
Abstract
Gelatinous organisms apparently play a central role in deep pelagic ecosystems, but lack of observational methodologies has restricted information on their behaviour. We made acoustic records of diel migrating jellyfish Periphylla periphylla forming small, ephemeral groups at the upper fringe of an acoustic scattering layer consisting of krill. Groups of P. periphylla were also documented photographically using a remotely operated vehicle (ROV). Although the adaptive value of group formation remains speculative, we clearly demonstrate the ability of these jellyfishes to locate and team up with each other.
Similar content being viewed by others


Introduction
Deep pelagic habitats are characterized by dark waters, dilute populations, few conspecifics and scarce food and animals evidently require unique physiological and behavioural adaptations to succeed in this realm1. From the use of manned submersibles and remotely operated vehicles (ROVs) it has become increasingly apparent that gelatinous forms play a central role in deep pelagic ecosystems1,2. Yet, with the low concentrations of organisms present only scattered observations exist and lack of observational methodologies generally restricts behavioural information from this inaccessible part of the ocean.
In Lurefjorden, Norway, the cosmopolitan mesopelagic jellyfish Periphylla periphylla occurs in concentrations several orders of magnitude higher and at shallower depths than in its normal oceanic habitat3,4,5, offering unmatched opportunities to study behavioural processes, that are otherwise difficult to observe in deep water jellyfish. The high abundance in Lurefjorden has been ascribed to the particularly high light extinction of the water in this basin, which may have excluded visually foraging mesopelagic fish from the dark, deep waters6,7. This in turn leaves a surplus of prey for the jellyfish6,8.
The use of echo sounders enables the observation of individual jellyfish in situ and acoustic studies have documented strikingly diverse behavioural patterns among P. periphylla, with the population segregating into assemblages with distinctly different vertical distribution and diel vertical migration (DVM) behaviour5. Individuals constituting the shallow-most of these assemblages (behavioural Mode 1, sensu 5) inhabit the upper 100 m, where they carry out synchronous DVM. These are the jellyfish addressed in this study. Nocturnal groups of P. periphylla have previously been reported at the surface and ascribed to reproduction, as seeking the two dimensional surface would increase the chance of encounter and mating compared to the three dimensional water column9. We here show that P. Periphylla readily teams up also at depth, forming ephemeral groups of a few individuals that may stay together for 10’s of minutes.
Results
Acoustic studies
2006: Acoustic studies were made one afternoon using the echosounders of the RV “Trygve Braarud” and a submerged echosounder, kept at 50 m most of the registration period. The 38 kHz echosounder of the RV revealed individual targets (jellyfish; see Methods) below ~40 m, with a core of particularly strong echoes at 60–70 m. More diffuse echoes (krill) appeared below (Fig. 1). The echogram from the RV also displays the trace of the submerged echosounder. As the dielly migrating targets ascended towards the surface, the core of dense registrations passed the submerged echo sounder (Fig. 1).

Acoustic records made from RV “Trygve Braarud”s echosounder (38 kHz) in the afternoon 12 October 2006.
The echogram depicts targets ascribed to dielly migrating Periphylla periphylla at the upper fringe of a migrating scattering layer of krill. A submerged echosounder is seen as a horizontal line at 50 m depth during most of the registration period. Colour scale refers to backscattering strength (SV), with brownish-red representing the strongest echoes.
High resolution data from the submerged echo sounder (200 kHz) show that acoustic targets (jellyfish) in the core of the layer occurred pair-wise, or in small groups (Fig. 2). This likely explains the strong echoes from the hull mounted transducer, which has too coarse vertical resolution to distinguish between the individuals in these asssemblages, thereby giving added echo strength. On several occasions individuals could be observed as they actively joined already existing groups (Fig. 2b,c). In such cases, an individual ascended from below and positioned itself aligned beneath another one (Fig. 2d). The acoustic data suggest that one jellyfish could detect a neighbour from at least 2 m below. This vertical distance is suggested from the observations of individuals, which abruptly changed their behaviour and rapidly ascended when passing beneath another speciemen at this range (Fig. 2b). The two ascending individuals displayed in Fig. 2b swam vertically at speeds of ~17 and 18 cm s−1 (derived from the echogram).

