.
Calculating murine dosage regimens bioequivalent to clinically relevant exposures
Monte Carlo simulations were used to calculate murine dosage regimens that give bioequivalent exposures to standard clinical 100–200 mg/day doses of second-generation tetracyclines. The distributions of human exposures of doxycycline and minocycline were based on pharmacokinetic parameters from the literature31,32. Figure 2 shows the simulated human pharmacokinetic profiles of 200 mg doses of doxycycline and minocycline overlaid with their respective bioequivalent murine pharmacokinetic profiles given as either once-daily or twice daily regimens. From simulation it has been determined that a 25 mg/Kg bid dosage regimen closely emulates overall daily exposure of a 100–200 mg clinical dose for both tetracyclines. For doxycycline oral doses of 100 mg and 200 mg in humans give mean exposures of 28.4 and 56.9 μg.hr/mL, whereas in mice 25 mg/Kg doses once or twice daily give mean exposures of 22.8 and 40.6 μg.hr/mL, respectively. For minocycline, 100 mg and 200 mg doses in the clinical yield exposures of 17.7 and 35.4 μg.hr/mL, whereas in mice 25 mg/Kg doses administered once or twice daily give exposures of 7.8 and 14.9 μg.hr/mL, respectively. Multiple daily doses are more preferable to higher dose once daily as this results in peak plasma concentrations that are more consistent with those most often achieved clinically with a standard doses of tetracycline and the length of exposure is more reflective of the clinical plasma concentration-time profile.

Concentration-time profiles for human and murine pharmacokinetic studies.
Median human pharmacokinetic profile (200 mg dose) is shown as a solid black line and the blue dashed lines depict the 5th and 95th percentile values for a population simulated population of 1000 individuals. Median 25 mg/Kg bid and 50 mg/Kg murine pharmacokinetic profiles are show in green and red solid lines, respectively.
Comparative in vivo efficacy of clinical bioequivalent doxycycline and minocycline regimens against adult stage Brugia malayi Wolbachia
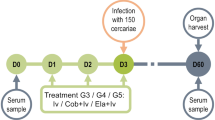
To directly compare efficacy of minocycline against doxycycline, dosages determined bioequivalent to typical human exposures; 25 mg/Kg either once or twice daily, were orally administered to B. malayi-infected SCID mice (Fig. 3a). Dose durations were extended up to 42 days, to match proven macrofilaricidal 4–6 week doxycycline clinical dosage regimens (defined as depletion of >90% Wolbachia from nematode tissues)14,33.

(a): Schematic of Brugia malayi (Bm) SCID mouse PD model with oral dosing regimens (MK: mg/Kg). (b): Effect on Wolbachia depletion following indicated human bioequivalent dosing of doxycycline (DOX) or minocycline (MIN). Data plotted is Wolbachia surface protein (wsp) single gene copy number per adult female worm, horizontal bars are median levels. Shaded area indicates levels above 90% depletion compared with median vehicle control level (dashed line). Data is derived from between 10 and 40 adult female worms pooled from groups of between 4–12 mice per drug treatment. Significant differences between the 13 treatment groups were identified by Kruskal-Wallis, two-tailed 1-way ANOVA, alpha = 0.05, P < 0.0001 with Dunn’s multiple tests post-hoc comparing drug groups vs vehicle control, indicated *P < 0.05, **P < 0.01 and ***P < 0.001. (c): Effect on microfilariae (mf) production following human bioequivalent dosing of doxycycline or minocycline. Data is total mf enumerated from the infection site in groups of 4 to 12 mice per drug group. Horizontal bars are median levels. Significant differences between the 13 treatment groups were identified by Kruskal-Wallis, two-tailed 1-way ANOVA, alpha = 0.05, P < 0.0001 with Dunn’s multiple tests post-hoc comparing drug groups vs vehicle control, indicated *P < 0.05, **P < 0.01 and ***P < 0.001.
Table 3 details the adult parasitological outcomes post-doxycycline or minocycline treatment. Approximately 90% (19/22) of vehicle control treated SCID mice harboured fecund, motile B. malayi at +12 weeks (median 6 worms/animal, range 0–19, sex ratio 59.2% female). As predicted for slow-acting macrofilaricides targeting Wolbachia, neither doxycycline nor minocycline, at any regimen tested, exerted a negative effect on adult filarial burdens at +12 weeks. Sex ratios were also not significantly distorted by drug treatment.
Median Wolbachia loads per female B. malayi were estimated at 2.57 × 107 (range 7.74 × 106–4.76 × 107) derived from untreated SCID mouse infections. Filarial Wolbachia loads from vehicle control treated mice levels were also similar (median 3.35 × 107, range 6.51 × 106–9.33 × 107). Doxycycline reduced Wolbachia in both a dose and dose-duration dependent-manner (Fig. 3b). In line with clinical data from doxycycline-treated LF patients and confirming bio-equivalent dose calibrations, doxycycline mediated significant (>90% threshold) depletion of Wolbachia in 28 days, dependent on dose (median reduction 90.35%, 25 mg/Kg bid x 28d). In comparison, minocycline mediated improved efficacy with reduced dosages and/or dose-durations. A 25 mg/Kg qd minocycline regimen yielded significant, yet sub-optimal, reductions in Wolbachia following 28 day dosing, which was an improvement compared with matching doxycycline dose delivered for 42 days (median reduction 85.08%, minocycline x 28d vs 66.30%, doxycycline x 42d, P = 0.0307, Mann-Whitney test). Increasing the dose of minocycline to 25 mg/Kg bid mediated a significant, above clinical threshold effect in depletion of >90% of Wolbachia following 28 day dosing, an improvement on matching doxycycline dose (median reduction 99.51%, minocycline vs 90.35%, doxycycline, P < 0.0001, Mann-Whitney test).
