
Abstract
Fossil records indicate that the genus Pinus L. split into two subgenera by the Late Cretaceous, although subgenus Strobus (D. Don) Lemmon is less well documented than subgenus Pinus L., especially in eastern Asia. In this paper, Pinus maomingensis sp. nov. is established based on a compressed seed cone from the upper Eocene of the Maoming Basin of southern China. This species is attributed to genus Pinus, subgenus Strobus, section Quinquefoliae Duhamel, subsection Strobus Loudon based on the combination of morphological characters obtained from the cone scales, specifically from the terminal umbo, rhombic apophysis and cuticle structure. Associated fascicles of needle leaves with deciduous sheaths and bulbous bases are recognized as Pinus sp. and also represent Pinus subgenus Strobus. This new discovery from the Maoming Basin constitutes the first megafossil record of subgenus Strobus from southern China and implies that the members of this subgenus arrived in the southern region of China by the late Eocene. The extant species of subgenus Strobus are mainly distributed in northern temperate and tropical to subtropical mountainous regions. We propose that the Maoming Basin was adjacent to a mountainous region during the late Eocene.
Similar content being viewed by others


Introduction
Pinus L., consisting of more than 110 extant species, is the largest and the most widespread genus of Pinaceae in the Northern Hemisphere1,2,3. This genus is subdivided into subgenus Pinus L. (subgenus Diploxylon (Koehne) Pilger, the hard pines) and subgenus Strobus (D. Don) Lemmon (subgenus Haploxylon (Koehne) Rehder, the white or soft pines)3,4,5,6. The major differences between the above two subgenera are the number of fibrovascular bundles per needle, sheath persistence and the position of the umbo on ovuliferous scales. Their general distinguishing criteria are listed in Table 1.
The origin of the genus Pinus is thought to date to the Early Cretaceous7. Ryberg et al.8 suggest that some species of the fossil genus, Pityostrobus, might be reassigned to genus Pinus and the evolutionary diversification of Pinaceae began earlier than previously recognized from fossil evidence. Currently the oldest fossil record of this genus is a seed cone P. yorkshirensis Ryberg, Stockey, Hilton, Mapes, Riding et Rothwell8. It was discovered in the Early Cretaceous Wealden Formation (Fm.) of Yorkshire, United Kingdom and was placed in subgenus Pinus based on morphological and anatomical structure. Although pine fossils are well documented in a variety of stratigraphic and geographic settings9, fossil records of the two subgenera of Pinus differ greatly in their past abundances. Most of these fossils have affinity to subgenus Pinus and relatively few have been confidently placed in subgenus Strobus2,10. Cretaceous fossil needles of Pinus sp.11, Pinus yezoensis12 and a fossil cone of Pinus magothensis13 were initially accepted as members of subgenus Strobus. However, they were later thought to have affinity with subgenus Pinus or possibly with other pinaceous genera14,15. Presently, the rise of subgenus Strobus can be dated confidently to the permineralized wood, Pinuxylon sp., from the Late Cretaceous Aachen Fm. of northeastern Belgium16. More recent records of subgenus Strobus, for example ovulate cones, occur in the mid-Eocene strata10, no megafossils are known from the Paleocene7.
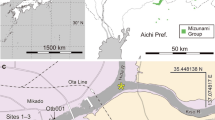
The megafossil record of subgenus Strobus in China is very restricted. Only two Pliocene fossil wood species from Yunnan Province, Pinus armandii Franchet and P. cf. armandii, were reported by Yi et al.17,18. In this study, we describe two species of subgenus Strobus based on a fossil pine cone and needle remains collected from the Huangniuling Fm. of the Maoming Basin in southern China (21°42′ N, 110°53′E) (Fig. 1).

Geographic map of the Maoming Basin, Guangdong Province, China and stratigraphic column of the fossil locality.
(A) location of the Maoming Basin (red star) (drawn by Q.X., using Adobe Photoshop CS5). (B) Stratigraphic column of Huangniuling Formation, modified from Aleksandrova et al.22. The fossil cone and the single isolated ovuliferous scale were collected from layer 14 (red arrow) and the needle fossils were collected from layer 18 (blue arrow).
The Maoming Basin is a small, upper Mesozoic to Cenozoic sedimentary basin oriented along a northwestern to southeastern axis in Guangdong Province. The Upper Cretaceous to Neogene deposits of the Maoming Basin are subdivided into eight formations; in ascending order they are: the Sanyajiang Fm., Tongguling Fm., Shangdong Fm., Youganwo Fm., Huangniuling Fm., Shangcun Fm., Laohuling Fm. and Gaopengling Fm19,20. The Huangniuling Fm., from which the megafossils were recovered, consists principally of fluvial grey, yellow to white sandstones, siltstones and conglomerates with beds and lenses of varicolored mudstones and claystones, ranging from whitish pink to grayish green to brownish gray. Based on a magnetostratigraphic study by Wang et al.21, buttressed by palynological data by Aleksandrova et al.22, the age of the Huangniuling Fm. is considered to be late Eocene. Therefore, the occurrences of fossil pine cone and needle remains from Huangniuling Fm. provide new information on the distribution of subgenus Strobus and imply that the members of this subgenus arrived in the southern region of China by the late Eocene.
Results
Systematics.
The fossil cone.