Acoustic records made from a 200 kHz echosounder, deployed at 50 m depth in Lurefjorden on 12 October 2006. Each echo trace is ascribed to Periphylla periphylla. a) 3 groups of closely spaced individuals b) 2 individuals ascending towards a jellyfish above, forming a group of 3 individuals c) Group of 2 individuals being joined by 2 more individuals, forming a group of 4 d) Same group as in “c”, but in 3-D, showing that individuals align just beneath each other. Colour scale for “a-c” refers to backscattering strength (SV); colour scale for “d” refers to individual target strength (Ts), with brownish-red representing the strongest echoes in both cases.
Groups were detected as these acoustic targets were ascending towards the submerged echosounder in the afternoon, but not after the strong core of the vertically migrating SL had migrated past the echosounder. However, as these jellyfish approached the surface in the evening, corresponding behaviour was suggested also by the vessels’ 120 kHz echo sounder, which has a narrower beam and shorter pulse length than the 38 kHz echosounder, providing better resolution. These observations included a few examples of individuals which appeared to join groups from below and for a few meters ascended with a speed of ~10 cm sec−1, thereafter aligning themselves below the other members (not shown). While individuals evidently changed their behaviour to form/join a group, the groups appeared to follow the same DVM patterns as for single individuals (cf. Fig. 1).
2010. Based on the results from 2006, jellyfish in the upper 100 m appeared to be the ones forming groups. Therefore, an upward-looking echosounder (200 kHz) was deployed in a buoy floating at 100 m depth. This depth was selected to obtain high resolution data for jellyfish with expected daytime distribution at 50–100 m. Group behaviour was repeatedly recorded during a 7 days registration period from this upward-looking echo sounder. Even with the short pulse length applied at 200 kHz (0.128 ms; vertical resolution 2,4 cm), individuals in groups could for most of the time not be resolved as the acoustic targets (jellyfish) stayed too tightly together. Groups appeared as stronger and slightly more vertically extended echo traces than individual jellyfish, but occasionally sufficient separation between individuals forming a group made it possible to detect them separately (Figs 3,4).

Two individuals swimming together, subsequently splitting up.
Records are from a stationary, upward looking 200 kHz echosounder, floating in a buoy at 100 m depth, October 2010. Results are visualized in a normal echogram (a) and in 3-D based on acoustic target tracking (b). Colour scale for “a” refers to backscattering strength (SV) and colour scale for “b” refers to individual target strength (Ts), with brownish-red representing the strongest echoes in both cases.

Examples of groups recorded from the stationary upward looking 200 kHz echosounder, October 2010.
Examples are from descent in the morning, early day, late day and ascent in the afternoon. Arrows depict examples of group behaviour; arrows in “d” suggest the formation and splitting of a group of two individuals. Colour scale refers to backscattering strength (SV), with brownish-red representing the strongest echoes.
The acoustic data contained examples of both group formation and separation (Figs 3,4). We could not decide any typical “life span” of a group, as groups would normally drift by and not spend sufficiently long time in the acoustic beam to assess both their formation and separation. Therefore, the duration of observations would rather reflect current speed than jellyfish behaviour. The maximum duration in our data was ~20 minutes (until the group drifted out of the acoustic beam). However, also short durations were detected, with individuals joining and separating after a few minutes (Fig. 4).
In total, 152 groups were observed during the registration period, while ~4000 echo traces of single individuals were recorded in the course of the same period. This suggests that ~10% of the jellyfish in the upper part of the water column were engaged in group behaviour, as each group normally appeared to consist of 2–3 individuals. Groups were observed during descent in the morning, daytime and ascent in the afternoon. No groups were observed at night.
Visual observations
In February 2010, direct visual observations of groups were made at the surface, as well as with the ROV which documented that P. periphylla would form groups also at depth (Fig. 5). In total 5 pairs of individuals with their tentacles entangled were recorded with the ROV. Quantifications of size or distances were not possible, thus any other forms of group behaviour (jellyfish swimming close to each other without physical contact) could not be adequately assessed. Yet it was evident that the vast majority of the filmed jellyfish occurred as single individuals during these records in winter.