Depleting Wolbachia from filarial nematodes blocks embryogenesis34 and results in a waning of circulating LF mf in the blood or O. volvulus mf in the skin, which can be detected 2–4 months after start of effective doxycycline treatment14,18,22. Therefore, comparative effects of doxycycline and minocycline in blockading B. malayi embryogenesis were assessed in vivo by measuring accumulations of released mf at the peritoneal infection site (Fig. 3c). At 12 weeks post-infection, 89% of all vehicle treated mice contained mf (median 5396, range 0–23008). Doxycycline mediated a significant reduction in peritoneal mf loads following 28 or 42 days 25 mg/Kg qd doses and bid dosing of 25 mg/Kg for 28 days (Fig. 3c). However, at all doses and durations tested, at the evaluation endpoint blockade of embryogenesis was incomplete with between 20–50% of treated animals harbouring motile mf (Fig. 3c). In comparison, minocycline mediated significant reduction in peritoneal mf loads at all durations and doses tested, as little as 7 days at 25 mg/Kg qd, and rendered a complete blockade of embryogenesis after 3–4 weeks of exposure to 25 mg/Kg qd or bid regimens.
Immediate anti-Wolbachia pharmacological effect of escalating minocycline dose regimens
To assess the immediate pharmacological effect of minocycline dose regimens (i.e. without the impact of a washout period, post antibiotic effect) adult B. malayi parasitized mice were dosed with oral minocycline at 25, 40 or 80 mg/Kg bid for 7 days or 25 mg/Kg bid for 14 or 28 days (Fig. 4a). Adult worms were recovered immediately following the last dose and Wolbachia loads in female adult nematode tissue was enumerated (Fig. 4b). Seven day dosing of 25, 40 or 80 mg/Kg bid lead to sub-threshold, yet significant, depletions of Wolbachia (71.65–84.08% median depletions, 25–80 mg/Kg bid, Kruskal-Wallis 1-way ANOVA P < 0.0001). There were no significant changes in the level of Wolbachia depletion when comparing dose escalations between 25–80 mg/Kg bid over a 7-day course by 1-way ANOVA. When comparing extended dose durations, both 2 week and 4 week exposures led to >90% Wolbachia depletions (94.93% and 98.17%, respectively).

(a): Schematic of Brugia malayi (Bm) SCID mouse PD model with oral dosing regimens (MK: mg/Kg, ip: intraperitoneal route). (b): Effect on Wolbachia depletion following indicated minocycline (MIN). Data plotted is Wolbachia surface protein (wsp) single gene copy number per adult female worm, horizontal bars are median levels. Median vehicle control level is indicated by dashed line. Data is from 10–15 adult female worms pooled from between 5–10 mice per drug treatment. Significant differences between 7 day MIN dose escalation groups and vehicle were compared by Kruskal-Wallis, two-tailed 1-way ANOVA, alpha = 0.05, P < 0.0001 with Dunn’s multiple tests post-hoc comparing all groups indicated *P < 0.05, **P < 0.01 and ***P < 0.001. Significant differences between MIN 25MKbid for 14 or 28d vs corresponding length of vehicle were compared by Mann-Whitney tests, two-tailed, alpha = 0.05, P < 0.0001 indicated***.
Pharmacokinetic-Pharmacokinetic modelling
PKPD modelling incorporating an effect compartment was employed to assess the rate of Wolbachia reduction. The anti-Wolbachia potencies at the effect site were set equal to potencies determined in previously validated assay using Wolbachia infected cells and transfer coefficients to the effect site were fitted using non-parametric non-linear mixed effects modelling. Table 4 shows in vitro potencies and fitted transfer coefficients for both tetracyclines. The combination of minocycline’s increased potency and faster transfer to the effect compartment leads to 1.7-fold greater anti-Wolbachia effect in comparison to doxycycline with the transfer rate into the effect site being 24-fold higher for minocycline.
Discussions
The A∙WOL consortium aims to reduce the treatment time of anti-Wolbachia macrofilaricidal chemotherapies to improve test-and-treat drug regimen delivery, adherence and to reduce costs of treatment. Minocycline has been shown to have a similar anti-Wolbachia effect to doxycycline in vitro with improved in vivo efficacy, making it an ideal candidate for further PKPD investigation28.
