
Abstract
Dusky-like (Dyl) is associated with the morphogenesis of embryonic denticle, adult sensory bristle and wing hair in Drosophila melanogaster. And whether Dyl involved in insect post-embryonic development and its signal transduction are poorly understood. Here, phylogenetic analysis revealed that dyl displayed one-to-one orthologous relationship among insects. In Tribolium castaneum, dyl is abundantly expressed at the late embryonic stage. Tissue-specific expression analysis at the late adult stage illustrated high expression of dyl in the fat body and ovary. Knockdown of dyl resulted in the defects in larval epidermal pigmentation and completely blocked the transitions from larval to pupal and pupal to adult stages of T. castaneum. We further discovered that dyl RNAi phenotypes were phenocopied by blimp-1 or shavenbaby (svb) silencing and dyl was positively regulated by blimp-1 through svb in T. castaneum. These results suggest that Dyl functions downstream of Blimp-1 through Svb for larval epidermal pigmentation and metamorphosis. Moreover, ftz-f1 was down-regulated after RNA interference (RNAi) suppressing any of those three genes, indicating that Ftz-f1 works downstream of Dyl to mediate the effects of Blimp-1, Svb and Dyl on metamorphosis in T. castaneum. This study provides valuable insights into functions and signaling pathway of insect Dyl.
Similar content being viewed by others
Introduction
Dusky (Dy), Dusky-like (Dyl) and Miniature were identified as the transmembrane proteins containing a zona pellucid (ZP) domain in Drosophila melanogaster1. Phylogenetic analysis indicated that they have a common ancestor in D. melanogaster2. Sequence comparison illustrated that Dy has higher amino acid sequence identity with Dyl (70% amino acid sequence identity) than Miniature (only 45% amino acid sequence identity with Dy) at the ZP domains3.
RNA in situ hybridization and immunostaining showed that dy and miniature are expressed in pupal wings by 28 hours after puparium formation (APF), but the expression of dyl could not be detected in pupal wings at the same stage3. Consistent with their expressions, Dy and Miniature share similar roles for wing morphogenesis in D. melanogaster, either of which mutation reduced the size of the whole wings by decreasing the size of individual wing epidermal cell3. Different from Dy and Miniature, dyl mutant embryos were characterized by very small, unhooked denticles, often with a split extremely2. Knocking down dyl in adults resulted in stub bristles with pigmentation abnormalities4. Recently, it is found that dyl mutation caused split, thinned, multiple and often very short hairs and a unique planar cell polarity phenotype of adult wing in D. melanogaster5. These studies of Dyl only focused on embryonic denticles, adult bristles and wing hairs of D. melanogaster, which provides limited information about dyl functions in the insect. On the other hand, a confused problem has emerged among the current studies that dy is evolutionarily more close to dyl than miniature, whereas dy shows similar expressions and functions with miniature. Thus, investigation of the functions of Dyl in the insect will help to clarify the functional relationship among Dyl, Dy and Miniature.
At the transcriptional level, the epidermal expression of dyl was abolished in shavenbaby (svb) mutant embryos of D. melanogaster2, suggesting that Svb is upstream of Dyl. Svb is a key selector gene that integrates Wingless and DER pathways to control epidermis differentiation6 and it controls the expressions of miniature, singed and forked in embryonic denticles of D. melanogaster7. But the functional relationship between svb and dyl in insects is unclear up to now. Moreover, additional upstream genes and downstream genes of Dyl should be identified in the insect.
Recently, we discovered that not only dyl but also blimp-1 and ftz-f1 transcripts were down-regulated after RNA interference (RNAi) suppressing the G protein-coupled receptor gene, methuselah-like1 (Tcmthl1) (Accession number: HQ188199, orthologous to Drosophila mthl5) of Tribolium castaneum8, which is involved in lifespan, development, stress resistance and reproduction9, suggesting that there might be certain relationship among dyl, blimp-1 and ftz-f1. In D. melanogaster, blimp-1 RNAi caused lethality at the pupal stages in most of the observed animals10 and elevated expressions of ftz-f1 from 6 h11 to 8 h APF but reduced the expression level of ftz-f1 by 10 h APF10. While, Ftz-f1 was required for cholesterol converted into 20-hydroxyecdysone to control the metamorphosis in D. melanogaster12 and Leptinotarsa decemlineata (Say)13. Based on these clues, it is hypothesized that dyl might be associated with blimp-1 and ftz-f1 and involved in the metamorphosis of insect.
