
Abstract
The green alga Scenedesmus is known for its phenotypic plasticity in response to grazing risk. However, the benefits of colony formation induced by infochemicals from zooplankton should come with costs. That is, a tradeoff in benefit-to-cost ratios is likely under complex environmental conditions. In this study, we hypothesized that the coexistence of Scenedesmus and its competitors decreases the formation of anti-grazer colonies in Scenedesmus. Results demonstrated that the presence of a competitor Microcystis aeruginosa inhibited inducible defensive colony formation of Scenedesmus obliquus and the established defensive colonies negatively affected the competitive ability of S. obliquus. The proportion of induced defensive colonies in cultures was dependent on the relative abundance of competitors. Under low competition intensity, large amount of eight-celled colonies were formed but at the cost of decreased competitive inhibition on M. aeruginosa. By contrast, defensive colony formation of S. obliquus slacked in the presence of high competition intensity to maintain a high displacement rate (competitive ability). In conclusion, S. obliquus exhibited different responses to potential grazing pressure under different intensities of competition, i.e., Scenedesmus morphological response to grazing infochemicals was affected by competition against Microcystis.
Similar content being viewed by others


Introduction
The production of different phenotypes from a single genotype in response to environmental conditions is termed “phenotypic plasticity”, which is common in organisms1,2,3. Phenotypic plasticity is an adaptation to natural environmental selection pressures4. The ability to adapt to multiple environments is very important for the evolutionary success of organisms4,5. Among the biotic and abiotic factors that alter the expression of organisms’ genotypes, predation pressure plays an important role. Organisms may respond to the selective pressure of predation by producing defensive phenotypes against predators. For example, to avoid being attacked by lake crabs, blue mussels Mytilus edulis are induced by predation cues to increase shell thickness, enhance abductor muscle strength and produce more byssal threads6. Similarly, Daphnia pulex produces neck teeth on the dorsal anterior margin of the head in the presence of the predator Chaoborus americanus7. Predation-induced phenotypic plasticity is also prevalent in plants. Some plants produce toxins and defensive proteins or emit volatiles to discourage herbivores8.
Competition and predation are considered the major selective pressures that shape the composition of pelagic photoautotrophic communities9. In general, edible phytoplankton have less competitive advantage than resistant phytoplankton under high grazing pressure10. Many phytoplankton species have developed morphological and chemical defenses or other mechanisms to resist the attack of predators and avoid loss of biomass caused by grazing11,12. As for unicellular algae, which are smaller than their predatory enemies, one effective way to resist grazing threat is to increase in size13. Induced colony formation in the green alga Scenedesmus spp. is a typical case. Upon detection of infochemicals released by herbivores such as Daphnia, Scenedesmus populations become dominated by four- and eight-celled colonies instead of unicells14. Scenedesmus populations that are dominated by colonies can effectively deter feeding by small grazers such as Daphnia cucullata; therefore, induced colony formation is an efficient strategy for Scenedesmus to escape grazing by predators with a diet limited by the prey size15.
However, large Scenedesmus colonies experience a higher risk of sedimentation than unicells16. The light dependence of pelagic photoautotrophic organisms limits their size17. Large algal particles without additional buoyancy mechanisms, such as gas vesicles, may have limited capacity to suspend in the euphotic zone. Increase in colony size also decreases their surface-to-volume ratios. Algae absorb nutrients through the cellular membrane and large surface-to-volume ratios benefit nutrition uptake and light capture17,18. Under turbulent diffusion, phytoplankton species are mixed and coexist in water. All phytoplankton species and their competitors require almost the same nutrients from the same resource pool19. As a result, competition for light and nutrients will be an essential requirement for the population of phytoplankton to sustain domination in phytoplankton communities20,21. Hence, trade-offs potentially exist between the formation of large sinkable colonies (favoring defense) and maintenance of vulnerable unicells (favoring competition) in Scenedesmus.
The disadvantages of forming colonies, including accelerated sinking rate and decreased nutrition and light absorption, may affect the competitive ability of Scenedesmus. It has been recognized that organisms face trade-offs to allocate the limited resources to grow or to counteract stress22,23. The trade-off between competitive ability and resistance to predators or parasitoids is common in many species. For example, defended algae have less competitive ability than undefended species24,25.