Family: Pinaceae Lindley, 1836
Genus: Pinus Linnaeus, 1753
Subgenus: Strobus (D. Don) Lemmon, 1983
Section: Quinquefoliae Duhamel, 1755
Subsection: Strobus Loudon, 1838
Species: Pinus maomingensis Xu, Jin, Zhou, Kodrul et Naugolnykh sp. nov.
Etymology
The species name is derived from the Maoming Basin where the specimens were collected.
Holotype
MMJ2-1-005a (Fig. 2A) and MMJ2-1-005b (Fig. 2B), part and counterpart of a seed cone, which has seeds inside.

Pinus maomingensis sp. nov. (A,B) Holotype; part (A, MMJ2-1-005a) and counterpart (B, MMJ2-1-005b) of the cone with ovuliferous scales, arrowhead in (A) points to the protrusion. (C) Rhombic apophysis from (B) arrowhead points to the reflexed ridge. (D) Upper part of apophysis from (B) showing details of terminal umbo. (E) Paratype (MMJ2-1-006); single ovuliferous scale, arrowhead points to the protrusion. (F) The protrusion (arrowhead) in (A). (G–I) Seeds show little seed coat, arrowhead in figure (G) shows the position of the seed in the cone. Scale bars: A–C,E,G = 1 cm; D,F,H,I = 0.5 cm.
Paratype
MMJ2-1-006 (Fig. 2E), an isolated ovuliferous scale.
Repository
The Museum of Biology of Sun Yat-sen University, Guangzhou, Guangdong Province, China.
Type locality and horizon
Huangniuling Fm., upper Eocene. Specimens were collected in Jintang Town, Maoming City, Guangdong Province (Fig. 1).
Diagnosis
Seed cone oblong-elliptical to cylindrical. Ovuliferous scales thin with longitudinal ridges on abaxial side, spirally arranged. Apophyses rhomboidal to broadly rhomboidal; no sealing band. Umbo terminal, slightly sunken, bearing a slightly swelling transverse ridge in a diamond-shaped area.
Description
The seed cone (Fig. 2A,B) is variably oblong to elliptical or cylindrical. The visible portion of the seed cone is 16 cm in length. The cone width, generally consistent over the middle and upper part of the cone, is ca. 5 cm. The basal part of the cone is more or less cuneate (probably due to incomplete preservation) and ca. 1 cm in diameter. The ovuliferous scales are thin, oblong-obovate, helically arranged around the axis, about 5 cm long, 2.4 cm wide and about 0.1 cm thick in the middle, with a slightly expanded apophysis (Fig. 2D). The isolated ovuliferous scale (Fig. 2E) is rhomboidal to obovate, sessile, ca. 2.6 cm long by 2 cm wide. Apophyses are rhomboidal to broadly rhombic. Longitudinal ridges and furrows are clearly visible on the abaxial side of the scales. The umbones (Fig. 2A,D,E) are terminal and slightly sunken. A small protuberance (Fig. 2A,F) occurs in the umbonal area, also present in the isolated ovuliferous scale (Fig. 2E). A pair of arched ridges (Fig. 2C,D) near the umbo stretches transversely, gradually becomes robust and slightly recurved, then merges with upper lateral margin of the apophysis. Seeds (Fig. 2H,I) are elliptical to fusiform in shape, ca. 6 mm long by 3 mm wide.
Three types of epidermal cells (Fig. 3) were retrieved from the upper part of the abaxial surface of ovuliferous scale: (i) longitudinally elongate cells (Fig. 3C–E,H); (ii) irregular polygonal cells (Fig. 3C,D,H,I); and (iii) irregularly shaped cells (Fig. 3A,F). Anticlinal walls of all epidermal cells are well developed and express slightly undulatory outlines. In longitudinally elongate cells, the end walls are either transverse or oblique to the side walls. The trichome bases (Fig. 3C,G) are composed of two or three rings of small rectangular epidermal cells and unicellular trichomes (Fig. 3C) are sparsely dispersed on these cuticles. No stomatal complexes were observed.

Cuticle of Pinus maomingensis sp. nov. obtained from the apophysis. (A) The cells from the ridge area (green arrowhead); pink arrowhead shows the nearby irregularly shaped cells. (B) Enlarged from (A), the green arrowhead shows the ridge. (C,D) The longitudinally elongate cells (blue arrowhead) and irregularly polygonal cells (red arrowhead) from the ribs and furrows, yellow arrowhead in (C) shows the unicellular trichome and white arrowhead in (C) shows the trichome base. (E) The longitudinally elongate cells, enlarged from (D). (F) The irregularly shaped cells. (G) Trichome base. (H) Cuticle shows the longitudinally elongate cells (blue arrowhead) and irregularly polygonal cells (red arrowhead). (I) Irregularly polygonal cells. Scale bar = 50 μm.
The fossil needles.
Family: Pinaceae Lindley, 1836
Genus: Pinus Linnaeus, 1753
Subgenus: Strobus (D. Don) Lemmon, 1983
Species: Pinus sp.
Referred specimens
MMJ3-002a, MMJ3-002b, MMJ3-003, MMJ3-037 to MMJ3-049, MMJ3-094 to MMJ3-097, fossil needles.
Repository
The Museum of Biology of Sun Yat-sen University, Guangzhou, Guangdong Province, China.
Type locality and horizon
Huangniuling Fm., upper Eocene. Specimens were collected in Jintang Town, Maoming City, Guangdong Province (Fig. 1).
