
Abstract
Neuroendocrine cells store ATP in secretory granules and release it along with hormones that may trigger a variety of cellular responses in a process called purinergic chemical transmission. Although the vesicular nucleotide transporter (VNUT) has been shown to be involved in vesicular storage and release of ATP, its physiological relevance in vivo is far less well understood. In Vnut knockout (Vnut−/−) mice, we found that the loss of functional VNUT in adrenal chromaffin granules and insulin granules in the islets of Langerhans led to several significant effects. Vesicular ATP accumulation and depolarization-dependent ATP release were absent in the chromaffin granules of Vnut−/− mice. Glucose-responsive ATP release was also absent in pancreatic β-cells in Vnut−/− mice, while glucose-responsive insulin secretion was enhanced to a greater extent than that in wild-type tissue. Vnut−/− mice exhibited improved glucose tolerance and low blood glucose upon fasting due to increased insulin sensitivity. These results demonstrated an essential role of VNUT in vesicular storage and release of ATP in neuroendocrine cells in vivo and suggest that vesicular ATP and/or its degradation products act as feedback regulators in catecholamine and insulin secretion, thereby regulating blood glucose homeostasis.
Similar content being viewed by others


Introduction
Secretory granules are key cellular organelles in the secretory processes. They contain various neurotransmitters, hormones, enzymes and other products that are actively discharged from the cells through exocytosis1,2,3,4,5. Secretory granules contain the molecular machinery for the vesicular storage of neurotransmitters, which are similar to those found in synaptic vesicles, such as a vacuolar proton pump as the primary pump and secondary active transporters for the various neurotransmitters1,2,3,4,5. One of the particular and common properties is the relatively high concentration of nucleotides in secretory granules1,2,3,4. For example, bovine adrenal chromaffin granules, rat β-cell insulin granules and pig platelet dense granules contain ~120, ~12 and ~740 mM nucleotides, respectively1,2,3. When vesicular ATP is co-released with the internal vesicular constituents, extracellular ATP and its degradation products may act as intercellular signaling molecules in a paracrine or autocrine-mediated manner by binding various purinoceptors on the target cells6.
There are two types of purinoceptors: ligand-gated ion channel type P2X and G protein-coupled P2Y receptors6. In adrenal chromaffin cells, the P2Y12 receptor mediates inhibition of the exocytotic release of catecholamine by modulation of voltage-operated Ca2+ channels7,8,9. In contrast, several studies have shown that stimulation of P2Y2 receptor enhances inositol phosphate formation and increases exocytotic release of catecholamine in adrenal chromaffin granules10,11,12. P2X4-7 receptors are also known to function in chromaffin cells and their stimulation by specific agonists may stimulate catecholamine secretion by causing the opening of a nonselective cation channel10,11,13. In pancreatic β-cells, P2X1, P2X3 and P2Y1 receptors have been identified whose stimulation by agonists causes increased insulin secretion in humans14,15,16,17. In contrast, P2Y1 knockout (−/−) mice exhibit enhanced glucose-dependent insulin secretion from pancreatic β-cells, suggesting an inhibitory role of this protein for insulin secretion in mouse18. In platelets, the P2X1 receptor is responsible for rapid Ca2+ influx and platelet shape change in response to ATP release, leading to platelet activation induced by low concentrations of collagen19. The P2Y1 and P2Y12 receptors are essential for normal aggregation in response to ADP19,20,21. Overall, the physiological output of secretory granule-mediated purinergic chemical transmission is rather complex and sometimes produces contradictory secondary responses related to time-dependent, location-dependent and species-dependent changes in the expression of purinoceptors. Therefore, to evaluate the physiological relevance of the purinergic chemical transmission in vivo, it is important to examine the release of ATP under controlled conditions.
The vesicular nucleotide transporter (VNUT) is an SLC17A9-encoded membrane protein that actively transports various nucleotides across the membrane using an electrochemical gradient of protons established by vacuolar proton ATPases22. VNUT localizes to adrenal chromaffin granules and synaptic vesicles22,23. The suppression of gene expression by RNA interference leads to a significantly decreased release of ATP22,23. Thus, it is believed that VNUT is involved in vesicular storage and subsequent secretion of ATP, although the impact of VNUT in these processes in vivo is not defined. In the present study, we therefore generated knockout mice that lack Vnut and studied the effect of the absence of VNUT on the vesicular storage and secretion of ATP from neuroendocrine cells. We show evidence that vesicular ATP acts as a feedback regulator in catecholamine and insulin secretion in vivo and thereby regulates blood glucose homeostasis.
Results
Generation and characterization of the Vnut−/− mice
We produced mice lacking Vnut by homologous recombination in mouse embryonic stem cells containing a genetic deletion of the first four transmembrane domains of VNUT (Fig. 1A). Disruption of the gene was confirmed by Southern blot analysis and PCR (Figs. 1B and C).