Considering the trade-off mechanism and the high morphological plasticity of Scenedesmus in response to infochemicals released by grazers, we hypothesized that defense and competition operate at the expense of each other and Scenedesmus may make trade-offs under pressure from both predators and competitors. Accordingly, this study aimed to examine the costs and the trade-off of forming grazer-induced defensive colonies in Scenedesmus under different levels of inter-species competition. Microcystis aeruginosa, one of the most common bloom-forming cyanobacteria, was used as the competitor of S. obliquus. Both algae are found in freshwaters world-wide and often coexist in the same area26,27. M. aeruginosa has been reported to have significant allelopathic influence on co-existing phytoplankton28,29,30,31 and to produce secondary metabolites that are toxic to zooplankton32. In natural waters or under the grazing induction of predators33, M. aeruginosa can also form huge thousand-celled colonies, but the M. aeruginosa strain selected in this study was unicellular with no colony formation when exposed to Daphnia grazing infochemicals34. We used filtered test water containing infochemicals from Daphnia magna to simulate S. obliquus colony formation in the presence of competitors.
Results
Monocultures of S. obliquus in the control groups were dominated by unicells over the 7 d period, with less than 12% large eight-celled coenobia (Fig. 1a). The morphology of S. obliquus monocultures changed drastically under exposure to Daphnia filtrate compared with the control. The proportion of unicells decreased rapidly from day 2 and four- and eight-celled coenobia formed (Fig. 1b). Colony formation reached its maximum on day 4 (Fig. 2a) and the proportion of four- and eight-celled coenobia increased to approximately 85%. On day 5, the proportion of eight-celled coenobia decreased from more than 60% to approximately 40%. The mean number of cells per particle in the control groups remained below two during the 7 d period. In this experiment, the S. obliquus population of each culture was composed by unicells and two-, four- and eight-celled colonies; a few three-, five-, six- and seven-celled colonies were also present, but coenobia larger than eight cells were not observed.

Proportions of unicells and 2-, 4- and 8-celled ceonobia in Scenedesmus populations of monocultures in the absence
(a) and presence (b) of Daphnia filtrate, in co-cultures with different initial compositions (Scenedesmus:Microcystis = 1:1, absence (c) or presence (d) of Daphnia filtrate; and Scenedesmus: Microcystis = 1:5 absence (e) or presence (f) of Daphnia filtrate during the 7-day experiment. The “rest” group represents 3-, 5-, 6-, 7-celled colonies.

Changes in the mean number of cells per particle of Scenedesmus in pure cultures of Scenedesmus
(panel a) and in co-cultures of Scenedesmus and Microcystis with different initial compositions (panel b: Scenedesmus: Microcystis = 1:1; panel c: Scenedesmus : Microcystis = 1:5) in the absence (black bars) and presence (white bars) of Daphnia filtrate. Vertical lines represent 1SE.
In co-cultures of the control groups, the S. obliquus populations were dominated by unicells as the monocultures (Fig. 1c,e). However, the defensive morphological features of S. obliquus induced by Daphnia filtrate were significantly altered when M. aeruginosa was added as a competitor to the S. obliquus cultures. The induced colony formation of S. obliquus decreased in the presence of M. aeruginosa (Fig. 2b,c). The mean number of cells per particle of S. obliquus in the Daphnia filtrate-treated groups co-cultured with 1-fold M. aeruginosa reached its maximum on day 4 (Fig. 2b), which was similar to the pattern of the Daphnia filtrate-treated monocultures (Fig. 2a). By contrast, the colony size decreased rapidly from day 5 and unicells became dominant again. The proportion of unicells was approximately 40% on day 7 (Fig. 1d), which was significantly higher than that in the monocultures (Fig. 1b). However, in the co-cultures with 5-fold M. aeruginosa, the average colony size of S. obliquus was smaller than two cells even though colonies of S. obliquus were significantly induced by Daphnia filtrate (compared with the control group in this experiment, F = 67.398, p < 0.001) (Fig. 2c). Unicells accounted for approximately 40–48% of the total populations from day 2 to the end of the experiment (Fig. 1f). The competitive effect of M. aeruginosa emerged on day 3. Formation of four- and eight-celled large coenobia of S. obliquus ceased (Fig. 1d,f). In the control groups, M. aeruginosa was unicellular or paired cellular. M. aeruginosa exposed to Daphnia filtrate failed to form colonies and were found predominantly as unicells and paired cells.