To address these issues, we investigated the function of Dyl in T. castaneum by RNAi in this study. Furthermore, we explored how dyl relates to Blimp-1, Svb and Ftz-f1. These Results indicate that Dyl, Svb and Blimp-1 are essential for larval epidermal pigmentation and metamorphosis in T. castaneum.
Results
Dyl is widely distributed and conserved in insects
Using Drosophila dyl as a reference, 20 homolog genes were obtained from the genome database of D. melanogaster, T. castaneum, Anopheles gambiae, Bombyx mori, Apis mellifera, Acyrthosiphon pisum and Pediculus humanus humanus. Phylogenetic analysis revealed that dyl has the orthologue in each insect and shows one-to-one orthologous relationship among insects. In addition, its two paralogues, dy and miniature, also have this orthologous relationship in insects, respectively. And dyl is evolutionarily close to dy but far away from miniature (Figs 1 and S1). Sequence alignment of the proteins encoded by these three genes from D. melanogaster and T. castaneum also demonstrated that Dyl shares more sequence identity with Dy than Miniature. Structurally, these three genes are coding for the transmembrane proteins containing a ZP domain, respectively (Fig. S2).

Phylogenetic tree of insect dy and dyl by Neighbor-joining method.
TC NO. indicates the T. castaneum gene; GB NO. indicates the A. mellifera gene; PHUM NO. indicates the P. h. humanus gene; ACYPI NO. indicates the A. pisum gene; BGIBMGA NO. and XP_004923535 indicate B. mori genes; AGAP NO. indicates A. gambiae gene; CG NO. indicates the D. melanogaster gene. The tree is rooted by D. melanogaster no mechanoreceptor potential A (nompA) (CG13207). The bootstrap value below 60% was removed from phylogenetic tree.
Developmental and tissue expressions of dyl in T. castaneum
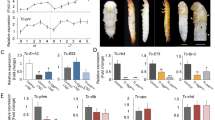
Before functional analysis of dyl, we measured mRNA levels of dyl at different stages and tissues by quantitative real-time PCR (qRT-PCR) analysis. In T. castaneum, dyl reached peak expression in the late embryonic stage, while its pupal and adult expression levels were very low (Fig. 2a). Tissue-specific expression analysis at the late adult stage illustrated dyl has high expressions in the fat body and ovary, moderate expressions in the epidermis and accessory gland but low expressions in the gut (Fig. 2b).

Developmental (a) and tissue (b) expression patterns of dyl in T. castaneum by qRT-PCR. EE, early eggs; LE, late eggs; EL, early larvae; LL, last-instar larvae; EP, early pupae; LP, late pupae; EA, early adults; LA, late adults. Tissues were isolated from adults (10-day old). One μg of total RNA from each sample was converted into cDNA for qRT-PCR.
dyl is essential for larval epidermal pigmentation and metamorphosis of T. castaneum
To clarify the function of dyl (Accession number: TC001440), dsRNAs of dyl were injected into larvae and pupae of T. castaneum. The qRT-PCR analysis indicated that the mRNA levels of dyl were significantly reduced after dsRNA injection (Fig. 3a). It is observed that all dyl dsRNA-treated insects arrested at the larval stage and failed to molt into pupae. The epidermis of these larvae has pigmentation abnormalities. Injection of dyl dsRNA into early pupae resulted in developmental arrest prior to eclosion in T. castaneum (Fig. 3a). Thus, dyl is essential for larval epidermal pigmentation and metamorphosis of T. castaneum.

Knockdown of dyl affected T. castaneum pupation, eclosion (a) and reduced ftz-f1 level (b). IB, beetles injected with physiological buffer; ds-dyl, beetles injected with dyl dsRNA. Three individuals of each group were used to extract total RNA for qRT-PCR analysis.
Dyl positively regulates ftz-f1 expression in T. castaneum
It is shown that reduction of dyl strongly inhibited the metamorphosis in T. castaneum and it is likely that there is cross-talk between Dyl and 20-hydroxyecdysone pathways. Therefore, we decided to check the expression of ftz-f1 that is required for 20-hydroxyecdysone synthesis. As expected, we found that the mRNA level of ftz-f1 (Accession number: TC002550) was down-regulated by nearly 89% compared with IB after dyl knockdown (Fig. 3b), suggesting that dyl positively regulates ftz-f1 expression in T. castaneum. The result illustrates that Dyl is implicated in 20-hydroxyecdysone pathway.