Description
The short shoot leaves are in fascicles with deciduous basal sheaths. The number of needles per fascicle is mostly five but varies from 3 to 5 (Fig. 4A–G). The needles are up to 10 cm long by ca. 1 mm wide, with triangular in cross section (Fig. 4C) and finely serrate margins (Fig. 4D). Needle widths are uniform along their entire length. The fascicles have bulbous bases (Fig. 4B,E,F,H). Membranous sheaths are deciduous at maturity, but they are retained (Fig. 4H) in the young fascicles.

Leaves of Pinus sp.
(A) Showing the preservation of needles (MMJ3-094). (B) Fascicle bearing five needles, showing bulbous base (arrowhead) (MMJ3-038). (C) Longitudinal furrow (arrowheads) of the needle, enlarged from (B). (D) Finely serrate margins (arrowheads), enlarged from (B). (E) Fascicle bearing three needles, showing bracts (arrowhead) and leaf scars (MMJ3-095). (F) Needle fascicle bears four needles (MMJ3-096). (G) Fascicle from which cuticle fragments were obtained bears four needles (MMJ3-002-a). (H) Bulbous base (MMJ3-097), arrowhead points to a pair of sheath scales. (I,J) Cuticle fragments obtained from (G), showing epidermal cells and stomatal complexes. Scale bar: A,B,E–G = 1 cm; C = 0.5 cm; D,H = 0.2 cm; I,J = 50 μm.
Cuticles obtained from fossil needles preserve the following characters: Epidermal cells are rectangular and longitudinally oriented (Fig. 4I). The anticlinal walls of epidermal cells are straight or sinuous and slightly thickened. Stomatal complexes are paracytic, elliptical to oval in shape (Fig. 4J) and arranged in longitudinal rows. Well-developed cuticular flanges are preserved between guard cells and subsidiary cells. The polar subsidiary cells are smaller than the lateral subsidiary cells.
Discussion
Classification of the species with terminally-positioned umbones
Over 40 taxonomic treatments have been proposed for the genus Pinus4,23,24. The early classificatory systems are based mainly on morphology25,26, whereas more recent studies use molecular phylogenetic approaches4,6,23,27. The recent system of Pinus by Gernandt et al.6 based on rbcL + matK gene sequences and morphological characters is relatively widely accepted. This classification includes two subgenera (Pinus and Strobus), four sections (Pinus, Trifoliae Duhamel, Quinquefoliae, Parrya Mayr) and 12 subsections. The strictly North American section Parrya is restricted to the subsections Cembroides Engelmann, Nelsoniae Van Der Burgh and Balfourianae Engelmann and is characterized by a dorsal umbo on the ovulate cone scale. The subsections Strobus, Krempfianae Little et Critchfield and Gerardianae Loudon have been placed in the Eurasian and North American section Quinquefoliae. The group with a terminally-positioned umbo on the ovulate cone scale was regarded as one subsection (subsection Strobus) of section Quinquefoliae instead of subdividing them into subsection Strobus (the correct name for subsection Strobi6) and Cembrae Loudon4,23,26,27. In our study, the classification of Gernandt et al.6 is adopted.
The fossil cone
The fossil cone (Fig. 2 and Fig. 5) described above possesses the following morphological characters: (i), thin ovuliferous scales helically arranged around the axis; (ii), the apex of the ovuliferous scale is slightly inflated with an apophysis and terminal umbo. Consequently, this fossil cone can be assigned to the genus Pinus, according to distinguishing features summarized by Miller9. The umbo position and absence of a sealing band on the lower side of apophysis exclude a close affinity to the subgenus Pinus28 and judging from its distinct terminal umbo, this cone can be easily classified into the subgenus Strobus. The isolated ovuliferous fossil scale is considered to be conspecific with the cone due to the morphological similarities of the apophysis and umbo. Wing development and the size of seeds also are important in Pinus classification5. However, seeds discovered in the middle and upper part of the present cone provide only limited evidence for classification, principally because it is difficult to distinguish whether these seeds are winged or not.

Reconstruction of Pinus maomingensis sp. nov. (A–C) Three kinds of epidermal cells of Pinus maomingensis sp. nov.: (A) shows irregularly shaped cells, (B) shows the longitudinally elongate cells and (C) shows irregularly polygonal cells. Sketched from Fig. 3C,F. Arrows in the right show their positions in the ovuliferous scale. (D) Lower side of an ovuliferous scale. Its entire length was measured from the middle ovuliferous scales of Fig. 2A and width and shape of the apophysis was derived from the imprints of Fig. 2B,C,E. Details of umbo area derived from Fig. 2B,D,E. Scale bar = 1 cm. (E) Upper side of an ovuliferous scale sketched from a side view. (F) Seed which may have a wing. (G) Part of a longitudinal section of the cone. (H) General reconstruction of the cone with ovuliferous scales helically arranged around the axis. (A–H) drawn by Q.X. and S.N. Scale bar = 1 cm.