Growth of S. obliquus decreased significantly in the Daphnia filtrate-treated co-cultures with 1-fold M. aeruginosa when compared to the control monoculture group (Fig. 3; one-way ANOVA; p = 0.001). Growth rates of S. obliquus in other four group cultures showed no significant differences compared with the control monoculture group. Two-way ANOVA was conducted to analyze the influence of Daphnia filtrate and the density of competitors on the growth rates of S. obliquus and M. aeruginosa (Table 1). Results showed that Daphnia filtrate had a strong effect on the growth rates of S. obliquus and significant interaction was detected between these two factors. M. aeruginosa had significantly higher growth rates in co-cultures with higher initial cell density. Daphnia filtrate had no significant effect on M. aeruginosa growth.

Growth rates of S. obliquus and M. aeruginosa populations incubated under different conditions during the 7-day experiment.
Vertical lines represent 1 SE, short lines with asterisk above two bars represent significant difference (p < 0.05) within groups.
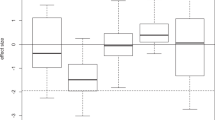
Separate linear regressions were fitted for all co-cultures to compare the displacement of M. aeruginosa by S. obliquus between the control and Daphnia filtrate-treated groups (Fig. 4). The slopes of Y(t) versus t were positive in all cultures, indicating that S. obliquus could displace M. aeruginosa in both experiments. In co-cultures with the same initial cell densities of both algae, the slope decreased in the Daphnia filtrate-treated group compared with the control, but the decrease was not statistically significant (F = 1.723, p = 0.196). The displacement rates during days 3 to 7 were further analyzed, because colony formation induced by Daphnia infochemicals peaked on day 3. Since multicellular colonies have lower surface area-to-volume ratios than unicells, nutrition uptake of S. obliquus may be affected under competition by M. aeruginosa and the potential costs may emerge after day 3. Significant decrease was detected in the Daphnia filtrate-treated group (F = 4.789, p = 0.037). For co-cultures with 5-fold M. aeruginosa, the slopes of the control and Daphnia filtrate-treated groups were similar (F = 0.843, p = 0.212). Coexistence with M. aeruginosa decreased the formation of inducible colonies by S. obliquus for potential reallocation of resources in response to competition.

Comparison of displacement rates for Scenedesmus between controls and Daphnia filtrate-treated groups in the presence of Microcystis with different initial densities.
Linear regressions were separately fitted for different treatments.
Discussion
Scenedesmus is considered to benefit from the ability of forming plastic morphs in response to biotic and abiotic environmental factors, particularly herbivory15. The algal defense theory for freshwater systems predicts that edible species of algae with no or weak defensive mechanisms are more vulnerable under the selective stress of predation35,36. Resistant algae with effective defensive mechanisms can escape from being consumed and replace edible algae in algal communities12. However, some grazer-resistant species have slower growth rates than undefended ones25,37, so it has been suggested that resistance to herbivory could be costly. For Scenedesmus, the benefits of being plastic have been well documented15,38, but the direct costs and limits involved in inducible resistance still lack enough evidence. The higher sinking rates of coenobia compared with unicells might be the only indirect cost detected in multicellular Scenedesmus16. Yokota and Sterner24 showed that coenobial Desmodesums subspicatus preinduced and prolonged by Daphnia infochemicals had suppressed competitive ability compared to unicellular ones. Thus, it is possible that colonial morph interferes with the rapid growth of S. obliquus for the reduction of resource uptake owing to its decreased surface area-to-volume ratio. Therefore, we hypothesized that Scenedesmus formed different morphs as a trade-off between defense and competition.