Traditionally pre-clinical and clinical studies of anti-filarial chemotherapies have linked drug dosage to pharmacological effect35,36,37. Linking dosage to pharmacological effect when comparing chemotherapies is inherently flawed as the pharmacokinetics (absorption, distribution, metabolism and excretion) of different chemotherapies within a target population can vary radically and therefore the systemic exposure of the drug at the effect site can be significantly different for the same dose of two different drugs. This is an even greater problem in preclinical models where species and strain differences in PK can be radically different. The resultant pharmacological effect can therefore deviate for drugs with similar in vitro potencies. The use of rational drug development employing PKPD modelling or model-based drug development (MBDD) is a paradigm that has been used extremely effectively with preclinical and clinical development in other disease areas38,39,40. We propose, supported by the data presented here, that the NTD community, particularly those involved in the development of new chemotherapies, need to embrace these strategies if we are ever to optimise global treatment and elimination programmes to meet control and elimination targets.
In this work, for the first time, we use PKPD modelling to rationally develop an anti-filarial chemotherapy by linking drug exposure to pharmacological effect. Specifically, we use PKPD analysis to evaluate the relative efficacy of minocycline to the gold standard anti-Wolbachia macrofilaricide doxycycline in our Brugia malayi murine infection model of lymphatic filariasis. There is an inherent disconnect between the human pharmacokinetics and murine pharmacokinetics of tetracyclines with murine body weight normalised clearances being far greater than is the case in their human counterparts30,31,41. To address this disconnect we determined the murine pharmacokinetics of these tetracyclines, minocycline was found to have inferior pharmacokinetics to doxycycline, giving approximately 3-fold less exposure in comparison to doxycycline in our murine models. Monte-Carlo simulations of murine and human exposures were used to establish that a bi-daily 25 mg/Kg murine dose gave an approximately bioequivalent exposure to the currently recommended 100–200 mg daily clinical dose of the tetracycline. At bioequivalent exposures minocycline was found to deplete Wolbachia levels by >99% after a 28 day treatment regimen in the Brugia malayi murine model. This Wolbachia reduction is far greater than the 1 log-unit drop associated with irreversible sterilisation filarial tissues in clinical studies, the target clinical PD endpoint14,33. In contrast doxycycline at a 25 mg/Kg bid bioequivalent dosing regimen achieved a 90.35% reduction within the same 28 day time frame.
PKPD modelling was used to quantify that minocycline is 1.7-fold more effective than doxycycline when administered at the 25 mg/Kg bid bioequivalent dosage regimen, despite a 3-fold lower exposure than doxycycline and having a similar potency at the effect site as modelled by in vitro potencies. A PKPD model incorporating an effect compartment was employed; this enabled us to further dissect the origins of the superior efficacy of minocycline given its inferior systemic exposure and similar potency to doxycycline. These properties don’t account for the increase in efficacy indicating there are other factors influencing its in vivo efficacy. The transfer rates to and from the effect site were determined from the PKPD analysis, the transfer rate constant to the effect site was predicted to be 24-fold higher for minocycline in comparison to doxycycline whereas the efflux from the effect site was predicted to only marginal faster for minocycline. The difference in transfer rates in to the effect site could be due to increase bioavailability at the effect site from the systemic exposure or the transfer coefficient could be a lumped parameter masking the production of an active metabolite of minocycline that has a far greater potency than doxycycline. We have sought to explain the difference in transfer rate to the effect site through analysis of the physicochemical properties of doxycycline and minocycline focusing on physicochemical properties synonymous with permeability42,43, namely calculated octanol-water partition coefficient (ALogP), number of hydrogen bond acceptors/donors (H-Acceptor/H-Donor), molecular mass (MW), number of rotatable bonds (RotB) and molecular polar surface area (PSA). Figure 5 shows a radar plot of the physicochemical properties of doxycycline and minocycline, it can be seen that as expected the physicochemical properties of doxycycline and minocycline are similar, however there are notable differences. The optimal region for physicochemical properties with respect to permeability and oral bioavailability as defined by previous analyses of experimental in vitro and in vivo data42,43,44 are shown as shaded areas in Fig. 5. The physicochemical parameters of minocycline confer with more optimal regions of physicochemical space than doxycycline, in particular calculated LogP and number of hydrogen bond donors were sub-optimal for doxycycline in comparison to minocycline, furthermore minocycline’s molecular polar surface area (PSA) is more in-line with the recommend 140 Å2 cut-off limit for permeable compounds. These subtle differences may lead to a change in the transfer rates into the effect site.

(a). Physicochemical properties of doxycycline and minocycline shaded regions represents those synonymous with favourable permeation. (b). Chemical structures of doxycycline and minocycline.
The overall conclusion of this study is that minocycline is predicted to be superior to doxycycline as an anti-Wolbachia dependent macrofilaricide. The data suggest that not only can minocycline be used as an alternative to doxycycline in test and treat strategies it may offer some advantages through either improved Wolbachia kill rates or lower dosage requirements, which may impact on tolerability or safety profiles. Although the incidence of AEs with either drug is very low there is a slightly higher incidence of AE with minocycline45. The shorter treatment period may offset this slight increase in risk of AE with minocycline. It will now be important to undertake clinical evaluations of minocycline, supported through full PK analysis, to confirm these benefits. Recently, a clinical trial has shown a single dose triple combination of diethylcarbamazine, albendazole and ivermectin has shown microfilaremia suppression in Bancroftian Filariasis for up to a year or more. These results are clearly promising, with the potential to accelerate MDA endpoints in non-African settings, where co-endemicity with either Loa loa or Onchocerca volvulus do not present a risk of severe adverse events. Ultimately, a number of chemotherapeutic tools will be need to eliminate filariasis, including anti-Wolbachia agents for settings where standard anti-filarials are contraindicated or compromised by reduced efficacy46.