Svb plays vital roles in larval epidermal pigmentation, metamorphosis and promotes expressions of dyl, miniature, singed and forked in T. castaneum
By RNAi, svb (Accession number: TC008099) was silenced at larval and pupal stages in T. castaneum, respectively. Knock-down of svb resulted in arrested development at the larval stage of T. castaneum. These larvae have abnormally pigmented epidermis, could not molt into pupae and then died at larval stage. Pupal RNAi of svb also led to the failure to initiate the eclosion of T. castaneum (Fig. 4a). In svb dsRNA-treated insects, approximately 85% of dyl mRNA level was eliminated. It is also observed that more than 50% expressions of miniature (Accession number: TC003770), singed (Accession number: TC006673) and forked (Accession number: TC005627) were declined by svb RNAi (Fig. 4b). Thus, dyl, miniature, singed and forked are indeed the targets of Svb in T. castaneum. Furthermore, svb knockdown abolished 89% transcript of ftz-f1 (Fig. 4c). Taken together, these results indicate that Svb is required for larval epidermal pigmentation and metamorphosis and promotes the expressions of dyl, miniature, singed and forked in T. castaneum.

Svb RNAi affected pupation and eclosion (a) and reduced the expressions of dyl, miniature, singed, forked (b) and ftz-f1 (c) in T. castaneum. IB, beetles injected with physiological buffer; ds-svb, beetles injected with svb dsRNA. Three larvae or pupae of each group were used to extract total RNA for qRT-PCR analysis.
Blimp-1 is required for metamorphosis, epidermal pigmentation and positively regulates svb and its targets and ftz-f1 expressions in T. castaneum
Injections of dsRNAs for blimp-1 (Accession number: TC014741) into larvae or pupae depleted about 90% mRNA of itself in T. castaneum. Reduced blimp-1 levels caused developmental arrest at the larval stage of T. castaneum. The treated larvae with abnormally pigmented epidermis could not molt into pupae. Injection of blimp-1 dsRNAs into pupae led to the failure to start the eclosion (Fig. 5a). By qRT-PCR, we demonstrated that blimp-1 knockdown significantly reduced mRNA levels of svb and its targets and ftz-f1 in T. castaneum (Fig. 5b,c). Overall, Blimp-1 is also essential for larval epidermal pigmentation, metamorphosis and positively modulates expressions of svb and its targets in T. castaneum.

Blimp-1 RNAi affected pupation and eclosion (a) and down-regulated the mRNA levels of svb and its targeted genes (b) and ftz-f1 (c) in T. castaneum. IB, beetles injected with physiological buffer; ds-blimp-1, beetles injected with blimp-1 dsRNA. Three larvae or pupae of each group were used to extract total RNA for qRT-PCR analysis.
Discussion
In this study, we reveal that dyl, svb and blimp-1 are required for larval epidermal pigmentation and metamorphosis of T. castaneum (Figs 3a, 4a and 5a). At the transcriptional level, dyl positively regulated ftz-f1 expression but was modulated by blimp-1 and svb (Figs 3, 4, 5).
In D. melanogaster, dyl was detected in trichome cells of embryo epidermis but not pupal wings2,3. While, dyl mutation or knockdown resulted in very small and unhooked denticles of embryo2, stub bristles with pigmentation abnormalities4, split, thinned, multiple and often very short hairs and a unique planar cell polarity phenotype of adult wing5 in D. melanogaster. In this study, dyl also showed high expressions in the late embryonic stage and low expressions in the early pupal stage of T. castaneum (Fig. 2a). However, RNAi knocking down dyl at the late larval and early pupal stages caused the defects in larval epidermal pigmentation and metamorphosis of T. castaneum (Fig. 3). Moreover, parental RNAi of dyl led to significant embryonic lethal effect in T. castaneum, indicating that dyl also played some key roles in embryo development of T. castaneum, which may be much more critical than that of it in D. melanogaster. Tissue-specific analysis revealed that dyl is highly expressed in the fat body and ovary of adult in T. castaneum (Fig. 2b), supporting the embryonic lethal effect induced by parental RNAi of dyl. Thus, it seems that dyl shares some common functions between D. melanogaster and T. castaneum, but has occurred functional shift between them.