Comparisons with fossil cone taxa
Pinus existed at the onset of the Cretaceous and is well documented for the Cretaceous and Cenozoic29. Relatively few Cretaceous fossils have affinity with the subgenus Strobus and some specimens originally attributed to this subgenus were reassigned to the subgenus Pinus or other genera. Pinus magothensis Penny, from the Magothy Fm. of Delaware, U.S.A. is 9–10 cm long by 3–4 cm wide with thin flattened scales, each subtending two winged seeds; and with a thin apophysis and an inconspicuous terminal umbo13. There is no detailed description for the apophysis and umbo of P. magothensis, therefore it is difficult to compare it with P. maomingensis. Pinus magothensis was considered to be the most important evidence for the subgenus Strobus during the Cretaceous. However, Miller29 thought the characters were insufficient to show conclusive affinity with this subgenus due to the lack of details involving internal structure. As a result, the cone was reassigned to Pityostrobus by Miller and Malinky14. Willyard et al.15 also accepted this reassignment.
The fossil record of pines continues into the Paleogene and Neogene where fossil seed cones are more abundant and are overwhelmingly assigned to the subgenus Pinus10,14. The diversity of cone types, as well as the megafossil record of external impressions, implies that a number of species in various subsections were already in existence during the Eocene10. Those cones, such as Pinus lindgrenii Knowlton30, P. balfouroides Axelrod31 and P. sanjuanensis Axelrod10, appear assignable to the subsections of section Parrya and are excluded for comparison here. We focus on those fossil cones that exhibit affinity to the section Quinquefoliae, subsection Strobus sensu Gernandt et al.6.
The middle Eocene cone Pinus delmarensis Axelrod10, from the Del Mar Fm. near San Diego, California (U.S.A.) is larger (estimated as 26 cm long) than P. maomingensis, but is the most similar fossil species to our specimens. The similarities involve overall shape and the size of the ovuliferous scale and terminally positioned umbo. However, the lack of detailed published information on the apophysis and umbo of P. delmarensis precludes an informative comparison. Axelrod10 observed that P. delmarensis was very similar to the extant species P. lambertiana Douglas. However, the extant species P. lambertiana is readily distinguished from P. maomingensis (see the comparison with extant species below), indicating that additional detailed comparisons with P. delmarensis are unnecessary.
Pinus florissanti Lesquereux32 was established based on an ovoid seed cone with large scales from the upper Eocene Florissant Fm., Colorado, subsequently supplemented with seeds and needles by MacGinitie33. MacGinitie33 also reassigned P. sturgisi Cockerell34 to this species. Pinus florissanti was considered to be related to extant P. ponderosa Douglas ex C. Lawson (subgenus Pinus)32,33, but Axelord10 suggested that this fossil species was more closely allied to P. flexilis E. James (subgenus Strobus, section Quinquefoliae, subsection Strobus) because the characters of the cone and needles of P. florissanti were similar to those of the extant P. flexilis. Millar7 also supported the close affinity of P. florissanti with subsection Strobus. The cone of P. florissanti is shorter but wider (11 cm long by 6 cm wide33) than our specimens and the cone scales are shorter and narrower (4.5 cm long by 1.5 cm wide33). Furthermore, this fossil species differs from our specimens in having conical and rhomboidal umbo.
Pinus echinostrobus Saporta35 from the upper Oligocene of Armissan in Aude, France, has an ovate, flat and slightly striated apophysis with a terminal umbo. The nearest living relative of this species was considered to be P. koraiensis Siebold et Zuccarini36 which is distinguished from our specimen by having a reflexed apex with a basal-uncinate umbo. The cone of P. echinostrobus differs from our specimen in that the cone scales are shorter (1.0–1.4 cm long by 1.2–1.4 cm wide) and the apophysis is slightly curved and basally uncinate at the end of the umbo.
Pinus grossana Ludwig37 was recovered from the lower Miocene of Rockenberg locality of Wetterau, Germany. This cone is similar in width (5.5 cm) but is much longer (23 cm) than the present specimen according to the descriptions of Mai36. The thin cone scale with stripes on the abaxial side and the rhomboidal and slightly convex apophysis with a slightly reflexed apex are both similar to our specimens, although P. grossana is much wider and its length is unknown. In addition, this species differs from P. maomingensis in its erect, conical umbo. Ludwig37 thought that the nearest relative of this species was P. lambertiana. However, Mai36 considered that the overall features of the cone and the structure and testa anatomy of its associated winged seed indicated a relationship with extant P. wallichiana Jackson. This extant species is different from P. maomingensis in having wedge-shaped cone scales and grooved apophyses with an obviously incurved apex and a blunt umbo.
Pinus letzii Kirchheimer38 was described from the upper Miocene of the lower Rhenish Basin, Germany. The cones were 6–12 cm long by 2.5–3.5 cm wide and the cone scales were 2.5–3 cm long by 1–1.7 cm wide. The closest relative of P. letzii is thought to be P. dalatensis de Ferré36,39. The cone of P. letzii is shorter and narrower than that of P. maomingensis. Additionally, the flat apophyses of P. letzii are triangular or pentagonal in shape, with a triangular and terminal sunken umbo, whereas P. maomingensis bears rhombic to broadly rhombic apophyses with a diamond shaped and sunken umbo.
Pinus monticola var. fossilis40 from the Pliocene of Siberia, Russia, differs from our present specimens in possessing more robust and thicker cone scales. This species was once thought to be similar to the extant American species P. monticola Douglas ex D. Don41, but Axelrod10 assessed its affinities as more nearly allied to extant Asian species, notably P. armandii Franchet of central China. The extant species P. armandii differs from P. maomingensis in having a triangular or rhombic and thickened apophysis with an obtuse umbo. The Miocene cone P. itelmenorum Dorofeev42 from the Mammoth Mountain flora of Aldan River, Russia, is similar to P. monticola var. fossilis. However, this relatively complete fossil cone bears more massive and broader, thicker scales. Pinus itelmenorum also shows a relationship with the extant species P. armandii and its relatives and this fossil species can also be distinguished from our specimens.