We used PKPD modelling to ensure that drug exposures for our dosing regimens in our Brugia malayi preclinical murine model are indicative of those of recommended clinical doses, given the Wobachia reduction achieved at bioequivalent doses of minocycline. We recommend that further clinical studies investigating the anti-Wolbachia effect of 2–4 weeks of 100 or 200 mg/day minocycline in patient population with lymphatic filariasis and onchocerciasis. Furthermore, treatment time maybe further truncated by combination with other classes of anti-Wolbachia registered drugs or in combination with current anti-filarial drugs, which are actively being pursued by A∙WOL.
Methods
Parasites
The parasite life cycle of B. malayi (Bm) filarial nematode (TRS strain)47 was maintained in mosquitoes and susceptible Meriones unguiculatus gerbils at LSTM. Infective Bm larvae (BmL3) were generated using protocols as previously described29. Briefly, female adult Aedes aegypti mosquitoes were fed with microfilariae collected from infected gerbils by catheterisation, as described previously48. Microfilariae were mixed with human blood and fed to mosquitoes using an artificial membrane feeder (Hemotek®). BmL3 stage larvae were propagated by rearing the blood-fed mosquitoes for 14 days. BmL3 larvae were then harvested from infected mosquitoes by crushing. Purification of the inoculum was performed using previously optimised protocols25.
Animals
Male BALB/c SCID mice were procured from Harlan Laboratories, UK. All animals were housed at an approved animal housing facility under specific pathogen-free (SPF) conditions. Approval was obtained for all animal experiments from the ethical committees of the University of Liverpool and LSTM. Experiments were conducted according to Home Office (UK) requirements.
Parasite Infections
100 freshly collected, motile Brugia malayi L3 larvae were injected via the intra-peritoneal route into BALB/c SCID mice. Efficiencies of inoculations were confirmed by needle washout.
Preparation and administration of Drugs/Compounds
Infected mice were administered 100 μL of either doxycycline or minocycline solution via oral gavage six weeks after infection. Doxycycline was administered once or twice daily at 25 mg/Kg for 7, 14, 21, 28 or 42 days. Minocycline was administered in dosage regimens mirroring those for doxycycline. Additionally, minocycline was administered twice daily at 25, 40 and 80 mg/Kg for 7 days. Both doxycycline and minocycline were dissolved in water and all reagents were purchased from Sigma unless otherwise stated.
Parasitological Readouts
Mice were necropsied 12 weeks after infection unless otherwise stated. A total of 5 individual screens were run, each containing groups of between 4/5 SCID mice per treatment arm. Peritoneal washes using RPMI medium were employed to recover adults and microfilariae (mf). Adult worms were removed and stored overnight in fresh medium at 4 °C before being washed in cold PBS. Adult stages were subsequently counted and their gender determined. For subsequent qPCR analysis of Wolbachia loads, 10–15 female adults were utilised per treatment group, derived from each screen. Between 2–4 female worms were selected, randomly, from individual mice, to avoid bias due to intra-group dosing variation. Mf were pelleted from peritoneal washings by centrifugation (300 g, 5 minutes, 4 °C, low brake), re-suspended in 1–2 ml of medium and the total mf quantity contained within peritoneal lavage enumerated by microscopy.
Quantification of Wolbachia bacteria numbers using qPCR assay
Extraction of DNA from individual female worms was performed using a DNeasy Blood and Tissue Kit (Qiagen) according to manufacturer’s instructions. Quantification of Bm Wolbachia wsp copy numbers was performed using qPCR as previously described49.
Determination of in vitro potencies
The anti-Wolbachia potency of doxycycline and minocycline for use in the PKPD model was determined in vitro, utilising the routine A·WOL screening assay as described previously50. In brief the mosquito (Aedes albopictus) derived cell line (C6/36), stably infected with Wolbachia pipientis (wAlbB) (C6/36 (wAlbB)) was incubated with the relevant drugs in a concentration range in order to determine a dose response. The drugs were incubated for 7 days with 2,000 cells per well on a 384 well plate (CellCarrier-384 Ultra, PerkinElmer) in Leibovitz media (Life Technologies™) supplemented with 20% foetal bovine serum (FBS, Fisher Scientific), 2% tryptose phosphate broth (Sigma-Aldrich) and 1% non-essential amino acids (Sigma-Aldrich). The end-point read out utilised DNA staining of both the host cell nuclei and intracellular Wolbachia (SYTO®11) combined with a high content imaging system (Operetta®, PerkinElmer) and analysed using the associated Harmony® software through a cytoplasm texture analysis.