In D. melanogaster, the epidermal expression of dyl was abolished or reduced in svb mutant embryos2, indicating that dyl is positively regulated by svb. In addition to dyl, the expressions of miniature, singed and forked were also controlled by svb in D. melanogaster7. In T. castaneum, svb knockdown reduced dyl, miniature, singed and forked (Fig. 4b), whereas dyl silencing showed no effects on svb and forked expressions (Fig. S3). RNAi of dyl or svb led to the defects in larval epidermal pigmentation and metamorphosis in T. castaneum (Figs 3a and 4a). These results confirm that Svb is upstream of Dyl for larval epidermal pigmentation and metamorphosis.
In D. melanogaster, blimp-1 knockdown showed lethality at pupal stages in most of the observed animals. Many of them eclosed but died shortly thereafter or died during eclosion10. Interestingly, silence of blimp-1 also resulted in the defects of larval epidermal pigmentation and metamorphosis of T. castaneum (Fig. 5a), which is similar to dyl or svb RNAi phenotypes (Figs 3a and 4a). At the transcriptional level, blimp-1 RNAi reduced the levels of svb, dyl, miniature, singed and forked in T. castaneum (Fig. 5b). Therefore, Dyl is downstream of Blimp-1 through Svb for larval epidermal pigmentation and metamorphosis in T. castaneum.
In blimp-1 knockdown flies, high-level expressions of ftz-f1 was detected from 6h11 to 8h APF, but the expression level of ftz-f1 was greatly reduced by 10 h APF, demonstrating that Blimp-1 works as a repressor for premature expression of ftz-f1 but a activator for latish expression of ftz-f110. By qRT-PCR analysis, it is shown that blimp-1 knockdown down-regulated ftz-f1 expression at the last-instar larvae of T. castaneum (Fig. 5c), suggesting that the expression of ftz-f1 is positively regulated by blimp-1 at the last-instar larvae. Likewise, svb or dyl RNAi also reduced ftz-f1 level in T. castaneum (Figs 3b and 4c). In T. castaneum, high expression levels of dyl (Fig. 2b) and ftz-f114 were detected in the adult fat body and ovary, demonstrating that dyl and ftz-f1 possessed the similar tissue-specific expression pattern. These results indicated that Ftz-f1 works downstream of Dyl, Blimp-1 and Svb in T. castaneum. As reported previously, Ftz-f1 was required for cholesterol converted into 20-hydroxyecdysone in D. melanogaster12 and L. decemlineata (Say)13. Mutation of ftz-f1 resulted in arrested development at embryonic, larval and pupal stages in D. melanogaster15. Reduced levels of ftz-f1 by RNAi also caused arrested development at larval stage12. These studies demonstrate that ftz-f1 is essential for metamorphosis. Therefore, the effects of Dyl, Svb and Blimp-1 on metamorphosis were modulated by Ftz-f1 in T. castaneum. It is concluded that Dyl functions downstream of Blimp-1 through Svb but upstream of Ftz-f1 for metamorphosis (Fig. 6).

Preliminary regulation model of Dyl in T. castaneum.
Developmental study showed that the presence/absence of svb expression ultimately determined the pattern of denticles and dorsal hairs16. It was further found that Svb controls the expression of cuticle proteins17 and enzymes that increase trichome pigmentation and hardness7. These studies reveal the vital roles of Svb for epidermis differentiation. In T. castaneum, an obvious defect in epidermal pigmentation was observed in svb knockdown larvae (Fig. 4a), confirming the involvement of Svb in epidermal pigmentation. As the target of Svb, dyl was only expressed in embryonic tissues that will secrete cuticle, including epidermis, trachea and foregut2,3. Knocking down dyl function disrupted cuticle formation in bristles4. As is the case in bristles, hairs lacking dyl function also showed abnormality in chitin deposition5. In T. castaneum, dyl silencing caused defects in larval epidermal pigmentation (Fig. 3a). In D. melanogaster, Chitinase 6 was identified as a potential candidate of Dyl5. It is likely that epidermal pigmentation defects arose from dyl, svb or blimp-1 knockdown are associated with chitin deposition. However, how Dyl, Svb and Blimp-1 modulate larval epidermal pigmentation is unclear at present and need to be studied in the future.