Pictures of a pair of Periphylla periphylla (from ROV video) taken 12 sec apart (109 m depth).
Discussion
The current study has documented apparent social behaviour among a jellyfish. The observations of group-forming jellyfish adds to the conspicuously varied individual behaviour within this population5,10 and in jellyfish at large (see below). Tiemann et al. 9 made visual observations of clusters of P. periphylla at the surface at night, with individuals on occasions having their tentacles entangled, as also observed – but not being the focus - in the current study. Here we show that individuals team up also at depth.
The behavioural observations support that the encounters are not entanglements of chance, but active group forming. Jellyfish are basal metazoans and thought to have very limited sensory capabilities (but see11,12) and never thought to have social interactions. The findings reported here challenge our current notions about jellyfish behaviour and raise questions about the cue(s) that the jellyfish use to seek each other out at depth as well as about the adaptive values of group behaviour. In our observations of individuals joining another individual, or an already existing group, individuals joined from below in daylight or at dusk. The reaction distance would be at least ~2 m, derived from the observed behavioural changes in response to other jellyfish/groups at this range (e.g. Fig. 2). Recognizable physical disturbance from a swimming jellyfish is on the scales of cm’s13 and is not likely to extend several meters. The approaching Periphylla did not seem to follow the trail of its conspecific (Fig. 2), as seems to be the case among small plankton following chemical tracks14, but rather ascended abruptly when passing below upcoming partners.
Many jellyfish are bioluminecent, which is traditionally believed to be a warning to potential predators15,16. Jarms et al.15 assumed that bioluminescence as part of social interactions was unlikely in P. periphylla, because of its simple photoreceptors. However, the wide repertoire of bioluminescent displays of P. periphylla16 may indicate multiple functions and many other deep sea animals use bioluminescence for intraspecific communication17,18. One way of testing for intraspecific bioluminescent communication in jellyfish in situ might be to record their various bioluminescent signals in the laboratory and display these to the jellyfish in the field, studing their response by means of a stationary, submerged echosounder.
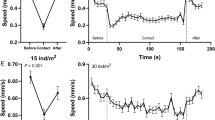
Bioluminescence functions best in darkness19 and our registrations were in the upper part of the water column during times with surface light. Alternatively, the jellyfish might locate the shadow/silhouettes of conspecifics towards the surface. When detecting a conspecific, individuals abruptly ascended to join a jellyfish directly above (Fig. 2), the ascent speed in such cases being about 5–10 times their normal vertical swimming speed20.
Many marine animals with dilute populations have sophisticated adaptations for locating conspecifics, often for mating. Tiemann et al.9 speculated that P. periphylla migrating to the surface would increase mate encounter when moving from 3 to 2 dimensions and that this behaviour was developed in response to low abundances in oceanic environments. However, we here show that surface encounters are not a necessity to seek out conspecifics. While mating is a plausible function, alternative explanations may apply. Also, groups forming at depths are not necessarily equivalent to those forming at the surface. Likewise, motivation for teaming up when faced with high concentrations of prey may differ from motivation outside of prey patches, or when faced with predators.
The records of group-forming jellyfish were made at the upper fringe of an acoustic scattering layer of the krill Meganyctiphanes norvegica, which is known as prey for P. periphylla in Lurefjorden4,21. Predators of various taxa may accumulate in respons to local prey aggregations (e.g.22). These may not be social aggregations as such but simply a response to a common cue related to e.g. prey density. However, while jellyfish may have accumulated at the top of the krill layer for favourable feeding conditions, the very close associations and coherent behaviour of the groups do not support that plain aggregation in prey patches would be the explanation for the group behaviour reported here.