Pharmacokinetic studies in BALB/c SCID mice
Rich pharmacokinetics studies were performed to characterise the pharmacokinetic profiles of orally administered doxycycline and minocycline in uninfected BALB/c SCID mice (weight 24–28 g). 25 mg/Kg of doxycycline or minocycline was administered via oral gavage in the appropriate vehicles (PEG300/Propylene Glycol/H20 (55%,25%,20%)). Blood samples were collected via microsampling from the tail vein. For both drugs, serial blood samples were collected up to 8 hours after a single dose and on day 7 after once daily dosing.
Blood samples (20 μL) were collected via a pipette with a pre-heparinised tip and were immediately lysed with 40 μL of ice cold ddH2O and subsequently frozen at −80 oC until time of analysis. Sparse samples were obtained from the pharmacodynamic studies in the Brugia malayi BALB/c SCID murine infection model to ensure that drug exposures were in line with those observed in the rich pharmacokinetic studies.
Bioanalysis
Doxycycline and minocycline plasma drug concentrations were quantified using LCMS (liquid chromatography mass spectrometry) on a UHPLC (ultrahigh pressure liquid chromatography) system linked to a triple-quadruple TSQ Quantum Access mass spectrometer (Thermo Scientific, Hemel Hempstead, UK) with a heated-electrospray ionization (H-ESI) source. Chromatographic separation was performed using a gradient via a Hypersil C18 Gold column (100 × 2.1 mm, 1.9 μm particle size) reverse-phase column (Thermo Scientific, UK). All methods incorporated the use of appropriate internal standards and were validated using FDA guidelines which are internationally recognised51.
For both doxycycline and minocycline samples were prepared as follows: 20 μL of blood sample containing the drug was extracted with 120 μL acetonitrile/methanol (80%/20%) containing the internal standard (doxycycline and minocycline at a concentration of 1 μg/mL were employed as internal standards for each other). Samples were then filtered using 96 well filter plates (MultiScreen Solvinert, 0.45 μm hydrophobic PTFE filter plates, Millipore Ltd, Ireland) and the filtrates were transferred to clean glass autosampler vials for analysis. Samples were assayed alongside a whole blood calibration curve (range 0 - 50 μg/mL), with quality control samples at low 0.5 μg/mL, medium 15 μg/mL and high 40 μg/mL concentrations for both doxycycline and minocycline. All standard curves were described using an equal weighted linear regression equation using data acquisition software LC Quan (Version 2.5.6.Thermo Scientific, Hemel Hempstead, UK). The correlation coefficient (r2) for both DOX and MN calibration curves exceeded 0.99. Lower limits of quantification for doxycycline and minocycline were 250 ng/mL and 300 ng/mL, respectively, with a signal-to-noise ratio of 5:1 and a coefficient of variation (CV) of less than 13% for both tetracyclines.
Pharmacokinetics-Pharmacodynamic Modelling
All pharmacokinetic and pharmacodynamics modelling and simulation was performed using Pmetrics52 within R version 3.1.053 A one-compartment model with an absorptive compartment for oral dosing was used for modelling the pharmacokinetics of doxycycline and minocycline as described by the differential equations 1a and 1b:


where X1 and X2 are the amounts of drug in the absorptive and central compartments, representing gut and systemic circulation, respectively. The pharmacokinetic parameters ka, CL, V represent the absorption rate constant, clearance and volume of distribution, respectively.
The link between drug exposure and pharmacological effect was quantified using PK/PD modelling. The pharmacodynamics response for the anti-Wolbachia effect exerted by doxycycline and minocycline was defined to be number of Wolbachia in each worm normalised to the relevant vehicle treated control and expressed as a percentage reduction. An effect compartment, X3, was used to account for the delay in observed effect as has been previously recommended54,55,56 and as defined by the differential equations 2a and 2b:


the parameters K12 and K21 in equations 2a and 2b represent transfer rate constants to and from the effect compartment. The pharmacodynamics response was modelled using an Emax model (equation 3) that depends on drug concentration in the effect compartment.

where EC50, Emax and Vf represent the maximum effect, the concentration to achieve 50% of the maximal effect and the effect site volume of distribution. The relative EC50 at the effect site was fixed to the in vitro EC50 determined in Wolbachia infected A. albopictus cells, Emax 99.999% and Vf is assumed to be proportional to the volume of distribution of the central compartment. The transfer rate constants, K12 and K21 to and from the effect compartment were fitted for each tetracycline.
Model fitting was performed via protocol used previously established protocols57,58 Briefly, linear regression (intercept close to 0, slope close to 1) was used to assess the goodness-of-fit of the observed/predicted values, the coefficient of determination of the linear regression and log-likelihood values. A statistically significant improvement in the log-likelihood value (P, 0.05) was required for a more complex model to be supported.
Statistical tests
Continuous variables (wsp copy number/female B. malayi worm and total peritoneal mf/SCID mouse) were pooled per treatment group from independent screens. Statistical significance of differences between independent groups was assessed using two-tailed non-parametric statistical hypothesis. The Mann-Whitney U test was employed when comparing two groups whereas the Kruskal-Wallis test followed by Dunn’s multiple comparisons test, post-hoc, was used in the case of comparing more than two groups. Significance level was set to alpha = 0.05.