Comparisons with extant cone taxa
Almost all members of subsection Strobus6 have been compared with Pinus maomingensis. The results show that all members of subsection Cembrae sensu Price et al.4 are easily distinguished from P. maomingensis because they bear an erect or basal-uncinate umbo. Other species, in the subsection Strobus sensu Price et al.4, including P. ayacahuite Ehrenberg ex Schlechtendal, P. flexilis James and P. lambertiana that bear an apical-uncinate or basal-uncinate umbo, are excluded. The other nine species which possess a similar apophysis shape and an obscure, terminal umbo are listed in Table 2 for detailed comparison. Pinus dalatensis and P. morrisonicola Hayata show less similarity with our specimens in having smaller cone scales with recurved or slightly recurved terminal umbones. Additionally, P. morrisonicola bears an obtuse apophysis apex. Pinus dabeshanensis Cheng and Law has a weakly developed umbo, but the apex of the apophysis is obtuse and the upper lateral side of the apophysis clearly is reflexed. The umbones of P. chiapensis (Martínez) Andresen, P. peuce Grisebrach and P. kwangtungensis Chen are flat, straight, or slightly incurved and this character is different from that of P. maomingensis. The species P. fenzeliana Handel-Mazzetti, P. parviflora Siebold et Zuccarini and P. wangii Hu et Cheng bear sunken to slightly sunken umbones which are very similar to the present specimens. However, the apex of apophysis of P. parviflora is rounded and its lateral side is reflexed. The lateral side of the apophysis of P. wangii is slightly incurved.
After detailed comparisons with the extant species, the results show that, although the cones of Pinus fenzeliana are somewhat smaller than the cone of P. maomingensis, it is the most similar extant species given its cone’s general shape, possession of both a broadly rhombic apophysis with slightly reflexed upper lateral side and a sunken umbo. Nevertheless, there is a subtle difference in the umbonal area. The middle part of the umbonal area of P. maomingensis is sunken and slightly folded and the transversely stretched ridge is situated near the center of the umbonal area and extends to the slightly reflexed apex. This character seems more similar to the cone scale apex of P. sibirica Du Tour of the subsection Cembrae sensu Price et al.4, but a key difference is that P. sibirica has a basal-uncinate umbo. Although we find two slight protrusions on the center umbonal area of our specimens, there is insufficient evidence to conclude that the fossil cone is a member of the subsection Cembrae sensu Price et al.4. Pinus maomingensis illustrates the close relationship between the subsections Strobus and Cembrae of Price et al.4 which were combined into one subsection (subsection Strobus) by Gernandt et al.6.
The cone morphology and epidermal structure of the cone scales of Pinus maomingensis was compared with modern species and was found to have similarities both to P. armandii and P. fenzeliana. Pinus armandii is similar to our specimens in the general size and shape of its cone and cone scales. However, epidermal structures of the fossil cone are more similar to those of P. fenzeliana. Both of these species possess three kinds of epidermal cells: longitudinally elongate cells, irregular polygonal cells and irregularly shaped cells. They also share longitudinally elongate cells and irregularly polygonal cells over the ribs and furrows. However, P. fenzeliana has longer cells over the ribs. The trichome bases of P. fenzeliana have one ring of epidermal cells and the shapes of these cells are different from those of the fossil cone. Pinus armandii has shorter epidermal cells over the ribs and the trichomes have two rings of epidermal cells surrounding their bases. However, they are distinguished from P. maomingensis by smaller numbers and the trapezoidal shape of epidermal cells.
Therefore, based on the detailed comparisons, we conclude that our specimens represent a new species of subgenus Strobus, section Quinquefoliae, subsection Strobus. We formally describe it as Pinus maomingensis sp. nov., in light of the following defining characters: (i) the shape and size of the cone; (ii) the shape and size of the cone scales; (iii) the shape, size and umbo of apophyses; (iv) epidermal structures.
The fossil needles
Abundant fossil needles of pinaceous affinity were collected from the Huangniuling Fm. of the Maoming Basin. A majority of the specimens have a bulbous base and deciduous sheaths. These characters also are diagnostic of needle fascicles of the subgenus Strobus3,4,5. Because scales of the fascicle sheaths abscise as the needles elongate in most species of this subgenus4, we speculate that our specimens were preserved in different growth stages, as we found a pair of sheath scales arising from surrounding bud scales that had not yet abscised. The bracts that subtend the fascicles are non-decurrent in subgenus Strobus4,5,28; this feature is clearly displayed in several fossil specimens. Moreover, characters of well-preserved fossil fascicles are very similar to subgenus Strobus needle fascicles described by Stults et al.43 from a Pliocene deposit in the coastal plain of the Gulf of Mexico. Both of these taxa have short, bulbous fascicle bases. Both species have deciduous sheaths. Consequently, we believe that the fossil needle fascicles associated with the same sediments as P. maomingensis also belong to subgenus Strobus.