Though dyl, dy and miniature of D. melanogaster were originated from one ancestral gene2, sequence comparison illustrated that Dyl showed more sequence similarities with Dy than Miniature at ZP domains3. Based on seven insect genome sequences, our study shows that insect dyl, dy and miniature were divided into three separate clusters of the phylogenetic tree (Figs 1 and S1), providing clues to the functional relationship among these three genes. In D. melanogaster, dy and miniature were detected in pupal wings by 28 hours APF, while dyl expression was not detected in pupal wings of the same stage3. Functionally, both Dy and Miniature are required for cytoskeletal reorganisation during wing morphogenesis3, while dyl shows significant effects on embryonic denticles formation2, adult bristle cuticle formation4 and adult hair integrity and planar cell polarity in D. melanogaster5. In T. castaneum, dyl is shown to be required for larval epidermal pigmentation and metamorphosis (Fig. 3a). These results suggest that Dyl is functionally divergent with Dy and Miniature in the insect.
Methods
Insect strains
The Tribolium castaneum GA-1 strain was used for all experiments. Insects were reared in whole wheat flour containing 5% brewer’s yeast at 30 °C under standard conditions as described previously18,19.
RNA extraction and cDNA synthesis
Using RNAisoTMPlus (TaKaRa), total RNA was isolated from the eggs, larvae, pupae and adults of T. castaneum. And 1 μg of total RNA was converted to cDNA by Moloney Murine Leukemia Virus reverse transcriptase (TaKaRa) and an Oligo(dT)18 primer (TaKaRa).
Phylogenetic analysis
With the sequences of D. melanogaster dy, dyl and miniature (http://flybase.org/), we searched the genome databases of these insects, including T. castaneum (http://beetlebase.org/), A. gambiae (https://www.vectorbase.org/index.php), B. mori (http://silkworm.genomics.org.cn/), A. mellifera (http://hymenopteragenome.org/beebase/), A. pisum (http://www.aphidbase.com/aphidbase/), P. h. humanus (https://www.vectorbase.org/index.php) and National Center for Biotechnology Information (NCBI, http://www.ncbi.nlm.nih.gov/guide/). These sequences were aligned with Clustal Omega (http://www.ebi.ac.uk/Tools/msa/clustalo/). Then the Neighbour-joining tree was reconstructed by MEGA 5 using the bootstrap method with 1000 replications (Fig. 1). To test the topology of Neighbor-joining tree, we reconstructed the Maximum likelihood tree by MEGA 5 (Fig. S1).
qRT-PCR
To check the gene expression profiles, qRT-PCR was performed with FastStart Universal SYBR Green Master (ROX) (Roche). The data are expressed here as the relative mRNA levels normalized to a control gene, T. castaneum ribosomal protein S3 (rps3)20, using the ΔΔCT method21. Three batches of samples were used for qRT-PCR. The primers were listed in Table 1.
RNAi
RNAi was performed as previously9,20. Negative controls consisted of injections of either vermillion (ver) dsRNA or an equal volume of buffer only (IB). Knock-down levels of the target genes were examined by qRT-PCR at the fifth day after injection. These experiments were performed for three biological replications.
Statistical analysis
The mean values of the RNAi-treated insects versus the mean values of the control insects were compared using the one-way ANOVA program of SPSS version 13.0. All the data are presented as the mean ± standard error. “*” indicates p < 0.05 and “**” indicates p < 0.001.
Additional Information
How to cite this article: Li, C. et al. Dusky-like is required for epidermal pigmentation and metamorphosis in Tribolium castaneum. Sci. Rep. 6, 20102; doi: 10.1038/srep20102 (2016).
References
Jazwinska, A. & Affolter, M. A family of genes encoding zona pellucida (ZP) domain proteins is expressed in various epithelial tissues during Drosophila embryogenesis. Gene Expr. Patterns 4, 413–421 (2004).
Fernandes, I. et al. Zona pellucida domain proteins remodel the apical compartment for localized cell shape changes. Dev. Cell 18, 64–76 (2010).
Roch, F., Alonso, C. R. & Akam, M. Drosophila miniature and dusky encode ZP proteins required for cytoskeletal reorganisation during wing morphogenesis. J. Cell Sci. 116, 1199–1207 (2003).