In the present study, we observed a trade-off in S. obliquus between defense and growth in the presence of competitors. Competitive displacement rates indicated that colony formation exerted adverse effects on the growth and competitive ability of S. obliquus (Fig. 4). Formation of large defensive colonies is a type of adaptive phenotypic plasticity under high predation stress, but this morphology will be a liability to rapid growth of the S. obliquus population when the threat is relieved and thus competition stress is increasing (Fig. 4). The Scenedesmus population of large defensive coenobia is eventually redominated by unicells when infochemicals in the water are eliminated39 and enhanced competition accelarated the deformation of colonies. This strategy presumably benefits organisms by facilitating resource allocation between defense and growth40. In this study, S. obliquus showed highly morphological plasticity in the presence of competitors with different initial densities. Under different degrees of competitive stresses, the population of S. obliquus balanced the formation of defensive colonies and growth of the population. M. aeruginosa is a common bloom-forming phytoplankton that can coexist with Scenedesmus in temperate eutrophic lakes41. In the presence of relatively low competitive pressure from M. aeruginosa, S. obliquus responded to the grazer infochemicals by forming a high percentage of colonies at the expense of decreased competitive exclusion rate. The significant decline in the displacement rate from day 3 observed in this study may be the remarkable evidence of cost associated with colony formation. With proliferation of both algae, inter-specific competition for resources increased. The negative effect on the competitive ability of defensive colonies might inhibit further formation of colonies, as evidenced by the gradual decrease in colony size from day 4. When the initial density of M. aeruginosa was increased to 5-fold, competitive stress increased, limitations of phenotypic plasticity became prominent and weak morphological modulations were detected in S. obliquus. Defense to herbivory was reduced to maintain competitive superiority under high competitive stress.
As the two major selective pressures affecting phytoplankton, inter-specific competition and predation can shape the composition of algal communities42. Intra-specific variability provided by phenotypic plasticity is known to be the mechanism that enhances the competitive and adaptive abilities of phytoplankton under varying environmental conditions43,44. Defensive responses to predators may alter the competition between prey species45. In the present study, as we hypothesized, inter-specific competition was found to influence the plastic response of Scenedesmus to herbivory. Inducible defense is more energy-saving than constitutive defense in the long term, but both defensive mechanisms are expensive46. One of the main benefits of the “just-in-time” inducible defense system is effectiveness47. The investment in a plastic defense mechanism is more effective for organisms especially when facing multiple environmental pressures. Continuous reproduction of large colonies would affect the competitive ability of Scenedesmus24 and unicellular or small colonial forms would be favored under intense competitive environments. The failure to form typical anti-predator colonies might be due to the strong competition pressure and the sense of relative reduction in the risk of being grazed on. Because higher density of co-existing species will relieve the grazing pressure on a single species, there is no need for S. obliquus to form colonies that have higher sedimentation rate and lower resource acquisition at the expense of competitive ability.
Besides the possible trade-off of S. obliquus between forming defensive colonies and competition, there are other possible reasons for decreased colony formation in S. obliquus when competitors co-exist. M. aeruginosa may interfere with the regular response of S. obliquus to grazing cues, as M. aeruginosa can produce secondary metabolites that have inhibitory effects on photoautrophic algae29,31; these allelochemicals may interfere with the response of S. obliquus to Daphnia infochemicals. In addition, some infochemicals released by Daphnia may be absorbed by M. aeruginosa cells (despite M. aeruginosa not showing change in morphology), which lowers the amount of infochemicals detected by S. obliquus cells and thus lowers their morphological response.
In summary, the presence of a competitor M. aeruginosa inhibited inducible defensive colony formation of S. obliquus. The proportion of induced defensive colonies in cultures was dependent on the relative abundance of competitors. Under low competition intensity, large amount of eight-celled colonies were formed but at the cost of decreased competitive inhibition on M. aeruginosa. By contrast, defensive colony formation of S. obliquus slacked in the presence of high competition intensity to maintain a high competitive ability. S. obliquus exhibited different responses to potential grazing pressure under different intensities of competition. These results demonstrate that phenotypic plasticity allows organisms to undergo controllable phenotypic changes to reduce fitness costs by optimizing the benefit-to-cost ratios when responding to different environmental challenges.