Generation of Vnut−/− mice.
(A) Targeted disruption of mouse Vnut gene. The Vnut locus containing exons 2–3 was replaced with a neomycin resistance gene by homologous recombination of the linearized targeting vector. The positions of probes for southern blotting analysis are shown. (B) Southern blotting analyses of genomic DNA from neomycin-resistant ES cell clones. Genomic DNA was digested by EcoT22I and NotI for 5′ probe (upper panel) and SpeI for 3′ probe (lower panel). The targeted allele yielded a 10.3-kb band using the 5′ probe and a 7.6-kb band using the 3′ probe. The full-length blotting images are presented in Supplementary Fig. S3. (C) PCR amplification of genomic DNA from the tails of wild-type (WT) and homozygous Vnut−/− (KO) mice. A 217 bp band was amplified from the wild-type allele and a 314 bp band was amplified from the recombinant allele. The full-length gel image is presented in Supplementary Fig. S4. (D) Western blot of crude brain synaptic vesicles (30 μg) prepared from WT and KO mice with the antibodies indicated. The full-length blotting images are presented in Supplementary Fig. S5. (E–G) Depolarization-dependent ATP (E), glutamate (F) and aspartate (G) release from hippocampal neuron of WT (open bars, n = 3) and KO mice (filled bars, n = 3) was assessed 20 min after the addition of 50 mM KCl (+). Error bars represent mean ± S.E. *: p < 0.05. NS: not significant.
Vnut−/− mice appeared to be normal when compared to wild-type mice in terms of weight gain, body size, morphology, food intake, water intake, oxygen consumption, carbon dioxide emission, respiratory exchange ratio (RER), locomotor activity and open field and plus maze behaviors (Figs. S1 and S2).
Release of ATP from brain neurons
The loss of VNUT protein in Vnut−/− mice was confirmed by immunoblotting of brain synaptic vesicles (Fig. 1D). Other vesicular neurotransmitter transporters, such as vesicular monoamine transporter 2 (VMAT2), vesicular glutamate transporter 1 (VGLUT1), VGLUT2, vesicular excitatory amino acid transporter (VEAT) and vesicular GABA transporter (VGAT), showed normal expression (Fig. 1D). Moreover, the expression levels of synaptic vesicle glycoprotein 2 (SV2), vesicle-associated membrane protein 2 (VAMP2), synaptophysin (Syn) and synaptotagmin (Syt) were unaffected by the deletion of VNUT, while expression of N-ethylmaleimide sensitive factor (NSF) and subunit A of the vacuolar proton ATPase (V-ATPase) were slightly different (Fig. 1D). Neurons from wild-type brains exhibited KCl-dependent release of glutamate, aspartate and ATP as reported previously23. Although Vnut−/− neuronal cells showed 30%–40% reductions in glutamate and aspartate release, they showed complete loss of ATP secretion (Figs. 1E–G).
Vesicular storage and release of ATP in adrenal chromaffin granules
Since the adrenal chromaffin granules are one of the most characterized ATP-containing secretory granules, we studied the role of VNUT in the storage and release of ATP in granules. Consistent with the loss of VNUT in the brain, VNUT immunoreactivity as well as mRNA expression were absent in the adrenal glands, while gene expression and protein levels of other chromaffin granule proteins including VMAT1 and V-ATPase were unaffected (Figs. 2A to C). Electron microscopy showed that there were no apparent morphological differences between wild-type and Vnut−/− mice (Fig. 2D).

Loss of VNUT impaired the vesicular accumulation and exocytosis of ATP in adrenal glands.