Jellyfish swimming behaviour is often related to feeding strategy (e.g.21,23,24). As tactile predators, jellyfish rely on collisions with prey. Encounters are mediated by predator, prey25, or water motion26. P. periphylla swims slowly with its tentacles first, allowing the jellyfish to approach prey with a minimum amount of fluid disturbance caused by swimming13,21. Nevertheless, escapement is prevalent when foraging on agile organisms. For example, krill jump in random directions in response to contact with tentacular predators27. Prey escapes from one predator may therefore enhance the chance of capture by its neighbour and vice versa, thus benefiting both individuals in the long run. To evaluate if instantaneous prey escapes - elicited by one predator – enhance the success of a neighbour, we formulated a probabilistic model of cooperation (Table 1). That exercise suggests that hunting in groups indeed may be profitable. However, the fact that groups are ephemeral and vary temporally and with depth may suggest that there also are significant costs involved with this behaviour.
Regardless of the adaptive significance, our data imply active social behaviour among a jellyfish that normally lives in the deep sea and that they have ways of locating and finding each other in the open water masses. Although jellyfish are simple organisms, with poorly developed sensory capabilities and neural networks, jellyfish may display well-developed behavioural repertoires28,29,30. Horizontal migrations guided by the position of the sun have been recorded in several species28,31, vertical migration may prevent tidal dispersal29 and recent studies of jellyfish using electronic tracking tags have revealed directional swimming related to current flows that may help in bloom maintenance in apparent favourable habitats32. Such direct tracking of jellyfish has also unveiled vertical movements that seem linked to searching the water column for prey and that jellyfish can search the water column like fish33. Our results on social behavior of P. periphylla add to the emerging pattern that jellyfish are not simply passive drifters but rather display active and varied behavioural repertoirs of adaptive significance.
Methods
Results in this paper are based on observations from 3 cruises to Lurefjorden. In October 2006, small groups of acoustic targets interpreted as Periphylla periphylla were observed during their ascent in the afternoon. During a cruise in February 2010 the primary aim was to film groups of P. periphylla using an ROV. During this cruise we also made abundant observations of P. periphylla forming groups at the surface at night (cf.9). In September/October 2010 we conducted more comprehensive acoustic studies to further document the group behaviour. Additional video footages from ROV were made 31 October 2011; abundant records of single individuals were made, but no groups were observed during that field effort, which is not further referred to here.
Acoustic studies
October 2006
The studies were carried out during the field campaign described by5. Results presented here were obtained while the 22 m RV Trygve Braarud (University of Oslo) was freely drifting in the calm fjord waters. Acoustic records were made with the ships’ hull mounted Simrad EK500 38 kHz (11.8° opening angle) and 120 kHz (7°) echo sounders and with a submerged Simrad EK60, 200 kHz (7°) cabled to a PC on board. The EK60 was deployed at various depths down to 150 m, with measurements both day and night. This comprised measurements above and below a relatively dense scattering layer observed by the ships’ echosounder. This layer passed the submerged echosounder at 50 m depth when ascending in the afternoon (Fig. 1). The submerged echosounder was operated with a short pulse length (0.128 ms) for the highest possible vertical resolution (2.4 cm).
September/October 2010
An upward-looking Simrad EK60, 200 kHz echosounder was deployed in a buoy floating at 100 m depth. This depth was selected to obtain high resolution data for jellyfish with expected daytime distribution at 50–100 m. The buoy was anchored to the bottom (near the location of the acoustic studies by5 and the echosounder was cabled to shore for power and transfer of data during the deployment (27 September to 3 October). The echo sounder was operated at short pulse length (128 ms) with a ping rate of 2 s−1. From these acoustic records we documented the formation of groups and roughly quantified the fraction of P. periphylla engaging in groups.
Upward-facing echosounders cabled to shore were also located at 200 m (120 kHz) and at the bottom (290 m; 38 kHz). Echograms were visually inspected without detecting apparent groups of P. periphylla in the deeper part of the water column covered by these two moorings.
Processing acoustic data
The presence of groups was assessed by visually scrutinizing echograms made using the software Sonar 5 Pro version 6.0.034. This software was also applied for acoustic target tracking (TT) and for assessing target strength (TS), the latter being a proxy for size. TT groups individual echoes into tracks utilising information on the proximity of sequential echoes in determining the tracks so that a particle can be followed both horizontally and vertically as it moves through the acoustic beam35. Presentations of these 3-D data are based on smoothed data (linear interpolation). Echograms for the figures were made in Matlab.
