
Abstract
Mendelian randomisation studies from Asia suggest detrimental influences of alcohol on cardiovascular risk factors, but such associations are observed mainly in men. The absence of associations of genetic variants (e.g. rs671 in ALDH2) with such risk factors in women – who drank little in these populations – provides evidence that the observations are not due to genetic pleiotropy. Here, we present a Mendelian randomisation study in a South Korean population (3,365 men and 3,787 women) that 1) provides robust evidence that alcohol consumption adversely affects several cardiovascular disease risk factors, including blood pressure, waist to hip ratio, fasting blood glucose and triglyceride levels. Alcohol also increases HDL cholesterol and lowers LDL cholesterol. Our study also 2) replicates sex differences in associations which suggests pleiotropy does not underlie the associations, 3) provides further evidence that association is not due to pleiotropy by showing null effects in male non-drinkers and 4) illustrates a way to measure population-level association where alcohol intake is stratified by sex. In conclusion, population-level instrumental variable estimation (utilizing interaction of rs671 in ALDH2 and sex as an instrument) strengthens causal inference regarding the largely adverse influence of alcohol intake on cardiovascular health in an Asian population.
Similar content being viewed by others

Introduction
Previous epidemiological studies have reported potential beneficial effects of moderate alcohol intake on cardiovascular health1,2. In a recent review paper combining results from 84 observational studies, moderate drinkers were shown to have reduced risks of cardiovascular disease outcomes compared with non-drinkers, although heavy drinkers had the highest risks of all3. However, such evidence is not adequate for promotion of moderate alcohol use in prevention of heart disease given the known limitations of observational studies2,4. First, observed cardio-protective effects may be a form of reverse causation whereby individuals with the early stages of disease reduce their alcohol intake2,4,5. Second, observed effects might be due to confounding factors such as socioeconomic position, diet or other health-related behaviours and therapeutic regimes4. Therefore, the causal nature of association beyond observed correlation must be investigated in order to fully evaluate the benefits or harms of alcohol use.
Potential relationships should be interrogated through methods that can manipulate exposure and observe corresponding outcomes while accounting for confounding factors6. The gold standard is a randomised controlled trial (RCT), but this may be impossible to implement, prohibitively expensive or unethical. One alternative approach is that of Mendelian randomisation7. This method utilises a genetic variant that is allocated at conception in a manner that is independent of environment; people with the same genotype are thus akin to randomly allocated group of people in an RCT8. In essence, Mendelian randomisation exploits the idea such that a genetic variant, which proxies for the exposure, is expected to be related to the outcome to the degree anticipated given its association with the exposure. When Mendelian randomisation is implemented as a form of instrumental variable analysis, the genetic variant is referred to as an instrumental variable (IV)9. Using a Mendelian randomisation approach, causal effects of alcohol intake on cardiovascular outcomes have been investigated in several studies5,10,11,12. Robust IVs for alcohol intake include genetic variants in aldehyde dehydrogenase 2 (ALDH2)13,14 and alcohol dehydrogenase 1B (ADH1B)10. Both these genes are involved in alcohol metabolism (Supplementary Fig. S1), although the ALDH2 variants have substantially more influence on alcohol intake than the ADH1B variants10. The ALDH2 variants are polymorphic mainly in East Asian populations (Supplementary Fig. S2). Individuals who carry the variant allele experience on average greater discomfort after drinking alcohol, including nausea and facial flushing (so called Asian flush) since the variant allele codes for an inactive form of the enzyme, that leads to build-up of acetaldehyde in the circulation following alcohol consumption. As a result, carriage of the ALDH2 variant has consistently been linked with drinking behaviours15,16,17,18,19 and alcohol related diseases or risk factors in a number of Asian population studies11,15,20,21,22,23,24,25,26. For example, these studies consistently suggest that alcohol intake is associated with higher blood pressure21,24, not only in heavy drinkers but also in moderate drinkers11, corroborating some observational epidemiological studies3. Use of the variant also implies alcohol drinking is associated with coronary artery disease25 and coronary spastic angina26. On the other hand, some studies suggest alcohol drinking may have a favourable influence by increasing high density lipoprotein (HDL) cholesterol or decreasing low density lipoprotein (LDL) cholesterol11,22. However, these findings are not always robust10,25 and furthermore, the causal relationship between HDL cholesterol and cardiovascular health is uncertain27,28.
For cardiovascular outcomes showing association with the ALDH2 variant, associations have largely been confined to Asian men5,21,24. Weak or null associations observed in Asian women are due to a low level of alcohol consumption in females irrespective of the genotype, which is analogous to the situation within a RCT framework where randomly allocated groups receiving a very low amount of exposure would not result in any difference in outcomes21. The difference of associations between men and women provides an excellent rationale that the variant influences outcomes only through the exposure (i.e. alcohol intake), validating an assumption of Mendelian randomisation. The reasoning behind this is that if it were not the case – for example, if pleiotropic effects of the genetic variant influenced the outcomes - the same association between the variant and outcomes would have been seen in women as well as in men, as discussed in detail elsewhere5,20. Nevertheless, this sex stratification of alcohol intake also raises a question whether using the rs671 genotype alone as an IV would be sufficient to properly assesses causal effects in the whole population when both genetic variants and sex influences on alcohol intake should be considered29.