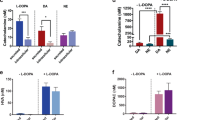
(A) Real-time PCR analysis was performed with total RNA isolated from adrenal glands of wild-type (WT, open bar, n = 3) and Vnut−/− mice (KO, filled bar, n = 3). The relative amount of mRNA was defined as the ratio to that of G3pdh. (B) Western blot of adrenal chromaffin membrane vesicles (5 μg) prepared from WT and KO mice and probed with the indicated antibodies. The full-length blotting images are presented in Supplementary Fig. S6. (C) Indirect immunofluorescence microscopy revealed that VNUT was expressed in WT mouse adrenal glands (WT). No VNUT immunoreactivity was seen in adrenal glands from KO mice (KO). C, cortex; M, medulla. Scale bar = 20 μm. (D) Electron microscopy indicated that adrenal chromaffin cells from KO mice were morphologically normal. A, adrenaline-storing cells; NA, noradrenaline storing cells. Scale bar = 1 μm. (E) ATP uptake by chromaffin granules isolated from WT mice (open bar, n = 3) and KO mice (filled bar, n = 3). The final concentrations of the substances tested were: Evans blue, 10 μM; bafilomycin A1, 50 nM; oligomycin, 5 μM; atractyroside, 10 μM. (F) Dopamine uptake by chromaffin granules isolated from WT (n = 3) and KO mice (n = 3). The dopamine uptake values in the presence of ATP were corrected by subtracting the values obtained in the absence of ATP. (G) The depolarization-dependent release of ATP from chromaffin cells of WT (n = 3) and KO mice (n = 3) was assessed 20 min after the addition of 50 mM KCl. (H) and (I) Depolarization-dependent release of noradrenaline (H) and adrenaline (I) from chromaffin cells of WT (n = 3) and KO mice (n = 3) after the addition of 50 mM KCl (+). When indicated, 50 mM KCl plus 0.1 mM ATP was included in the medium. (J) The amounts of noradrenaline and adrenaline in adrenal chromaffin granules of WT (n = 3) and KO mice (n = 3). (K) Serum levels of catecholamines in WT (n = 4) and KO (n = 4) mice after overnight fast. Error bars represent mean ± S.E. *: p < 0.05. NS: not significant.
We examined whether chromaffin granules prepared from wild-type and Vnut−/− mice accumulate ATP and dopamine. The membrane vesicles were incubated in buffer containing ATP and Mg2+ to establish an electrochemical gradient of protons across the membrane by V-ATPase, which provides the driving force for VNUT and VMAT. We found that the membrane vesicles of chromaffin granules from wild-type mice exhibited ATP uptake with properties similar to that of purified VNUT-containing proteoliposomes22. ATP uptake was sensitive to Evans blue (an inhibitor of VNUT) and bafilomycin A1 (an inhibitor of V-ATPase) and was insensitive to oligomycin (an inhibitor for oxidative phosphorylation) (Fig. 2E). In contrast, no ATP uptake was observed in the membrane vesicles of chromaffin granules from Vnut−/− mice (Fig. 2E). The same batch of the membrane vesicles from both wild-type and Vnut−/− mice exhibited appreciable and comparable reserpine-sensitive dopamine uptake (Fig. 2F). These results indicated that chromaffin granules of Vnut−/− mice selectively lost ability for vesicular ATP storage.
Subsequently we measured the release of ATP, noradrenaline (NA) and adrenaline (Ad), from cultured chromaffin cells of wild-type and Vnut−/− mice. As expected, appreciable amounts of ATP were released from the chromaffin cells of wild-type mice upon stimulation with high concentrations of K+ (depolarization), whereas no such stimulation was observed in the cells of Vnut−/− mice (Fig. 2G). Chromaffin cells from both wild-type and Vnut−/− mice exhibited depolarization-dependent NA and Ad release (Figs. 2H and I). The amount of NA and Ad released from chromaffin cells were significantly lower in Vnut−/− mice, although the ratio of stimulation of NA release by K+ was similar in cells from both wild-type and Vnut−/− mice (Figs. 2H and I). The addition of ATP increased the NA and Ad release from Vnut−/− chromaffin cells (Figs. 2H and 2I). The adrenal chromaffin granules of Vnut−/− mice contained significantly lower levels of NA and Ad than those from wild-type controls (Fig. 2J). However, the blood concentrations of NA and Ad were not significantly different between Vnut−/− and wilt-type mice (Fig. 2K).
Role of vesicular ATP in β−cells in the islet of Langerhans
Islet of Langerhans is a small organ that contains several neuroendocrine cells expressing various purinoceptors14,15,16,17,18. ATP and insulin are co-stored and co-secreted24,25. Insulin secretion is dependent on high blood glucose and ATP may regulate insulin secretion in a paracrine or autocrine manner24,25. However it is not still clear how ATP is stored in β-cells. As shown in Figure 3A, VNUT immunoreactivity was distributed throughout the islets of Langerhans. Double immunohistochemical labeling indicated that both α−cells and β-cells but not δ-cells expressed VNUT (Fig. 3B). Immunoelectron microscopy clearly indicated that VNUT immunoreactivity is associated with insulin granules of β-cells (Fig. 3C). Immunoreactivity to VNUT was also observed in exocrine acinar cells (Fig. 3A). In Vnut−/− mice, no VNUT immunoreactivity was observed in the islets of Langerhans or in pancreatic acinar cells (Fig. 3A). At the electron microscopic level, no apparent morphological changes in terms of number and diameter of secretory granules were observed in both α-cells and β-cells of islets from Vnut−/− cells (Fig. 3D).

VNUT acted as a negative regulator of insulin secretion in pancreatic islets.