Additional Information
How to cite this article: Sharma, R. et al. Minocycline as a re-purposed anti-Wolbachia macrofilaricide: superiority compared with doxycycline regimens in a murine infection model of human lymphatic filariasis. Sci. Rep. 6, 23458; doi: 10.1038/srep23458 (2016).
Change history
08 January 2018
Scientific Reports 6: Article number: 23458; published online: 21 March 2016; updated: 08 January 2018 The original version of this Article contained a typographical error in the spelling of the author Ghaith Aljayyoussi, which was incorrectly given as Ghaith Al Jayoussi. This has now been correctedin the PDF and HTML versions of the Article.
References
WHO, Lymphatic filariasis factsheet No. 102, (2015). Available at: http://www.who.int/mediacentre/factsheets/fs102/en/. (Accessed: 18th December 2015).
WHO, Onchocerciasis, fact sheet No. 374, (2015). Available at: http://www.who.int/mediacentre/factsheets/fs374/en/. (Accessed: 18th December 2015).
Taylor, M. J., Hoerauf, A. & Bockarie, M. Lymphatic filariasis and onchocerciasis. Lancet 376, 1175–1185 (2010).
Hoerauf, A., Pfarr, K., Mand, S., Debrah, A. Y. & Specht, S. Filariasis in Africa-treatment challenges and prospects. Clin. Microbiol. Infect 17, 977–985 (2011).
Gardon, J. et al. Serious reactions after mass treatment of onchocerciasis with ivermectin in an area endemic for Loa loa infection. Lancet 350, 18–22 (1997).
Boussinesq, M., Kamgno, J., Pion, S. D. & Gardon, J. What are the mechanisms associated with post-ivermectin serious adverse events? Trends Parasitol. 22, 244–246 (2006).
Osei-Atweneboana, M. Y. et al. Phenotypic evidence of emerging ivermectin resistance in Onchocerca volvulus. PLoS Negl. Trop. Dis. 5, e998 (2011).
Winnen, M. et al. Can ivermectin mass treatments eliminate onchocerciasis in Africa? Bull. World Health Organ. 80, 384–391 (2002).
Stolk, M. A., Walker, M., Coffeng, L. E., Basanez, M.-G. & de Vlas, S. J. Required duration of mass ivermectin treatment for onchocerciasis elimination in Africa: a comparative modelling analysis. Parasite. Vectors 8, 552 (2015).
Taylor, M. J., Hoerauf, A., Townson, S., Slatko, B. E. & Ward, S. A. Anti-Wolbachia drug discovery and development: safe macrofilaricides for onchocerciasis and lymphatic filariasis. Parasitol. 141, 119–27 (2014).
Turner, J. D. et al. Wolbachia Lipoprotein Stimulates Innate and Adaptive Immunity through Toll-like Receptors 2 and 6 to Induce Disease Manifestations of Filariasis. J. Biol. Chem. 284, 22364–22378 (2009).
Tamarozzi, F., Halliday, A., Gentil, K., Hoerauf, A., Pearlman, E. & Taylor, M. J. Onchocerciasis: the role of Wolbachia bacterial endosymbionts in parasite biology, disease pathogenesis and treatment. Clin. Microbiol. Rev. 24, 459–68 (2011).
Cross, H. F., Haarbrink, M., Egerton, G., Yazdanbakhsh, M. & Taylor, M. J. Severe reactions to filarial chemotherapy and release of Wolbachia endosymbionts into blood. Lancet 358, 1873–1875 (2001).
Turner, J. D. et al. A randomized, double-blind clinical trial of a 3-week course of doxycycline plus albendazole and ivermectin for the treatment of Wuchereria bancrofti infection. Clin. Infect. Dis. 42 (2006).
Chirgwin, S. R. et al. Removal of Wolbachia from Brugia pahangi is closely linked to worm death and fecundity but does not result in altered lymphatic lesion formation in Mongolian gerbils (Meriones unguiculatus). Infect. Immun. 71, 6986–6994 (2003).
Hoerauf, A. et al. Tetracycline therapy targets intracellular bacteria in the filarial nematode Litomosoides sigmodontis and results in filarial infertility. J. Clin. Invest. 103, 11–17 (1999).
Taylor, M. J. & Hoerauf, A. A new approach to the treatment of filariasis. Curr. Opin. Infect. Dis. 14, 727–731 (2001).
Taylor, M. J. et al. Macrofilaricidal activity after doxycycline treatment of Wuchereria bancrofti: a double-blind, randomised placebo-controlled trial. Lancet 365, 2116–2121 (2005).
Hoerauf, A. et al. Doxycycline as a novel strategy against bancroftian filariasis - depletion of Wolbachia endosymbionts from Wuchereria bancrofti and stop of microfilaria production. Med. Microbiol. Immunol. 192, 211–216 (2003).
Hoerauf, A., Mand, S., Adjei, O., Fleischer, B. & Buttner, D. W. Depletion of wolbachia endobacteria in Onchocerca volvulus by doxycycline and microfilaridermia after ivermectin treatment. Lancet 357, 1415–1416 (2001).