In this study, we carried out a Mendelian randomisation study to investigate the causal effects of alcohol intake on a range of cardiovascular outcomes and included the stratification of alcohol intake by gender. Data were collected from a total of 7,152 individuals from South Korea, including 3,365 men and 3,787 women. First, causal effects of alcohol intake were investigated in men and women separately, by conventional IV models using the rs671 genotype in ALDH2 as an IV. To demonstrate that the observed sex differences of association between the variant and cardiovascular outcomes were due to difference in corresponding drinking level rather than some particular influence of sex, the male-specific association was subsequently evaluated in sub-groups of never-drinkers and ever-drinkers. Finally, population-level causal effects were estimated by an extended IV model utilising interaction of the rs671 genotype and sex as an IV.
Results
General characteristics
Basic characteristics of male and female participants are shown in Table 1. Mean values were different between men and women in most variables with the exception of hip circumference, total cholesterol, the rs671 genotype and genotypic principal components. Men were younger, more likely to live in urban areas, more educated, doing less exercise and smoking more than women on average. Alcohol intake was considerably higher in men than women; 72% of men were current drinkers compared to 26% of women; the average alcohol intake was 18.8 ± 0.5 (g/day) in men and 1.3 ± 0.1 (g/day) in women; and gamma- glutamyl transpeptidase (GGT) was 55.4 ± 1.6 (IU/L) in men and 19.0 ± 0.3 (IU/L) in women. Men had higher prevalence of diseases and more generally unfavourable risk factors than women, although men had a few more generally favourable values as well (lower body mass index (BMI) and LDL cholesterol). It should be noted that there was no difference between men and women in the prevalence of rs671 genotype and genotypic principal components.
Characteristics were also provided according to the rs671 genotype in each sex group (Table 2). In both men and women, the rs671 genotype was in Hardy-Weinberg equilibrium with the A-allele frequency of 16%. The rs671 genotype was not associated with lifestyle or socioeconomic factors. Regarding alcohol intake, carriers of the rs671 A-allele had a lower proportion of current drinkers and consumed less alcohol than non-carriers in both men and women, though the magnitude of difference was bigger in men. With regard to disease prevalence and related risk factors, carriers of the rs671 A-allele appeared to have several potentially beneficial effects and a few potentially adverse effects than non-carriers (Table 2). All these associations were observed only in men and not in women. In addition to lifestyle or disease related factors, potential population stratification of the rs671 genotype was investigated through its association against the first five genotypic principal components. The second and the fourth principal components were correlated with the rs671 genotype in men (p = 0.02) and women (p = 0.04), respectively. Hence, these two principal components were included in the subsequent Mendelian randomisation analysis to correct for population stratification.
Next, the male population was divided into two groups by their drinking status: ever-drinkers and never-drinkers and then corresponding characteristics in each group were provided according to the rs671 genotype (Table 3). In this stratified analysis, potential collider bias30 was tested using generalized regression models with an interaction of genotype and drinking behaviour. Strong evidence of collider bias was observed for smoking behaviour (interaction p < 0.0001) where its association with genotype by strata were in opposite directions (Table 3). To minimize the effect of risk factors susceptible to collider bias, associations between the rs671 genotypes and cardiovascular outcomes were then assessed with adjustments for smoking with the results being closely similar to those without adjustments. In male ever-drinkers, the rs671 A-allele was associated with several potentially beneficial effects and a few potentially adverse effects after adjustments (Table 3). These associations were not observed in male never-drinkers, apart from weak associations with waist to hip ratio and fasting glucose level.
Observational associations
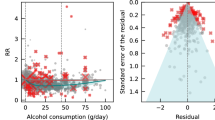
Association results based on the ordinary least squares (OLS) regression models can be found in Table 4 and Supplementary Tables S1. In men, alcohol intake was shown to be associated with higher hypertension risks, blood pressure, BMI, waist circumference, waist to hip ratio, log-transformed fasting blood glucose, HDL cholesterol, log-transformed triglycerides as well as with lower LDL cholesterol. In women, alcohol intake was associated with higher hypertension risks, blood pressure, BMI, hip circumference, log-transformed fasting glucose, total cholesterol and HDL cholesterol. The heterogeneity of OLS estimates in men and women was observed for diastolic blood pressure, log-transformed fasting blood glucose and HDL cholesterol and marginally for hypertension, hip circumference and total cholesterol under the fixed effect model. All regression analysis results were inspected based on plots of the dependent variable against the independent variable as well as plots of residuals against fitted values. Neither nonlinear association (such as U shape association) nor a structured pattern of residual distribution was evident (data available on request).