Methods
Microorganisms and culture conditions
S. obliquus (Chlorophyta) FACHB-416 and M. aeruginosa (Cyanobacteria) FACHB-927 were obtained from the Freshwater Algae Culture Collection at the Institute of Hydrobiology China (FACHB-collection) and maintained in our laboratory in a modified BG-11 medium48 with ammonium NH4Cl as nitrogen source. Before the experiment, stock algae were regularly transferred to fresh media at a ratio of 0.3 d−1 for 10 d to maintain the logarithmic growth phase. The grazer Daphnia magna used to produce infochemicals in the experiment was a laboratory clone. The only food source of D. magna during the cultivation was S. obliquus grown in continuous cultures. The stock cultures of S. obliquus, M. aeruginosa and D. magna were all stored in a climate-controlled chamber at 25 °C under fluorescent light at 40 μmol quanta·m−2·s−1 in a 14:10 h light-dark cycle. All experiments were conducted under these conditions.
Preparation of control and Daphnia filtrates
Since the infochemicals that evoke anti-grazing colonies of Scenedesmus were demonstrated to be associated with the grazing progress and linked to alga-grazer interaction11,49, it is necessary to use “Daphnia feeding Scenedesmus” to obtaining the infochemicals. Test water containing infochemicals from D. magna was obtained as previously described15,50. In brief, D. magna was washed to remove surface contamination and allowed to evacuate their guts in sterile distilled water for at least 12 h and then incubated at a density of 300 ind·L−1 with sufficient S. obliquus as food. After 24 h, D. magna was removed and the cultures were filtered through a 0.10 μm membrane filter (Millipore Corporation, USA). The filtrates served as the treatment media containing infochemicals released by Daphnia. Cultures of Scenedesmus without Daphnia were filtered as well and used as the control media. NH4Cl was used as nitrogen source to easily adjusting the nitrogen levels of filtrates since the nitrogenous metabolites of the crustacean is ammonia. Appropriate ammonium and phosphate were added to the control filtrate according to the measured concentrations of nitrogen and phosphorus in the Daphnia filtrate and thus the nitrogen and phosphorus concentrations in the treatment and control media were adjusted to the same levels.
Induction experiment under competition with Microcystis
A flow diagram of experimental design was shown in Fig. 5. Considering the differences in cell size between S. obliquus and M. aeruginosa, we executed the experiment using two initial density proportions (Scenedesmus: Microcystis = 1:1 and 1:5) with 10% filtered S. obliquus culture medium or Daphnia filtrate. The initial algal density of S. obliquus in each culture was approximately 1.0 × 105 cells·mL−1 and the initial algal densities of Microcystis were approximately 1.0 × 105 and 5.0 × 105 cells·mL−1, respectively. In the experiments, algal suspensions were transferred to 250 mL cellulose-plug stoppered Erlenmeyer flasks containing 150 mL of medium. Six treatments were carried out and each treatment was run in triplicate. To avoid sedimentation, the algal cultures were manually shaken twice a day.

Flow diagram of the experimental design
.
Microscopic observation
Samples (1 mL) were collected daily and preserved in Lugol’s fixative (2%). Algal densities and different-celled colony distributions were counted under a microscope (Olympus 6V20WHAL; Tokyo, Japan) using a hemocytometer(Qiujing HT-042; Shanghai, China). Different algal particles (unicells, two-celled, four-celled, eight-celled and the rest) were counted and the mean proportions of cells in different morphs were calculated. The numbers of cells per particle were calculated by dividing the total cell number by the number of particles. Growth rates of S. obliquus and M. aeruginosa were determined as the slope of logarithm abundance versus time51. To reveal the potential cost of grazer-induced defenses in Scenedesmus under competitors, the competitive displacement rate was adapted52,53 as an index in this study. The natural log of the ratio of the abundances of the two algae, Y(t) = ln[NScenedesmus(t)/NMicrocystis(t)], was calculated and regressed against time (d). The displacement rates for Scenedesmus were the slopes of the linear regressions24.