Hoerauf, A. et al. Wolbachia endobacteria depletion by doxycycline as antifilarial therapy has macrofilaricidal activity in onchocerciasis: a randomized placebo-controlled study. Med. Microbiol. Immunol. 197, 295–311 (2008).
Turner, J. D. et al. Macrofilaricidal activity after doxycycline only treatment of Onchocerca volvulus in an area of Loa loa co-endemicity: a randomized controlled trial. PLoS Negl. Trop. Dis. 4, e660 (2010).
Thong, Y. H. & Ferrante, A. Inhibition of mitogen-induced human-lymphocyte proliferation responses by tetracycline analogs. Clin. Exp. Immunol. 35, 443–446 (1979).
Kloppenburg, M. et al. The influence of tetracyclines on T cell activation. Clin. Exp. Immunol. 102, 635–641 (1995).
Debrah, A. Y. et al. Doxycycline reduces plasma VEGF-c/sVEGFR-3 and improves pathology in lymphatic filariasis. PLoS Pathog. 2, 829–843 (2006).
Mand, S. et al. Doxycycline Improves Filarial Lymphedema Independent of Active Filarial Infection: A Randomized Controlled Trial. Clin. Infect. Dis. 55, 621–630 (2012).
Walker, M. et al. Therapeutic Efficacy and Macrofilaricidal Activity of Doxycycline for the Treatment of River Blindness. Clin. Infect. Dis. 60, 1199–1207 (2015).
Johnston, K. L. et al. Repurposing of approved drugs from the human pharmacopoeia to target Wolbachia endosymbionts of onchocerciasis and lymphatic filariasis. Int. J. Parasitol.-Drugs Drug Resist. 4, 278–286 (2014).
Halliday, A. et al. A murine macrofilaricide pre-clinical screening model for onchocerciasis and lymphatic filariasis. Parasite. Vectors 7, 14 (2014).
Chang, H. R., Comte, R., Piguet, P. F. & Pechere, J. C. Activity of minocycline against toxoplasma-gondii infection in mice. J. Antimicrob. Chemother. 27, 639–645 (1991).
Saivin, S. & Houin, G. Clinical pharmacokinetics of doxycycline and minocycline. Clin. Pharmacokinet. 15, 355–366 (1988).
Agwuh, K. N. & MacGowan, A. Pharmacokinetics and pharmacodynamics of the tetracyclines including glycylcyclines. J. Antimicrob. Chemother. 58, 256–265 (2006).
Debrah, A. Y. et al. Macrofilaricidal effect of 4 weeks of treatment with doxycycline on Wuchereria bancrofti. Trop. Med. Int. Health 12, 1433–1441 (2007).
Landmann, F., Voronin, D., Sullivan, W. & Taylor, M. J. Anti-filarial Activity of Antibiotic Therapy Is Due to Extensive Apoptosis after Wolbachia Depletion from Filarial Nematodes. PLoS Pathog. 7, 11 (2011).
Devaney, E., Howells, R. E. & Smith, G. Brugia-pahangi in the balb/c mouse - a model for testing filaricidal compounds. J. Helminthol. 59, 95–99 (1985).
Townson, S., Dobinson, A., Connelly, C. & Muller, R. Chemotherapy of onchocerca-lienalis microfilariae in mice - a model for the evaluation of novel compounds for the treatment of onchocerciasis. J. Helminthol. 62, 181–194 (1988).
Neill, M. & Kazura, J. W. The effect of diethylcarbamazine in a murine model of Brugia malayi microfilaraemia. Bulletin of the World Health Organization 57, 329–330 (1979).
Chen, B., Dong, J. Q., Pan, W. J. & Ruiz, A. Pharmacokinetics/Pharmacodynamics Model-Supported Early Drug Development. Curr. Pharm. Biotechnol. 13, 1360–1375 (2012).
Milligan, P. A. et al. Model-Based Drug Development: A Rational Approach to Efficiently Accelerate Drug Development. Clin. Pharmacol. Ther. 93, 502–514 (2013).
Gabrielsson, J., Green, A. R. & Van der Graaf, P. H. Optimising in vivo pharmacology studies-Practical PKPD considerations. J. Pharmacol. Toxicol. Methods 61, 146–156 (2010).
Bocker, R. & Estler, C. J. Comparison of distribution of doxycycline in mice after oral and intravenous application measured by a high-performance liquid-chromatographic method. Arzneimittelforschung 31-2, 2116–2117 (1981).
Veber, D. F. et al. Molecular properties that influence the oral bioavailability of drug candidates. J. Med. Chem. 45, 2615–2623 (2002).
Lipinski, C. A., Lombardo, F., Dominy, B. W. & Feeney, P. J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliver. Rev. 23, 3–25 (1997).
Ghose, A. K., Viswanadhan, V. N. & Wendoloski, J. J. A knowledge-based approach in designing combinatorial or medicinal chemistry libraries for drug discovery. 1. A qualitative and quantitative characterization of known drug databases. J. Comb. Chem. 1, 55–68 (1999).
Smith, K. & Leyden, J. J. Safety of doxycycline and minocycline: A systematic review. Clin. Ther. 27, 1329–1342 (2005).