Causal estimates from Mendelian randomisation analysis
Causal effects of alcohol intake on cardiovascular health and life style factors were inferred by IV estimation techniques (Table 5 and Supplementary Table S2). Corresponding causal relationships were also assessed through the association of the rs671 genotype with cardiovascular health and life style factors (Supplementary Tables S3 and S4). In men, alcohol intake, instrumented by the rs671 genotype, was associated with higher risks of hypertension, blood pressure, waist circumference, waist to hip ratio, log-transformed fasting blood glucose, HDL cholesterol and log-transformed triglycerides as well as with lower LDL cholesterol (all p < 0.05). In women, there was little evidence for causal influences of alcohol intake on cardiovascular outcomes with an exception of hip circumference (p = 0.035). The heterogeneity of IV estimates in men and women was observed for hip circumference (p = 0.038) under the fixed effect model.
Population-level causal effects were assessed as IV estimates where interaction of the rs671 genotype and sex was used as an IV given that alcohol intake was stratified by sex as well in the whole population (Table 6). As a result, one unit of alcohol intake (g/day) was associated with higher hypertension risks, blood pressure, waist to hip ratio, log-transformed fasting blood glucose, HDL cholesterol, log-transformed triglycerides as well as with lower LDL cholesterol at a population level.
Discussion
Here, we present a Mendelian randomisation study on alcohol intake and cardiovascular outcomes by analysing 7,152 individuals (3,365 men and 3,787 women) in South Korea. Causal influences of the exposure cannot be properly measured if the exposure level (alcohol intake, in this study) is indistinguishably low although there is a potentially valid IV (the rs671 genotype, in this study). For this reason, potential health outcomes consequent on alcohol drinking are not easily assessed in Asian women compared to Asian men5,21. We first replicated null or weakly observed association of the rs671 genotype and cardiovascular outcomes in women. Furthermore, we ensured such null association in women was not because of any female-specific biological mechanism but because of low drinking levels, by demonstrating analogous null association in male never-drinkers. The average alcohol intake level was 18.8 g/day in men and 1.3 g/day in women and 22.9 g/day and 0.0 g/day in male ever- and never-drinkers.
We quantified influences of alcohol intake on a wide range of cardiovascular outcomes by using instrumental variable estimation techniques. In men, one unit of alcohol intake (g/day), explained by the rs671 genotype, was associated with higher hypertension risks and higher level of systolic blood pressure, diastolic blood pressure, waist circumference, fasting blood glucose, HDL cholesterol, triglycerides and with lower LDL cholesterol. In women, none of these associations were observed as expected due to a very low alcohol intake. In the whole population, alcohol intake instrumented by interaction of the rs671 genotype and sex, appeared to have the same effects on cardiovascular outcomes as in the male population, although the confidence intervals of the effect sizes were larger.
Overall, we showed that alcohol intake is detrimental to most cardiovascular outcomes in the general Asian population as shown previously in Asian male populations11,31. The exception is high HDL cholesterol and low LDL cholesterol as they are generally considered favourable risk profiles with respect to cardiovascular health, although the protective role of high HDL cholesterol may not be fully established compared to that of low LDL cholesterol which has been supported by a number of Mendelian randomisation and RCT studies1,27,28,32.
The credibility of the rs671 genotype in ALDH2 as an IV for alcohol intake has been discussed in many studies11,12,14,20,21. Biochemically, ALDH2 encodes the main enzyme in alcohol metabolism transferring toxic acetaldehyde, into non-toxic acetate. Simultaneously, it prevents another toxic chemical, aldehyde, from accumulating in the body. People carrying a mutated allele of this gene that produces an inactive form of the ALDH2 enzyme (which is the case mainly in Asians), experience discomfort after drinking such as facial flushing, nausea and a rapid heartbeat. This is likely to be underlying reason for the association between ALDH2 genotype and drinking behaviour.
In our data, supporting evidence was found that the rs671 genotype in ALDH2 satisfied three core assumptions for an IV. First, it was independent of known confounders including age, education, residential area, physical activities and smoking status, both in men and women, as expected. Potential residual confounding by population stratification might exist to some extent, but we adjusted for genotypic principal components as covariates in the instrumental variable model. Second, it was strongly associated with alcohol intake (g/day) (F-statistic = 262 in men and 38 even in women) confirming that it was an adequate IV unlikely to suffer weak instrument bias in this study. Furthermore, the rs671 genotype was also associated with other directly relevant alcohol-related traits. That is, people with slow alcohol metabolism due to carriage of a mutated allele (rs671 A-allele) appeared to have lower proportion of ever and current drinkers as well as lower levels of GGT. GGT is often used as a biomarker for heavy drinking and the lower levels of GGT are likely to be influenced by drinking less alcohol as suggested in the latest study33. The third assumption required for instrumental variable analysis (that the ALDH2 genotype influences cardiovascular outcomes only through alcohol intake, in this study) is (like the assumption of no unmeasured confounding) impossible to validate. However, in this study, null effects in women provided evidence that it was unlikely that the estimated causal effects were due to pleiotropic effects; if there were pleiotropic effects, causal effects would have been observed in women as well as in men, as argued in detail elsewhere5,21.