Statistical analysis
Data values are expressed as means of 3 replicates ± SE. Colony formation, proportions of different celled populations and growth rates were compared by two-way ANOVA, with the presence/absence of the competitor and Daphnia filtrate as the two factors. Significant differences between treatments were detected by t-test. Statistical analyses were performed using SigmaPlot 11.0. The differences between slopes of the linear regressions of Y(t) versus t were analyzed by ANCOVA in SPSS 10.0.
Additional Information
How to cite this article: Zhu, X. et al. Grazer-induced morphological defense in Scenedesmus obliquus is affected by competition against Microcystis aeruginosa. Sci. Rep. 5, 12743; doi: 10.1038/srep12743 (2015).
References
Travis, J. in Ecological morphology: integrative organismal biology Vol. 101 (eds Wainwright, P. C. & Reilly, S. M. ) 99–122 (University of Chicago Press, 1994).
Bradshaw, A. D. Evolutionary significance of phenotypic plasticity in plants. Adv. Genet. 13, 115–155 (1965).
Fordyce, J. A. The evolutionary consequences of ecological interactions mediated through phenotypic plasticity. J. Exp. Biol. 209, 2377–2383 (2006).
Price, T. D., Qvarnström, A. & Irwin, D. E. The role of phenotypic plasticity in driving genetic evolution. Proc. R. Soc. B: Biol. Sci. 270, 1433–1440 (2003).
Miner, B. G., Sultan, S. E., Morgan, S. G., Padilla, D. K. & Relyea, R. A. Ecological consequences of phenotypic plasticity. Trends Ecol. Evol. 20, 685–692 (2005).
Leonard, G. H., Bertness, M. D. & Yund, P. O. Crab predation, waterborne cues and inducible defenses in the blue mussel. Mytilus edulis. Ecology 80, 1–14 (1999).
Dodson, S. I. The ecological role of chemical stimuli for the zooplankton: predator-induced morphology in Daphnia. Oecologia 78, 361–367 (1989).
Howe, G. A. & Jander, G. Plant immunity to insect herbivores. Annu. Rev. Plant Biol. 59, 41–66 (2008).
Quintana, X. D. et al. Predation and competition effects on the size diversity of aquatic communities. Aquat. Sci. 77, 45–57 (2015).
Sterner, R. W. in Plankton ecology (ed Sommer, U. ) Ch. 4, 107–170 (Springer, 1989).
Lürling, M. Phenotypic plasticity in the green algae Desmodesmus and Scenedesmus with special reference to the induction of defensive morphology. Ann. Limnol.-Int. J. Lim. 39, 85–101 (2003).
Van Donk, E., Ianora, A. & Vos, M. Induced defences in marine and freshwater phytoplankton: a review. Hydrobiologia 668, 3–19 (2011).
Lehman, J. T. in Growth and reproductive strategies of freshwater phytoplankton (ed Sandgren, C. D. ) Ch. 9, 369–387 (Cambridge University Press, 1988).
Hessen, D. O. & Van Donk, E. Morpholigical changes in Scenedesmus induced by substances released from Daphnia. Arch. Hydrobiol. 127, 129–140 (1993).
Lürling, M. & Van Donk, E. Zooplankton-induced unicell-colony transformation in Scenedesmus acutus and its effect on growth of herbivore Daphnia. Oecologia 108, 432–437 (1996).
Lürling, M. & Van Donk, E. Grazer-induced colony formation in Scenedesmus: are there costs to being colonial? Oikos 88, 111–118 (2000).
Marañón, E. Phytoplankton size structure. in Elements of Physical Oceanography: A derivative of the Encyclopedia of Ocean Sciences (eds Steele, J. H., Thorpe, A. S., & Turekian, K. K. ) 85 (Academic Press, 2009).
Reynolds, C. S. Ecology of phytoplankton. (eds Usher, M. & Saunders, D. ) Vol. 535 (Cambridge University Press, 2006).
Kenitz, K., Williams, R. G., Sharples, J., Selsil, Ö. & Biktashev, V. N. The paradox of the plankton: species competition and nutrient feedback sustain phytoplankton diversity. Mar. Ecol. Prog. Ser. 490, 107–119 (2013).