The number of needles per fascicle is almost constant within many species of pines and frequently has been used as a species-specific character in many taxa2,4. Among the Haploxylon pines, the number of needles per fascicle always is five in section Quinquefoliae, but ranges from one to five in section Parrya. Fossil needle fascicles collected from the Huangniuling Fm. of the Maoming Basin mostly consist of five needles, although several specimens are preserved as three or four needles, possibly due to the preservation conditions of fossils or needles that fell singularly from mature fascicles with deciduous sheaths44. The margins of fossil needles are finely serrate. The needles of the extant species in the same section Quinquefoliae are finely serrate in eastern Asia and entire in North America10. Species of section Parrya that grow in China also possess finely serrate needles.
The anatomical characters of needles, such as the number of vascular bundles, the number and position of resin canals and cuticular structure also provide important characters for Pinus classification45,46,47,48. Although a large number of fossil needles were collected from the Huangniuling Fm. in the Maoming Basin, most are preserved as impressions and only a few specimens are preserved replete with cuticle fragments which show features of epidermal cells and stomatal complexes. Since cuticle characters of fossil needles were incomplete and lack anatomical structure in cross section, it is unreasonable to refer them to a certain species. These fossils are recognized as Pinus sp.
Biogeographic implications
The fossil record indicates that genus Pinus split into two subgenera by the Late Cretaceous. The earliest definitive representative of the subgenus Strobus was discovered in the Late Cretaceous (Santonian) Aachen Fm. of northeast Belgium16. During the Paleogene, the subgenus Strobus was common in Eurasia and North America10,36,49 but very rare in China. Pinus maomingensis sp. nov. and Pinus sp., collected from the Huangniuling Fm. in the Maoming Basin of Guangdong Province, assuredly belongs to subgenus Strobus. The discovery of these fossils indicates that white pines were distributed in southern China by at least the late Eocene.
Palynological assemblages from the Huangniuling Fm. of the Maoming Basin of Guangdong suggest that the late Eocene was warm and humid22. In addition, many fossil plants collected in the Maoming Basin, such as Podocarpaceae, Arecaceae, Dipterocarpaceae, Annonaceae, Juglandaceae, Euphorbiaceae, Myrtaceae, Fagaceae, Altingiaceae and Lauraceae22, provide evidence for tropical-subtropical components in the Eocene Maoming flora. The subgenus Strobus mainly inhabits the north and tropical-subtropical mountainous regions of China and grows well in habitats characterized by temperate and moist climate5,50,51,52. Presently, however, white pines do not occur in or adjacent to the Maoming Basin. Currently, only two species—Pinus fenzeliana and P. kwangtungensis—are naturally distributed in mid to high altitude areas of southern China. These two species grow at an altitude of ca. 1000 m in the northern part of Guangdong Province (Lechang and Ruyuan Mountains) and the Wuzhi Mountains of Hainan Island5. Because of these distributional patterns, we propose that the Maoming Basin was adjacent to a mountainous region during the late Eocene.
Methods
A seed cone, one isolated ovuliferous scale and large numbers of needles were collected from the Huangniuling Fm. of the Maoming Basin, southern China. This cone and the isolated ovuliferous scale are preserved as compressions. Cuticular fragments were obtained from the distal part of the ovuliferous scales of the compressed cone (MMJ2-1-005b). Most needles are impressions and only a few needle fragments preserved, albeit poorly, epidermal remains. Leaf cuticular fragments were obtained from specimen MMJ3-002a. Specimens were photographed using a Canon EOS 500D digital camera. Photomicrography of the seeds was done with using a Leica S8ap0 and Image-Pro software. The terminology for morphological description of subgenus Strobus follows Klaus53, Fu et al.5 and Earle3.
Fossil cuticle pieces were removed from the specimens and immersed in 30% HNO3 solution overnight and then washed in distilled water 2 or 3 times. The samples then were treated with very weak (ca. 1–2%) ammonia for 4 hours and then washed with distilled water. Cuticles were mounted on glass slides and observed and photographed using a Nikon microscope in transmitted light and processed with Adobe Photoshop CS5 (Adobe Inc., San Jose, California, USA). All the fossil specimens and slides are stored at the Museum of Biology of Sun Yat-sen University, in Guangzhou, China.
Additional Information
How to cite this article: Xu, Q. et al. Late Eocene white pines (Pinus subgenus Strobus) from southern China. Sci. Rep. 5, 16390; doi: 10.1038/srep16390 (2015).
References
Li, N. Studies on the geographic distribution, origin and dispersal of the family Pinaceae Lindl. Acta Phytotax. Sin. 33, 105–130 (1995).
Richardson, D. M. & Rundel, P. W. Ecology and biogeography of Pinus: an introduction. in Ecology and Biogeography of Pinus (ed Richardson, D. M. ) 3–46 (Cambridge University Press, Cambridge, 1998).
Earle, C. J. The Gymnosperm Database. (2014). Available at: http://www.conifers.org/. (Accessed: 5th December 2014).
Price, R. A., Liston, A. & Strauss, S. H. Phylogeny and systematics of Pinus. in Ecology and Biogeography of Pinus (ed Richardson, D. M. ) 49–68 (Cambridge University Press, Cambridge, 1998).
Fu, L. G., Li, N. & Mill, R. R. Pinaceae. inFlora of China, Vol. 4 (eds Wu, Z. Y. & Raven, P. H. ) 11–52 (Science Press, Beijing & Missouri Botanical Garden Press, St. Louis, 1999).