Several studies have previously reported the causal relationship between alcohol intake and cardiovascular outcomes10,11,20,21,22,23. One of main strengths of the current study lies in relatively accurate estimation of population-level causal effects of alcohol intake when the alcohol intake is stratified by gender. Instead of the genotype alone, we formally used interaction of the genotype and sex as an IV for alcohol for the first time to our knowledge. Secondly, we considered a broad spectrum of cardiovascular outcomes compared to previously studies in an Asian population. For example, Chen et al. reported a sex-specific causal effect of alcohol intake on systolic and diastolic blood pressures and hypertension21 in their meta-analysis based on results extracted from published data, whereas we estimated both sex-specific and population-level causal effects on 12 additional outcomes in as large as or larger samples in individual level data. In another previous study, Kato et al. reported a strong sex-specific association of the rs671 genotype and blood pressures22 and also showed that such association was mediated by alcohol intake implying a causal relationship between alcohol intake and blood pressures, but their approach was limited in terms of quantification of the causal effects compared to the IV analysis that we applied in the current study. Finally it should be also mentioned that in a recent paper, Holmes et al. extensively covered the causal relationship of alcohol intake with various cardiovascular events and risk factors in the largest samples to date10; however, they used a different IV, the rs1229984 genotype in ADH1B which is known to be a much weaker IV than the rs671 genotype in ALDH2 we used, as the latter is not polymorphic in European individuals of their study. Therefore, our current study is one of the most comprehensive study providing robust causal effects of alcohol intake on cardiovascular health outcomes. Another interesting feature of this study may be that it is one of the first Mendelian randomisation studies quantifying causal effects in alcohol intake in the Korean population, although there exists a relevant observational study34. The Korean population was selected, not only because it is an Asian population carrying a mutated rs671-A allele in ALDH2, but also because the population level alcohol intake is 71% and 84% higher than Japan and China, respectively, ranking it as the country with the highest level of heavy drinking in Asia (based on the alcohol per capita consumption on average between 2008 and 2010 in the 2014 WHO report). Our results were, however, consistent with those in other instrumental variable based studies in Japan21 and China11.
Nevertheless, our study has limitations. One of main limitation is the use of imputed genotype ALDH2 rs671 although the imputed genotype was generated by a standardised protocol. Genotypes were quality controlled prior to imputation (based on missing call rates, minor allele frequency, Hardy-Weinberg equilibrium and sex match) and publicly available reference datasets were used with commonly used and previously evaluated software35,36. In addition, imputed genotypes were evaluated based on imputation quality score and Hardy-Weinberg equilibrium test. Thus, our imputed genotype would be as informative as a directly measured genotype, as shown in numerous genome-wide association studies. Also we acknowledge the limitation of the stratified analysis, as the stratification of the male population on drinking behaviour history (ever vs. never drinkers) could introduce collider bias30. We identified a risk factor susceptible to collider bias in our data and adjusted for its effect on associations between genotype and cardiovascular outcomes, which produced little change in the effect estimates.
Despite providing evidence for a causal link between alcohol intake and a range of cardiovascular traits, our study did not observe clear causal effects of alcohol intake on cardiovascular disease or body mass index. This is consistent with previous evidence, such as that provided by Au Yeung et al. who reported a null effect of alcohol intake on cardiovascular disease in Chinese men11, although a recent meta-analysis by Holmes et al.10 reported strong effects on both in individuals of European descent. One possible explanation is that our study and the study by Au Yeung et al.11 were underpowered to detect the causal effect as the samples sizes were much smaller than those accrued by Holmes et al. (7,152 and 4,500 compared with 260,000, respectively)10. However, it is not straightforward to draw such conclusion yet because these studies have not only different sample sizes, but also different instruments (ALDH2 genotype being a stronger instrument than ADH1B genotype), different ethnic backgrounds (Korean and Chinese compared with European) and different methods were used to define cardiovascular disease (self- report and self- report compared with combination of self- report, medical records, clinical/lab measures, death certificate and ICD code). Thus, a carefully designed large-scale study in well-phenotyped Asian population would be needed to further investigate this discrepancy.
In conclusion, this study indicates that a reduction in alcohol intake may be beneficial to cardiovascular health through avoiding detrimental influences on cardiovascular risk factors.