Edwards, K. F., Thomas, M. K., Klausmeier, C. A. & Litchman, E. Light and growth in marine phytoplankton: allometric, taxonomic and environmental variation. Limnol. Oceanogr. 60, 540–552 (2015).
Edwards, K. F., Litchman, E. & Klausmeier, C. A. Functional traits explain phytoplankton community structure and seasonal dynamics in a marine ecosystem. Ecol. Lett. 16, 56–63 (2013).
Holt, R. D., Grover, J. & Tilman, D. Simple rules for interspecific dominance in systems with exploitative and apparent competition. Am. Nat. 144, 741–771 (1994).
Strauss, S. Y., Rudgers, J. A., Lau, J. A. & Irwin, R. E. Direct and ecological costs of resistance to herbivory. Trends Ecol. Evol. 17, 278–285 (2002).
Yokota, K. & Sterner, R. W. Trade-offs limiting the evolution of coloniality: ecological displacement rates used to measure small costs. Proc. R. Soc. B: Biol. Sci. 278, 458–463 (2011).
Yoshida, T., Hairston, N. G. & Ellner, S. P. Evolutionary trade–off between defence against grazing and competitive ability in a simple unicellular alga, Chlorella vulgaris. Proc. R. Soc. B: Biol. Sci. 271, 1947–1953 (2004).
Harel, M. et al. Interactions between Scenedesmus and Microcystis may be used to clarify the role of secondary metabolites. Environ. Microbiol. Rep. 5, 97–104 (2013).
Lürling, M. & Roessink, I. On the way to cyanobacterial blooms: Impact of the herbicide metribuzin on the competition between a green alga (Scenedesmus) and a cyanobacterium (Microcystis). Chemosphere 65, 618–626 (2006).
Żak, A., Musiewicz, K. & Kosakowska, A. Allelopathic activity of the Baltic cyanobacteria against microalgae. Estuar. Coast. Shelf Sci. 112, 4–10 (2012).
Ishida, K. & Murakami, M. Kasumigamide, an antialgal peptide from the cyanobacterium Microcystis aeruginosa. J. Org. Chem. 65, 5898–5900 (2000).
Sukenik, A. et al. Inhibition of growth and photosynthesis of the dinoflagellate Peridinium gatunense by Microcystis sp. (cyanobacteria): a novel allelopathic mechanism. Limnol. Oceanogr. 47, 1656–1663 (2002).
Leão, P. N., Vasconcelos, M. T. S. & Vasconcelos, V. M. Allelopathy in freshwater cyanobacteria. Crit. Rev. Microbiol. 35, 271–282 (2009).
Agrawal, M. K. et al. Characterization of proteases in guts of Daphnia magna and their inhibition by Microcystis aeruginosa PCC 7806. Environ. Toxicol. 20, 314–322 (2005).
Yang, Z., Kong, F. X., Shi, X. L. & Cao, H. Morphological response of Microcystis aeruginosa to grazing by different sorts of zooplankton. Hydrobiologia 563, 225–230 (2006).
Yang, Z. & Li, J. J. Effects of Daphnia-associated infochemicals on the morphology and growth of Scenedesmus obliquus and Microcystis aeruginosa. J. Freshwater Ecol. 22, 249–253 (2007).
Gliwicz, Z. M. Why do cladocerans fail to control algal blooms? Hydrobiologia 200, 83–97 (1990).
Sarnelle, O. Herbivore effects on phytoplankton succession in a eutrophic lake. Ecol. Monogr. 63, 129–149 (1993).
Agrawal, A. A. Algal defense, grazers and their interactions in aquatic trophic cascades. Acta Oecol. 19, 331–337 (1998).
van Donk, E., Lürling, M. & Lampert, W. Consumer-induced changes in phytoplankton: inducibility, costs, benefits and the impact on grazers. in The Ecology and Evolution of Inducible Defenses (eds Tollrian, R. & Harvell, C. D. ) 89–103 (Princeton University Press, 1999).
Verschoor, A. M., Bekmezci, O. K., Ellen, V. & Vijverberg, J. The ghost of herbivory past: slow defence relaxation in the chlorophyte Scenedesmus obliquus. J. Limnol. 68, 327–335 (2009).