Gernandt, D. S., Lopez, D. S., Geada, G. G., Garcia, S. O. & Liston, A. Phylogeny and classification of Pinus. Taxon 54, 29–42 (2005).
Millar, C. I. Impact of the Eocene on the evolution of Pinus L. Ann. Mo. Bot. Gard. 80, 471–498 (1993).
Ryberg, P. E., Rothwell, G. W., Stockey, R. A., Hilton, J. & Mapes, G. Reconsidering relationships among stem and crown group Pinaceae: oldest record of the genus Pinus from the early Cretaceous of Yorkshire, United Kingdom. Int. J. Plant Sci. 173, 917–932 (2012).
Miller, C. N. Early evolution in the Pinaceae. Rev. Palaeobot. Palyno. 21, 101–117 (1976).
Axelrod, D. I. Cenozoic history of some western American pines. Ann. Mo. Bot. Gard. 73, 565–641 (1986).
Jeffrey, E. C. On the structure of the leaf in Cretaceous pines. Ann. Bot. 22, 207–220 (1908).
Stopes, M. C. & Kershaw, E. M. The anatomy of Cretaceous pine leaves. Ann. Bot. 24, 395–402 (1910).
Penny, J. Studies on the conifers of the Magothy flora. Am. J. Bot. 34, 281–296 (1947).
Miller, C. & Malinky, J. Seed cones of Pinus from the late Cretaceous of New Jersey, USA. Rev. Palaeobot. Palyno. 46, 257–272 (1986).
Willyard, A., Syring, J. & Gernandt. D. S. Fossil calibration of molecular divergence infers a moderate mutation rate and recent radiations for Pinus. Mol. Biol. Evol. 24, 90–101 (2007).
Meijer, J. Fossil woods from the late Cretaceous Aachen Formation. Rev. Palaeobot. Palyno. 112, 297–336 (2000).
Yi, T. M., Li, C. S. & Jiang, X. M. Conifer woods of the Pliocene age from Yunnan, China. J. Integ. Plant Biol. 47, 264–270 (2005).
Yi, T. M., Li, C. S., Jiang, X. M. & Wang, W. F. Pliocene Rhododendron and Pinus fossil woods of Yunnan Province and palaeoclimate. J. Paleogeogr. 4, 90–98 (2002).
Bureau of Geology Mineral Resources of Guangdong Province. Regional geology of Guangdong Province. (Geological Publishing House, Beijing, 1988).
Nan, Y. & Zhou, G. Q. Stratigraphy (lithostratic) of Guangdong Province. (Wuhan, China University of Geosciences Press, 1996).
Wang, J. D. et al. Magnetostratigraphy of Tertiary Rocks from Maoming Basin, Guangdong Province, China. Chinese J. Geochem. 13, 165–175 (1994).
Aleksandrova, G. N., Kodrul, T. M., Liu, X. Y., Song, Y. S. & Jin, J. H. Palynological characteristics of the upper part of the Youganwo Formation and lower part of the Huangniuling Formation, Maoming Basin, South China. in Proceedings of the 2nd Sino-Russian Seminar on Evolution and Development of Eastern Asia Flora based on Palaeobotanical Data (ed Tang B.). 3–15 (School of Life Sciences, Sun Yat-sen University, Guangzhou 2012).
Eckert, A. & Hall, B. Phylogeny, historical biogeography and patterns of diversification for Pinus (Pinaceae): phylogenetic tests of fossil-based hypotheses. Mol. Phylogenet. Evol. 40, 166–182 (2006).
Wang, B. S. Hybridization and evolution in the genus Pinus. (Umeå University, Umeå, Sweden, 2013).
Shaw, G. R. The genus Pinus, Publications of the Arnold Arboretum No. 5. (Riverside Press, Cambridge, 1914).
Little, E. L. & Critchfield, W. B. Subdivisions of the genus Pinus (pines). (US Forest Service Miscellaneous Publication, Washington DC, 1969).
Liston, A., Robinson, W. A., Piñero, D. & Alvarez-Buylla, E. R. Phylogenetics of Pinus (Pinaceae) based on nuclear ribosomal DNA internal transcribed spacer region sequences. Mol. Phylogenet. Evol. 11, 95–109 (1999).
Frankis, M. (2002). Classification of the genus Pinus. Available at: http://www.pinetum.org/Lovett/classification.htm (Accessed: 28th February 2014).
Miller, C. N. Mesozoic conifers. Bot. Rev. 43, 217–280 (1977).
Knowlton, F. H. A fossil nut pine from Idaho. Torreya 1, 113–115 (1901).
Axelrod, D. I. History of the maritime closed-cone pines, Alta and Baja California. Vol. 120, 1–143 (University of California Press, California, 1980).
Lesquereux, L. Contribution to the fossil flora of the western territories, Part III. The Cretaceous and Tertiary floras. Rept. U. S. Geol. Surv. Terr. 8, 1–283 (1883).
MacGinitie, H. D. Fossil plants of the Florrissant beds, Colorado. Carnegie (Institution of Washington Publication, Washington DC, 1953).
Cockerell, T. D. A. Descriptions of Tertiary plants, II. Am. J. Sci. 26, 537–544 (1908).
Saporta, G. de . Etude sur la végétation du Sud-Est de la France à l'époque tertiaire, Armissan près de Narbonne (Aude). Ann. Sci. Nat., Bot. 5, 5–264 (1865).