Methods
Study participants
Subjects for the analysis were obtained from two population based studies within the Korean Genome and Epidemiology Study (KoGES), the rural Ansung and urban Ansan cohorts. Detailed information for each study has been described elsewhere37. Briefly, the Ansung-Ansan cohorts were designed as longitudinal prospective studies initiated in 2001 and adopted the same investigational method. Participants in each cohort (5,018 in Ansung and 5,020 in Ansan aged 39–70) were recruited using a two-stage cluster sampling method. All participants took part in a health examination, interviews and laboratory tests. The current study was based on the baseline data collected in 2001 from a total of 7,152 participants having the rs671 genotype in ALDH2 available. All participants provided informed consent which was approved by the Human Subjects Review Committee at the Korea University Ansan Hospital or the Ajou University Medical Centre. The current study was approved by the Institute Review Board at the Korea University (KU-IRB-14-EX-153-A-1).
Basic characteristics
Information was collected on demographic characteristics including age, area, education, physical activity and current smoking status. Education level was divided into four groups: elementary school, middle school, high school, or university. Physical activity was divided into two groups: practice or do not practice, according to whether or not the individual participated in any of the following daily activity types; intense physical activity at least 20 minutes, moderate physical activity at least 30 minutes, or walking at least 30 minutes. Current smokers were defined as a person who smoked cigarettes regularly at the time of the survey.
Alcohol traits
Participants were also asked about their lifetime drinking behaviour, current drinking behaviour and detailed drinking behaviour over the previous 30 days, including frequency, amount and type of alcoholic beverages. Using this information along with average alcohol content of each beverage, total alcohol intake (g/day) was calculated. Further information on alcohol intake can be found in a previous publication38. As well as total alcohol intake (g/day), current drinking status, alcohol intake in current drinkers and GGT were considered as alcohol-related traits in this study. A current drinker was defined as an individual who drank alcoholic beverages regularly at the time of the survey. GGT concentration (IU/L) was measured from blood samples in the Seoul Clinical Laboratories (Seoul, Republic of Korea) collected after at least 8 hours of fasting.
Blood pressure and other risk factors
Blood pressure was measured in a sitting position with a mercury sphygmomanometers after at least 5 minutes of rest. Two acceptable measurements of blood pressure were obtained within a 1 minute interval and recorded to the nearest 2 mmHg. Average measurements for systolic and diastolic blood pressure were used for statistical analysis. Height (cm) and body weight (kg) were measured to the nearest 0.1 cm or 0.1 kg without shoes, from which BMI (kg/m2) was derived. Waist circumference (cm) was measured at the narrowest part between the lower rib and the iliac crest to the nearest 0.1 cm and the average of 3 repeated measurements was calculated. Hip circumference was measured at the widest portion of the buttocks to the nearest 0.1 cm and the average of 3 repeated measurements was calculated. Waist to hip ratio was derived from waist circumference and hip circumference.
For laboratory tests, all participants had at least an 8 hour fasting period before blood collection. Collected blood samples were analysed in the Seoul Clinical Laboratories (Seoul, Republic of Korea) for assays including fasting blood glucose (mg/dL), total cholesterol (mg/dL), HDL cholesterol (mg/dL) and triglycerides (mg/dL). LDL cholesterol (mg/dL) was derived using the Friedewald formula39 in subjects with triglycerides less than 400 mg/dL as follows; LDL cholesterol = total cholesterol– HDL cholesterol– (triglycerides/5.0). For subjects with triglycerides of 400 mg/dL or more, LDL cholesterol value was marked as missing. GGT concentration (IU/L) was measured from the same blood samples.
Disease outcome
According to health interview and examination, participants with self-reported diagnosed hypertension, use of blood pressure medicine, or measured systolic blood pressure greater than 140 mmHg, or diastolic blood pressure greater than 90 mmHg were considered as hypertensive. Cardiovascular disease status was defined by doctor-diagnosed and self-reported questionnaire information on myocardial infarction, congestive heart failure, coronary artery disease, peripheral blood vessel disease and cerebrovascular disease. A coronary heart disease event was additionally defined by the same questionnaire information but only on myocardial infarction and coronary artery disease. Diabetes was defined by doctor-diagnosed and self-reported questionnaire information.
Genotyping quality control and imputation
Detailed information is provided elsewhere37. Briefly, DNA samples were isolated from the peripheral blood of participants and genotyped using the Affymetrix Genome-Wide Human SNP array 5.0 (Affymetrix, Inc., Santa Clara, CA, USA). The accuracy of the genotyping was calculated by Bayesian Robust Linear Modelling using the Mahalnobis Distance genotyping algorithm40. A total of 352,228 SNPs in 8,842 participants became available after pre-imputation QC, 1) excluding SNPs with high missing genotype call rates (> 5%), with minor allele frequency (MAF) < 0.01 and not in Hardy-Weinberg equilibrium (HWE, P value < 1 × 10−6) and 2) removing samples with sex mismatch. Genetic principal components were computed in a subset of 304,225 SNPs after excluding additional 48,003 SNPs (not in HWE under a more conservative criterion, P value < 1 × 10−5) through the EIGENSTRAT software package41.