Nagaraj, R. & Adler, P. N. Dusky-like functions as a Rab11 effector for the deposition of cuticle during Drosophila bristle development. Development 139, 906–916 (2012).
Adler, P. N., Sobala, L. F., Thom, D. & Nagaraj, R. dusky-like is required to maintain the integrity and planar cell polarity of hairs during the development of the Drosophila wing. Dev. Biol. 379, 76–91 (2013).
Payre, F., Vincent, A. & Carreno, S. ovo/svb integrates Wingless and DER pathways to control epidermis differentiation. Nature 400, 271–275 (1999).
Chanut-Delalande, H., Fernandes, I., Roch, F., Payre, F. & Plaza, S. Shavenbaby couples patterning to epidermal cell shape control. PLoS Biol. 4, e290 (2006).
Li, C. et al. Comparative RNA-sequencing analysis of mthl1 functions and signal transductions in Tribolium castaneum. Gene 547, 310–318 (2014).
Li, C. et al. Methuselah-like genes affect development, stress resistance, lifespan and reproduction in Tribolium castaneum. Insect Mol. Biol. 23, 587–597 (2014).
Agawa, Y. et al. Drosophila Blimp-1 is a transient transcriptional repressor that controls timing of the ecdysone-induced developmental pathway. Mol. Cell. Biol. 27, 8739–8747 (2007).
Murata, T., Kageyama, Y., Hirose, S. & Ueda, H. Regulation of the EDG84A gene by FTZ-F1 during metamorphosis in Drosophila melanogaster. Mol. Cell. Biol. 16, 6509–6515 (1996).
Talamillo, A. et al. Scavenger receptors mediate the role of SUMO and Ftz-f1 in Drosophila steroidogenesis. PLoS Genet. 9, e1003473 (2013).
Liu, X. P. et al. Involvement of FTZ-F1 in the regulation of pupation in Leptinotarsa decemlineata (Say). Insect Biochem. Mol. Biol. 55C, 51–60 (2014).
Xu, J., Tan, A. & Palli, S. R. The function of nuclear receptors in regulation of female reproduction and embryogenesis in the red flour beetle, Tribolium castaneum. J. Insect Physiol. 56, 1471–1480 (2010).
Yamada, M. et al. Temporally restricted expression of transcription factor betaFTZ-F1: significance for embryogenesis, molting and metamorphosis in Drosophila melanogaster. Development 127, 5083–5092 (2000).
Delon, I. & Payre, F. Evolution of larval morphology in flies: get in shape with shavenbaby. Trends Genet. 20, 305–313 (2004).
Andrew, D. J. & Baker, B. S. Expression of the Drosophila secreted cuticle protein 73 (dsc73) requires shavenbaby. Dev. Dynam. 237, 1198–1206 (2008).
Arakane, Y. et al. Functional analysis of four neuropeptides, EH, ETH, CCAP and bursicon and their receptors in adult ecdysis behavior of the red flour beetle, Tribolium castaneum. Mech. Dev. 125, 984–995 (2008).
Beeman, R. W. & Stuart, J. J. A Gene for Lindane + Cyclodiene Resistance in the Red Flour Beetle (Coleoptera, Tenebrionidae). J. Econ. Entomol. 83, 1745–1751 (1990).
Begum, K., Li, B., Beeman, R. W. & Park, Y. Functions of ion transport peptide and ion transport peptide-like in the red flour beetle Tribolium castaneum. Insect Biochem. Mol. Biol. 39, 717–725 (2009).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 25, 402–408 (2001).
Acknowledgements
This work was supported by the National Natural Science Foundation of China (No. 31172146 & 31572326) and the PAPD of Jiangsu Higher Education Institutions.
Author information
Authors and Affiliations
Contributions
Conceived and designed the experiments: C.J.L. and B.L. Performed the experiments: C.J.L. and X.P.Y. Analyzed the data: C.J.L. and B.L. Contributed reagents/materials/analysis tools: C.J.L. and X.P.Y. Wrote the paper: C.J.L. and B.L. All authors gave final approval for publication.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Li, C., Yun, X. & Li, B. Dusky-like is required for epidermal pigmentation and metamorphosis in Tribolium castaneum. Sci Rep 6, 20102 (2016). https://doi.org/10.1038/srep20102
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep20102
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