Clark, C. W. & Harvell, C. D. Inducible defenses and the allocation of resources: a minimal model. Am. Nat. 139, 521–539 (1992).
Miyazaki, T., Tainaka, K.-i., Togashi, T., Suzuki, T. & Yoshimura, J. Spatial coexistence of phytoplankton species in ecological timescale. Popul. Ecol. 48, 107–112 (2006).
Helmus, M. R., Mercado-Silva, N. & Vander Zanden, M. J. Subsidies to predators, apparent competition and the phylogenetic structure of prey communities. Oecologia 173, 997–1007 (2013).
Menden-Deuer, S. & Rowlett, J. Many ways to stay in the game: individual variability maintains high biodiversity in planktonic microorganisms. J. R. Soc. Interface 11, 20140031 (2014).
Sultan, S. E. Phenotypic plasticity for plant development, function and life history. Trends Plant Sci. 5, 537–542 (2000).
Persson, L. Behavioral response to predators reverses the outcome of competition between prey species. Behav. Ecol. Sociobiol. 28, 101–105 (1991).
Heil, M. Ecological costs of induced resistance. Curr. Opin. Plant Biol. 5, 345–350, (2002).
Karban, R., Agrawal, A. A. & Mangel, M. The benefits of induced defenses against herbivores. Ecology 78, 1351–1355 (1997).
Rodolfi, L. et al. Microalgae for oil: Strain selection, induction of lipid synthesis and outdoor mass cultivation in a low‐cost photobioreactor. Biotechnol. Bioeng. 102, 100–112 (2009).
Lürling, M. The effect of substances from different zooplankton species and fish on the induction of defensive morphology in the green alga Scenedesmus obliquus. J. Plankton Res. 25, 979–989 (2003).
Lampert, W., Rothhaupt, K. O. & Von Elert, E. Chemical induction of colony formation in a green alga (Scenedesmus acutus) by grazers (Daphnia). Limnol. Oceanogr. 39, 1543–1550 (1994).
Yang, Z. et al. Aggregate formation and polysaccharide content of Chlorella pyrenoidosa Chick (Chlorophyta) in response to simulated nutrient stress. Bioresour. Technol. 101, 8336–8341 (2010).
Grover, J. P. Dynamics of competition among microalgae in variable environments: experimental tests of alternative models. Oikos 62, 231–243 (1991).
Passarge, J., Hol, S., Escher, M. & Huisman, J. Competition for nutrients and light: stable coexistence, alternative stable states, or competitive exclusion? Ecol. Monogr. 76, 57–72 (2006).
Acknowledgements
This study was supported by the National Natural Science Foundation of China (31470508) and the Priority Academic Program Development of Jiangsu Higher Education Institutions.
Author information
Authors and Affiliations
Contributions
Z.Y. conceived this study; X.X.Z., J.W., Y.C.L. and Q.W.C. designed and performed the experiments; Z.Y., X.X.Z. and J.W. conducted the data analysis and prepared figures and tables. Z.Y. and X.X.Z. wrote the manuscript. All of the authors reviewed and approved the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Zhu, X., Wang, J., Lu, Y. et al. Grazer-induced morphological defense in Scenedesmus obliquus is affected by competition against Microcystis aeruginosa. Sci Rep 5, 12743 (2015). https://doi.org/10.1038/srep12743
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep12743
This article is cited by
-
Long-term continuous mismatch between grazing cues and real grazing losses causes attenuation of induced morphological defense in Scenedesmus
Journal of Applied Phycology (2024)
-
Overexpression of a novel gene (Pt2015) endows the commercial diatom Phaeodactylum tricornutum high lipid content and grazing resistance
Biotechnology for Biofuels and Bioproducts (2022)
-
Succession of Aphanizomenon flos-aquae and Microcystis aeruginosa in direct co-culture experiments at different temperatures and biomasses
Journal of Oceanology and Limnology (2022)
-
Indole-3-acetic-acid-induced phenotypic plasticity in Desmodesmus algae
Scientific Reports (2018)
-
Magnesium depletion suppresses the anti-grazer colony formation in Scenedesmus obliquus
Environmental Science and Pollution Research (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