To impute rs671 ALDH2 genotype (http://www.ncbi.nlm.nih.gov/projects/SNP/), all genotypes in chromosome 12 were imputed using the 1000 Genomes Phase 1 v3 reference panel. The reference datasets of all populations were downloaded from the IMPUTE2 website35. To minimise the computational intensity and increase efficiency, genotypes were pre-phased with SHAPEIT36 prior to imputation by IMPUTE235 with the default options. As a post-imputation QC, SNPs were removed if MAF was low (<0.05) or the imputation info value was low (<0.8). As a result, the rs671 genotype in ALDH2 became available in a total of 7,152 participants with expected MAF of 0.194 and imputation info value of 0.845.
Statistical analysis
Statistical analyses were performed using Stata SE 12.0 (Stata Corp, Carollina, USA). First, the distribution of variable values was investigated. Fasting blood glucose, GGT and triglycerides were log-transformed to mimic a Gaussian distribution. No outliers were detected by visual inspection. Descriptive statistics of all variable values were presented as mean ± standard error for a continuous variable and as number of counts and percentage for a categorical variable in men and women separately, according to their rs671 genotype in ALDH2. Apart from major dependent variables (e.g. hypertension) and major independent variables (e.g. alcohol intake), some variables included missing data points. Mean difference of these variables in men and women was evaluated through Student’s t-test for a continuous variable and by chi-squared test for a categorical variable. Similarly, mean differences in these variables among three different rs671 genotype groups were compared using one-way analysis of variance (ANOVA) for a continuous variable and using chi-squared test for a categorical variable; in men and women and in male ever-drinkers and male never-drinkers, separately. In addition, in order to evaluate potential collider stratification bias, the difference of variable distribution by genotype between male ever-drinkers and male never-drinkers was tested using generalized regression models which include rs671 genotype, drinking behaviour history (never vs. ever) and interaction of genotype and drinking behaviour history as the independent variables.
The association between alcohol intake and other variables was assessed under an OLS regression model in men and women, separately. Continuous risk factors were predicted by alcohol intake under a linear regression model adjusting for potential confounding factors such as age, area, education, physical activity and smoking status. Hypertension, cardiovascular disease, coronary heart disease and diabetes were also predicted by alcohol intake under a logistic regression model adjusting for the same potential confounding factors. In order to investigate the potential violation of assumptions such as linearity of association and normality of the error distribution, plots of dependent variables against independent variables as well as plots of residuals against fitted values were generated. Results are presented as estimated regression coefficients β with 95% confidence interval (CI) for a continuous variable and estimated odds ratio (OR) with 95% CI for a categorical variable. Corresponding p-values are also provided. The difference of estimates between men and women was assessed by Cochran’s Q test using fixed effect models assuming the true effect of alcohol is the same in men and women.
Lastly, the causal effect of alcohol intake on other variables was measured under an IV regression with a two stage least squares estimation method in men and women, separately, using the rs671 genotype as an instrument. For continuous risk factors, a two stage linear model was performed, with adjustments for age, area, education, physical activity and smoking status as well as additional adjustments for genotypic principal components to take into account population structure for the rs671 genotype. For hypertension and cardiovascular disease, a two stage logistic model was conducted; in the first stage, alcohol intake was predicted by rs671 genotype (with additive effect) under a linear regression model (adjusted for age, area, education, physical activity, smoking status and principal components); in the second stage, disease outcome was predicted by fitting the alcohol intake value from the first stage, under a logistic regression model (adjusted for the same potential confounding factors). Results were shown by providing estimated regression coefficients β with 95% CI for a continuous variable and estimated OR, with 95% CI for a categorical variable along with corresponding p-values. The difference of estimates between men and women was assessed by Cochran’s Q test using fixed effect models assuming the true effect of alcohol is the same in men and women.
The same causal effect was then quantified in the whole population, using interaction of the rs671 genotype and sex as an instrument. Instrumental variable regression models were additionally adjusted for the rs671 genotype and sex, the variables that were used to compute the interaction. It should be noted that interaction of the rs671 genotype and sex was used as an instrument even if the rs671 genotype was directly included in the model42.
Additional Information
How to cite this article: Cho, Y. et al. Alcohol intake and cardiovascular risk factors: A Mendelian randomisation study. Sci. Rep. 5, 18422; doi: 10.1038/srep18422 (2015).
Change history
27 May 2016
A correction has been published and is appended to both the HTML and PDF versions of this paper. The error has not been fixed in the paper.
References
Marmot, M. G. Alcohol and coronary heart disease. Int J Epidemiol 13, 160–7 (1984).
Bovet, P. & Paccaud, F. Commentary: alcohol, coronary heart disease and public health: which evidence-based policy. Int J Epidemiol 30, 734–7 (2001).