Mai, D. H. Über Typen und Originale tertiärer Arten von Pinus L. (Pinaceae) in mitteleuropäischen Sammlungen-Ein Beitrag zur Geschichte der Gattung in Europa. Feddes Repert. 97, 571–605 (1986).
Ludwig, R. Fossile Pflanzen aus der ältesten Abteilung der Rheinisch-Wetterauer Tertiär-Formation. Palaeontographica 8, 39–154 (1859).
Kirchheimer, F. Über die Pflanzenreste in den Begleitschichten der Braunkohle von Düren. Palaeont. Z. 18, 213–227 (1936).
Jährig, M. Beiträge zur Kenntnis der Sippenstruktur der Gattung Pinus L. Arch. Forstwes. 17, 173–205 (1968).
Sukachev, V. N. Nekotorye dannye k dolednikovoi flore severa Sibiri. Akad. Nauk, Geologischeskii musei imeni Petra Velikogo Trudy 5, 55–62 (1910).
Wolfe, J. A. & Leopold, E. B. Neogene and early Quaterary vegetation of Northwestern North America and northeastern Asia. in The Bering Land Bridge (ed Hopkins, D. M. ). 193–206 (Stanford University Press, California, 1967).
Dorofeev, P. I. The Miocene flora of the Mammoth Mountain on the Aldan River. (Akademia Nauk CCCP, 1969).
Stults, D. Z., Axsmith, B. J. & Liu, Y. S. (C) . Evidence of white pine (Pinus subgenus Strobus) dominance from the Pliocene Northeastern Gulf of Mexico Coastal Plain. Palaeogeogr. Palaeoclimatol. Palaeoecol. 287, 95–100. (2010).
Farjon, A. A Handbook of the World’s Conifers. (Brill Academic Publishers, Leiden, Netherlands, 2010).
Yoshie, F. & Sakai, A. Types of Florin rings, distributional patterns of epicuticular wax and their relationships in the genus Pinus. Can. J. Bot. 63, 2150–2158 (1985).
Hu, Y. S. SEM observations of the inner surface structure of needle cuticles in Pinus. Acta Phytotax. Sin. 24, 464–468 (1986).
Stružková, D. Cuticular analysis – A method to distinguish the leaves of Pinus sylvestris L. (Scots Pine) from those of Pinus mugo Turra s. str. (Dwarf Mountain-pine). Veg. Hist. Archaeobot. 11, 241–246 (2002).
Whang, S. S., Kim, K. & Hill, R. S. Cuticle micromorphology of leaves of Pinus (Pinaceae) from North America. Bot. J. Linn. Soc. 144, 303–320 (2004).
Klaus, W. Mediterranean pines and their history. Plant Syst. Evol. 162, 133–163 (1989).
Wu, G. & Feng, Z. W. The sociological characteristics and biomass of stone pine forests in China. Acta Ecol. Sin. 15, 260–267 (1995).
Liu, G. F. et al. Analysis of genetic relationship in 12 species of Section Strobus with ISSR markers. J. Forest. Res. 16, 213–215 (2005).
Sun, A. Z., Ma, Y. Z., Huang, C. Q. & Wu, H. N. Study of the pollen morphologic appraisable point of pinaceous main genera and their ecological environment. J. Lanzhou Univ. (Natural Sci.) 42, 18–21 (2006).
Klaus, W. Neue Beobachtungen zur Morphologie des Zapfens von Pinus und ihre Bedeutung für die Systematik, Fossilbestimmung, Arealgestaltung und Evolution der Gattung. Plant Syst. Evol. 134, 137–171 (1980).
Acknowledgements
This study was supported by the National Natural Science Foundation of China (Grant Nos. 41210001, 41572011), the National Basic Research Program of China (973 Program) (Grant No. 2012CB822003), the joint Project of the National Natural Science Foundation of China and the Russian Foundation for Basic Research (Grant Nos. 413111040, 14-05-91163), State Key Laboratory of Palaeobiology and Stratigraphy (Nanjing Institute of Geology and Palaeontology, CAS) (Grant No. 123110), the Fundamental Research Funds for the Central Universities (Grant No.12lgjc04), the Key Project of Sun Yat-sen University for inviting foreign teachers and the Scientific Research Fund, Hongda Zhang, Sun Yat-sen University, the State Project of the Geological Institute, Russian Academy of Sciences (Grant No. 01201459195) and the State Scholarship Fund of China Scholarship Council (CSC) (No. 201406380049). The authors thank Botany graduate students at Sun Yat-sen University for their field work and fossil collecting. We greatly appreciate the National Museum of Natural History, Smithsonian Institution, especially for letting us use the library resources. We also offer our sincere gratitude to Ms. Margaret Joyner (U.S.A.) for editing and Prof. Conrad Labandeira and Mr. Thomas F. Jorstad (U.S.A.) for modification.
Author information
Authors and Affiliations
Contributions
J.J., W.Z. and Q.X. did the fieldwork. J.J., Q.X. and S.N. took photos for the specimens. Q.X., J.J., T.K. and W.Z. discussed the results and wrote the main manuscript text. Q.X. prepared the figures and S.N. and Q.X. made illustrative drawing (Fig. 5). All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Xu, Q., Zhou, W., Kodrul, T. et al. Late Eocene white pines (Pinus subgenus Strobus) from southern China. Sci Rep 5, 16390 (2015). https://doi.org/10.1038/srep16390
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep16390
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