Thomsen, E. K. et al. Efficacy, Safety and Pharmacokinetics of Coadministered Diethylcarbamazine, Albendazole and Ivermectin for Treatment of Bancroftian Filariasis. Clin. Infect. Dis. 62, 334–341 (2016).
Taylor, M. J., Bilo, K., Cross, H. F., Archer, J. P. & Underwood, A. P. 16 S rDNA phylogeny and ultrastructural characterization of Wolbachia intracellular bacteria of the filarial nematodes Brugia malayi, B-pahangi and Wuchereria bancrofti. Exp. Parasitol. 91, 356–361 (1999).
Griffiths, K. G., Alworth, L. C., Harvey, S. B. & Michalski, M. L. Using an intravenous catheter to carry out abdominal lavage in the gerbil. Lab. Animal 39, 143–148 (2010).
McGarry, H. F., Egerton, G. L. & Taylor, M. J. Population dynamics of Wolbachia bacterial endosymbionts in Brugia malayi. Mol. Bioch. Parasitol. 135, 57–67 (2004).
Clare, R. H. et al. Development and Validation of a High-Throughput Anti-Wolbachia Whole-Cell Screen: A Route to Macrofilaricidal Drugs against Onchocerciasis and Lymphatic Filariasis. J. Biomol. Screen. 20, 64–69 (2015).
U.S. Food and Drug Administration (FDA). Guidances (Drugs). (2015). Available at: http://www.fda.gov/Drugs/GuidanceComplianceRegulatoryInformation/Guidances/default.htm. (Accessed: 18th December, 2015).
Neely, M. N., van Guilder, M. G., Yamada, W. M., Schumitzky, A. & Jelliffe, R. W. Accurate detection of outliers and subpopulations with Pmetrics, a nonparametric and parametric pharmacometric modeling and simulation package for R. Ther. Drug Monit. 34, 467–476 (2012).
R: A language and environment statistical computing. v. 3.1.0 (R Foundation for Statistical Computing, Vienna, Austria, 2014).
Holford, N. H. G. & Sheiner, L. B. Pharmacokinetic and pharmacodynamic modeling in vivo. CRC Crit. Rev. Bioeng. 5, 273–322 (1981).
Sheiner, L. B., Stanski, D. R., Vozeh, S., Miller, R. D. & Ham, J. Simultaneous modeling of pharmacokinetics and pharmacodynamics - application to d-tubocurarine. Clin. Pharmacol. Ther. 25, 358–371 (1979).
Holford, N. H. G. & Sheiner, L. B. Kinetics of pharmacologic response. Pharmacol. Ther. 16, 143–166 (1982).
Roberts, J. A. et al. Plasma and target-site subcutaneous tissue population pharmacokinetics and dosing simulations of cefazolin in post-trauma critically ill patients. J. Antimicrob. Chemother. 70, 1495–1502 (2015).
Grau, S. et al. Plasma and peritoneal fluid population pharmacokinetics of micafungin in post-surgical patients with severe peritonitis. J. Antimicrob. Chemother. 70, 2854–2861 (2015).
Acknowledgements
The A·WOL consortium is supported by a grant from the Bill and Melinda Gates Foundation awarded to the Liverpool School of Tropical Medicine. This work was also supported by Medical Research Council New Investigator Research Grant, MR/L018756/1 (JDT).
Author information
Authors and Affiliations
Contributions
R.S., J.D.T., S.A.W. and M.J.T. wrote the paper. R.S., G.A., J.D.T., S.A.W. and M.J.T. conceived the work. R.S. and G.A. performed the PK/PD modelling analysis. H.E.T., G.A., J.G., L.H., A.G., J.D., D.W., L.F. and J.D.T. performed or directed the preclinical in vivo work (PK or PD). H.E.T., G.A., J.G., L.H., A.G., J.D., D.W., D.A.N.C. and J.D.T. performed analysis to obtain quantitative biological readouts. D.W. performed the PK bioanalysis. D.A.N.C., L.J.M., R.H.C., A.C. and K.L.J. performed or directed the in vitro work. A.S., L.H. and D.A.N.C. maintained the parasite lifecycle.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Sharma, R., Jayoussi, G., Tyrer, H. et al. Minocycline as a re-purposed anti-Wolbachia macrofilaricide: superiority compared with doxycycline regimens in a murine infection model of human lymphatic filariasis. Sci Rep 6, 23458 (2016). https://doi.org/10.1038/srep23458
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep23458
This article is cited by
-
The pipeline for drugs for control and elimination of neglected tropical diseases: 2. Oral anti-infective drugs and drug combinations for off-label use
Parasites & Vectors (2023)
-
Acute minocycline administration reduces brain injury and improves long-term functional outcomes after delayed hypoxemia following traumatic brain injury
Acta Neuropathologica Communications (2022)
-
The Eagle effect in the Wolbachia-worm symbiosis
Parasites & Vectors (2021)
-
Comparison of immune responses to Loa loa stage-specific antigen extracts in Loa loa-exposed BALB/c mice upon clearance of infection
Parasites & Vectors (2020)
-
Mouse models of Loa loa
Nature Communications (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