Ronksley, P. E., Brien, S. E., Turner, B. J., Mukamal, K. J. & Ghali, W. A. Association of alcohol consumption with selected cardiovascular disease outcomes: a systematic review and meta-analysis. BMJ 342, d671 (2011).
Freiberg, M. S. & Samet, J. H. Alcohol and coronary heart disease: the answer awaits a randomized controlled trial. Circulation 112, 1379–81 (2005).
Davey Smith, G. Use of genetic markers and gene-diet interactions for interrogating population-level causal influences of diet on health. Genes Nutr 6, 27–43 (2011).
Richmond, R. C., Al-Amin, A., Davey Smith, G. & Relton, C. L. Approaches for drawing causal inferences from epidemiological birth cohorts: a review. Early Hum Dev 90, 769–80 (2014).
Davey Smith, G. & Ebrahim, S. 'Mendelian randomization': can genetic epidemiology contribute to understanding environmental determinants of disease? Int J Epidemiol 32, 1–22 (2003).
Davey Smith, G. & Ebrahim, S. What can mendelian randomisation tell us about modifiable behavioural and environmental exposures? BMJ 330, 1076–9 (2005).
Davey Smith, G. & Hemani, G. Mendelian randomization: genetic anchors for causal inference in epidemiological studies. Hum Mol Genet 23, R89–98 (2014).
Holmes, M. V. et al. Association between alcohol and cardiovascular disease: Mendelian randomisation analysis based on individual participant data. BMJ 349, g4164 (2014).
Au Yeung, S. L. et al. Moderate alcohol use and cardiovascular disease from Mendelian randomization. PLoS One 8, e68054 (2013).
Taylor, A. E. et al. Exploring causal associations of alcohol with cardiovascular and metabolic risk factors in a Chinese population using Mendelian randomization analysis. Sci Rep 5, 14005 (2015).
Davey Smith, G. & Ebrahim, S. Mendelian randomization: prospects, potentials and limitations. Int J Epidemiol 33, 30–42 (2004).
Au Yeung, S. L. et al. Is aldehyde dehydrogenase 2 a credible genetic instrument for alcohol use in Mendelian randomization analysis in Southern Chinese men? Int J Epidemiol 42, 318–28 (2013).
Okamoto, K., Murawaki, Y., Yuasa & Kawasaki, H. Effect of ALDH2 and CYP2E1 gene polymorphisms on drinking behavior and alcoholic liver disease in Japanese male workers. Alcohol Clin Exp Res 25, 19S–23S (2001).
Eng, M. Y., Luczak, S. E. & Wall, T. L. ALDH2, ADH1B and ADH1C genotypes in Asians: a literature review. Alcohol Res Health 30, 22–7 (2007).
Hendershot, C. S. et al. Associations of ALDH2 and ADH1B genotypes with alcohol-related phenotypes in Asian young adults. Alcohol Clin Exp Res 33, 839–47 (2009).
Tsuchihashi-Makaya, M. et al. Gene-environmental interaction regarding alcohol-metabolizing enzymes in the Japanese general population. Hypertens Res 32, 207–13 (2009).
Takeuchi, F. et al. Confirmation of ALDH2 as a Major locus of drinking behavior and of its variants regulating multiple metabolic phenotypes in a Japanese population. Circ J 75, 911–8 (2011).
Lewis, S. J. & Davey Smith, G. Alcohol, ALDH2 and esophageal cancer: a meta-analysis which illustrates the potentials and limitations of a Mendelian randomization approach. Cancer Epidemiol Biomarkers Prev 14, 1967–71 (2005).
Chen, L., Davey Smith, G., Harbord, R. M. & Lewis, S. J. Alcohol intake and blood pressure: a systematic review implementing a Mendelian randomization approach. PLoS Med 5, e52 (2008).
Kato, N. et al. Meta-analysis of genome-wide association studies identifies common variants associated with blood pressure variation in east Asians. Nat Genet 43, 531–8 (2011).
Shin, C. M. et al. Association between alcohol intake and risk for gastric cancer with regard to ALDH2 genotype in the Korean population. Int J Epidemiol 40, 1047–55 (2011).
Zhang, S. Y. et al. Meta-analysis of association between ALDH2 rs671 polymorphism and essential hypertension in Asian populations. Herz 40, 203–208 (2015).
Guo, Y. J. et al. The ALDH2 Glu504Lys polymorphism is associated with coronary artery disease in Han Chinese: Relation with endothelial ADMA levels. Atherosclerosis 211, 545–50 (2010).
Mizuno, Y. et al. East Asian Variant of Aldehyde Dehydrogenase 2 (ALDH2*2) is Associated with Coronary Spastic Angina: Possible Roles of Reactive Aldehydes and Implications of Alcohol Flushing Syndrome. Circulation (2015).