Video and photographic records
Video records were made in February 2010. Vertical casts throughout the water column were carried out with the ROV “Aglantha”, equipped with cameras for both normal and red light, the results reported here from use of the latter.
Sampling
Trawling for identification of acoustic targets was done in 2006; P. periphylla completely dominated trawl catches by weight at 50–100 m5. The prevalence of Periphylla accords with numerous studies from Lurefjorden3,4,36 as well as with ROV records in this study.
Additional Information
How to cite this article: Kaartvedt, S. et al. Social behaviour in mesopelagic jellyfish. Sci. Rep. 5, 11310; doi: 10.1038/srep11310 (2015).
References
Robison, B. H. Deep pelagic biology. J Exp Mar Biol Ecol 300, 253–272 (2004).
Sutton, T. T. Vertical ecology of the pelagic ocean: classical patterns and new perspectives. J Fish Biol 3, 1508–1527 (2013).
Fosså, J. H. Mass occurrence of Periphylla periphylla (Schypozoa, Coronatae) in a Norwegian fjord. Sarsia 83, 103–116 (1992).
Youngbluth, M. J. & Båmstedt, U. Distribution, abundance, behavior and metabolism of Periphylla periphylla, a mesopelagic coronate medusa in a Norwegian fjord. Hydrobiol 451, 321–333 (2001).
Kaartvedt, S., Titelman, J., Røstad. A. & Klevjer, T. A. Beyond the average: Diverse individual migration patterns in a population of mesopelagic jellyfish. Limnol Oceanogr 56, 2189–2199 (2011).
Eiane, K., Aksnes, D. L., Bagøien, E. & Kaartvedt, S. Fish or jellies - a question of visibility? Limnol Oceanogr 44, 1352–1357 (1999).
Aksnes, D. L., Nejstgaard, J., Sædberg, E. & Sørnes, T. Optical control of fish and zooplankton populations. Limnol Oceanogr 49, 233–238 (2004).
Bagøien, E., Kaartvedt, S., Aksnes, D. L. & Eiane, K. Vertical distribution and mortality of overwintering Calanus. Limnol Oceanogr 46, 1494–1510 (2001).
Tiemann, H., Sötje, I., Johnston, B. D., Flood, P. R. & Båmstedt, U. Documentation of potential courtship-behaviour in Periphylla periphylla (Cnidaria: Scyphozoa). J Mar Biol Ass UK 89, 63–66 (2009).
Ugland, K. I., Aksnes, D. L., Klevjer, T. A., Titelman, J. & Kaartvedt, S. Lévy night flights by the jellyfish Periphylla periphylla. Mar Ecol Prog Ser 513, 121–130 (2014).
Nilsson, D. E., Coates, M. M., Gislén, I., Skogh, C. & Garm, A. Advanced optics in a jellyfish eye. Nature 435, 201–205 (2005).
Garm, A., O’Connor, M., Parkefelt, L. & Nilsson, D.-E. Visually guided obstacle avoidance in the box jellyfish Tripedalia cystophora and Chiropsella bronzie. J Exp Biol 210, 3616–3623 (2007).
Raskoff, K. A. Foraging, prey capture and gut contents of the mesopelagic narcomedusa Solmissus spp. (Cnidaria: Hydrozoa). Mar Biol 141, 1099–1107 (2002).
Kiørboe, T. & Bagøien, E. Motility patterns and mate encounter rates in planktonic copepods. Limnol Oceanogr 50, 1999–2007 (2005).
Jarms, G., Tiemann, H. & Båmstedt, U. Development and biology of Periphylla periphylla (Schypozoa, Coronatae) in a Norwegian fjord. Mar Biol 141, 647–657 (2002).
Herring, P. J. & Widder, E. A. Bioluminescence of deep-sea coronate medusa. Mar Biol 146, 39–51(2004).
Widder, E. A. Bioluminescence in the ocean: origins of biological, chemical and ecological diversity. Science 328, 704–708 (2010).
Haddock, S. H. D., Moline, M. A. & Case, J. F. Bioluminescence in the sea. Annu Rev Mar Sci 2, 443–493 (2010).
Warrant, E. J. & Locket, N. A. Vision in the deep sea. Biol Rev 79, 671–712 (2004).