Do, R. et al. Common variants associated with plasma triglycerides and risk for coronary artery disease. Nat Genet 45, 1345–52 (2013).
Burgess, S., Freitag, D. F., Khan, H., Gorman, D. N. & Thompson, S. G. Using multivariable Mendelian randomization to disentangle the causal effects of lipid fractions. PLoS One 9, e108891 (2014).
Slichter, D. Testing Instrument Validity and Identification with Invalid Instruments. Technical report, Department of Economics, University of Rochester Rochester, USA (2014).
Cole, S. R. et al. Illustrating bias due to conditioning on a collider. Int J Epidemiol 39, 417–20 (2010).
Davey Smith, G., Timpson, N. & Ebrahim, S. Strengthening causal inference in cardiovascular epidemiology through Mendelian randomization. Annals of Medicine 40, 524–541 (2008).
Briel, M. et al. Association between change in high density lipoprotein cholesterol and cardiovascular disease morbidity and mortality: systematic review and meta-regression analysis. BMJ 338, b92 (2009).
Xu, L. et al. Alcohol Use and Gamma-Glutamyltransferase Using a Mendelian Randomization Design in the Guangzhou Biobank Cohort Study. PLoS One 10, e0137790 (2015).
Shin, C., Kwack, K., Cho, N. H., Kim, S. H. & Baik, I. Sex-specific differences in the association of a common aldehyde dehydrogenase 2 gene polymorphism and alcohol consumption with stroke risk in a Korean population: a prospective cohort study. Nutr Res Pract 9, 79–86 (2015).
Howie, B. N., Donnelly, P. & Marchini, J. A flexible and accurate genotype imputation method for the next generation of genome-wide association studies. PLoS Genet 5, e1000529 (2009).
Delaneau, O., Zagury, J. F. & Marchini, J. Improved whole-chromosome phasing for disease and population genetic studies. Nat Methods 10, 5–6 (2013).
Cho, Y. S. et al. A large-scale genome-wide association study of Asian populations uncovers genetic factors influencing eight quantitative traits. Nat Genet 41, 527–34 (2009).
Baik, I. & Shin, C. Prospective study of alcohol consumption and metabolic syndrome. Am J Clin Nutr 87, 1455–63 (2008).
Friedewald, W. T., Levy, R. I. & Fredrickson, D. S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin Chem 18, 499–502 (1972).
Rabbee, N. & Speed, T. P. A genotype calling algorithm for affymetrix SNP arrays. Bioinformatics 22, 7–12 (2006).
Price, A. L. et al. Principal components analysis corrects for stratification in genome-wide association studies. Nat Genet 38, 904–9 (2006).
Card, D. Using Geographic Variation in College Proximity to Estimate the Return to Schooling. in Aspects of labour market behaviour: essays in honour of John Vanderkamp (eds. Vanderkamp, J., Christofides, L. N., Grant, E. K. & Swidinsky, R. ) (University of Toronto Press, Toronto; Buffalo, 1993).
Acknowledgements
This study was provided with biospecimens and data from the Korean Genome Analysis Project (4845-301), the Korean Genome and Epidemiology Study (4851-302) and Korea Biobank Project (4851-307, KBP-2014-062) that were supported by the Korea Center for Disease Control and Prevention, Republic of Korea. This research was supported by BAsic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT and future Planning (2015R1A2A1A15054758). SYS, CLR and GDS are supported by the Medical Research Council Integrative Epidemiology Unit. SYS is supported by a Post-Doctoral Research Fellowship from the Oak Foundation.
Author information
Authors and Affiliations
Contributions
C.L.R., G.D.S. and M.S. conceived the study. M.S. acquired the data. S.Y.S. and G.D.S. developed the statistical analysis plan. Y.C., S.W. and M.S. analysed the data. S.Y.S. and M.S. prepared the first draft of manuscript. S.Y.S., Y.C., C.L.R., G.D.S. and M.S. contributed to the writing of the manuscript. All authors reviewed and agreed on the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Cho, Y., Shin, SY., Won, S. et al. Alcohol intake and cardiovascular risk factors: A Mendelian randomisation study. Sci Rep 5, 18422 (2015). https://doi.org/10.1038/srep18422
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep18422
This article is cited by
-
Assessment of cognitive function and sleep–wake rhythms in community-dwelling older adults
Sleep and Biological Rhythms (2024)
-
Moderate alcohol consumption on the risk of stroke in the Million Veteran Program
BMC Public Health (2023)
-
Effects of long sleep time and irregular sleep–wake rhythm on cognitive function in older people
Scientific Reports (2021)
-
The effect of ALDH2 rs671 gene mutation on clustering of cardiovascular risk factors in a big data study of Chinese population: associations differ between the sexes
BMC Cardiovascular Disorders (2020)
-
Exploiting horizontal pleiotropy to search for causal pathways within a Mendelian randomization framework
Nature Communications (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