Kaartvedt, S., Klevjer, T. A., Torgersen, T, Sørnes, T. A. & Røstad, A. Diel vertical migration of individual jellyfish (Periphylla periphylla). Limnol Oceanogr 52, 975–983 (2007).
Sørnes, T. A., Hosia, A., Båmstedt, U. & Aksnes, D. L. Swimming and feeding in Periphylla periphylla (Scyphozoa, Coronatae). Mar Biol, 153, 653–659 (2008).
Sims, D. W. et al. Scaling laws of marine predator search behaviour. Nature 451, 1098–1103 (2008).
Costello, J. H. & Colin, S. P., Flow and feeding by swimming scyphomedusae. Mar Biol 124, 339–406 (1995).
Colin, S. P. & Costello, J. H. Prey resource use by coexistent hydromedusae from Friday Harbor, Washington. Limnol Oceanogr 47, 934–942 (2002).
Gerritsen, J. & Strickler, J. R. Encounter probabilities and community structure in zooplankton: a mathematical model. J Fish Res Board Can 34, 73–82 (1977).
Kiørboe, T. & Saiz, E. Planktivorous feeding in calm and turubulent environments, with emphasis on copepods. Mar Ecol Prog Ser 122, 135–145 (1995).
Hamner, W. M. & Hamner, P. P. Behavior of Antarctic krill (Euphausia superba): schooling, foraging and antipredatory behavior. Can J Fish Aquat Sci 57 (Suppl 3): 192–202 (2002).
Hamner, W. M., Hamner, P. P. & Strand, S. W. Sun-compass migration by Aurelia aurita (Scyphozoa): population retention and reproduction in Saanich Inlet, British Columbia. Mar Biol 119, 347–356 (1994).
Albert, D. J. Aurelia labiata medusae (Scyphozoa) in Roscoe Bay avoid tidal dispersion by vertical migration J Sea Res 57, 281–287 (2007).
Albert, D. J. What’s on the mind of a jellyfish? A review of behavioural observations on Aurelia sp. jellyfish. Neurosci Biobehav R 35, 474–482 (2011).
Hamner, W. M. & Hauri, I. R. Long-distance horizontal migrations of zooplankton (Scyphomedusae: Mastigias). Limnol Oceanogr 26, 414–423 (1981).
Fossette, S. et al. Current-oriented swimming by jellyfish and its role in bloom maintenance. Current Biology 25, 342–347 (2015).
Hays, G. C. et al. High activity and Lévy searches: jellyfish can search the water column like fish. Proc R Soc B 279, 465–473 (2012).
Balk, H. & Lindem, T. Sonar 4 and Sonar 5-Pro Post processing systems. Operator manual 6.0.1. http://folk.uio.no/hbalk/sonar4_5/index.htm. Lindem Data Acquisition (2012). Date of access: 01/03/2014.
Brede, R., Kristensen, F. H., Solli, H. & Ona, E. Target tracking with a split-beam echo sounder. Rapp P-V Reùn Cons Int Explor Mer 189, 254–263 (1990).
Båmstedt, U., Kaartvedt, S. & Youngbluth, M. An evaluation of acoustic and video methods to estimate the abundance and vertical distribution of jellyfish. J Plankton Res 25, 1307–1318 (2003).
Acknowledgements
The study was funded by King Abdullah University of Science and Technology (KAUST) and the University of Oslo.
Author information
Authors and Affiliations
Contributions
SK conceived the research. SK, TAK, AR, IS and JT contributed during field work. SK, TAK and AR conducted the acoustic studies. IS analysed the acoustic results and AR and TAK made the figures. JT conducted the video analyses. KIU developed the model. SK, KIU and JT wrote the paper.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Kaartvedt, S., Ugland, K., Klevjer, T. et al. Social behaviour in mesopelagic jellyfish. Sci Rep 5, 11310 (2015). https://doi.org/10.1038/srep11310
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep11310
This article is cited by
-
Self-healing electronic skins for aquatic environments
Nature Electronics (2019)
-
Increased occurrence of the jellyfish Periphylla periphylla in the European high Arctic
Polar Biology (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