(A) Indirect immunofluorescence microscopy revealed that VNUT was expressed in mouse pancreatic islets (WT). No VNUT immunoreactivity was seen in pancreatic islets isolated from Vnut−/− mice (KO). L, islet of Langerhans; a, acinar cell. Scale bar = 20 μm. (B) Localization of VNUT in islets. Sections of pancreas were double immunostained with antibodies against VNUT and insulin, VNUT and glucagon (marker of α−cells) and VNUT and somatostatin (marker of δ-cells) and observed by confocal laser microscopy. Merged pictures are also shown to the right. Areas surrounded by dotted line were enlarged in Supplementary Fig. S7. Bar = 20 μm. (C) Immunoelectron microscopy indicated that VNUT was localized at dense core granules of β-cells. Arrows indicate VNUT-positive signals (gold nanoparticles). The inset shows the control sample for which normal serum instead of mouse VNUT antibody was used. Scale bar = 500 nm. (D) Electron micrographs of ultrathin-sections of islets from WT and KO mice. The number and morphology of secretory granules in islet β-cells of KO mice were normal. Scale bar = 1 μm. (E) High glucose concentration-dependent release of ATP from pancreatic islets of WT (open bars, n = 10, 19) and KO mice (filled bars, n = 15, 14). (F) High glucose concentration-dependent secretion of insulin from pancreatic islets of WT (n = 8, 14, 8) and KO mice (n = 8, 14, 6). Where indicated, insulin secretion was measured in the presence of 0.1 mM ATP. (G) The amount of insulin in pancreatic islets of WT (n = 3) and KO mice (n = 4). (H) The ratio of proinsulin/insulin + proinsulin in pancreatic islets of WT (n = 16) and KO mice (n = 13). (I) The amount of ATP content in pancreatic islets of WT (n = 4) and KO mice (n = 4). Error bars represent mean ± S.E. *: p < 0.05. NS: not significant.
We next examined whether ATP is secreted from islets of Langerhans isolated from wild-type and Vnut−/− mice. Islets were incubated with Krebs-Ringer bicarbonate buffer containing 2.5 mM glucose and transferred into the same medium containing 2.5 mM glucose or 20 mM glucose, after which the concentrations of ATP and insulin in the medium and in the islets were determined. Appreciable amounts of ATP were released from wild-type islets of Langerhans upon stimulation by high concentrations of glucose, whereas no glucose-stimulated secretion of ATP was observed from those isolated from Vnut−/− mice (Fig. 3E). Under these conditions, insulin was also secreted upon stimulation with glucose from wild-type islets (Fig. 3F). In the islets of Vnut−/− mice, the glucose-stimulated insulin secretion was about 1.5-fold higher than that of the wild-type controls (Fig. 3F). The addition of low concentrations of ATP inhibited the glucose-stimulated release of insulin from both wild-type and Vnut−/− islets (Fig. 3F). No significant differences were observed in the insulin content and proinsulin/insulin ratio between islets from wild-type and Vnut−/− mice (Figs. 3G and H). The intracellular ATP affects insulin release through modification of ATP-sensitive K+ channel, but it was not altered in Vnut−/− (Fig. 3I). These results indicated that vesicular storage and subsequent exocytosis of ATP from β-cells were impaired in Vnut−/− mice and that vesicular ATP acts as a negative regulator of insulin secretion.
Vnut−/− mice were hypoglycemic
Decreased secretion of catecholamines from adrenal grand and increased secretion of insulin from islets in Vnut−/− mice suggested the possibility that the loss of ATP signaling affected glucose homeostasis. As expected, we found that upon fasting, the blood glucose level in Vnut−/− mice was significantly lower than that in controls (Fig. 4A). It is noted that both serum glucagon and adrenaline levels in Vnut−/− mice were comparable to those in control mice, although glucose levels were significantly lower in Vnut−/− mice. These results suggested that both glucagon and adrenaline secretion in response to hypoglycemia may be defective in Vnut−/− mice. Then we examined the effect of VNUT knockout on glucose tolerance. As shown in Figures 4B and C, Vnut−/− mice showed enhanced glucose tolerance after oral injection of glucose, although blood insulin levels were lower in Vnut−/− mice than controls. Under this condition, the serum GLP-1 level was not altered in Vnut−/− mice (data not shown). In addition, similar results were obtained by intreperitoneal injection of glucose (data not shown). These results indicated VNUT, known to be expressed in intestinal L cell, is not likely to involve in GLP-1 secretion26. The insulin tolerance test (ITT) showed that Vnut−/− mice had lower blood glucose levels than wild-type mice, indicating increased sensitivity to insulin (Fig. 4D). Under these conditions, the serum glucagon level in Vnut−/− mice was comparable to that in wild-type controls (Fig. 4E).

Vnut−/− mice were hypoglycemic.
(A) Serum glucose, glucagon and adrenalin levels after 24 h fasting in wild-type (WT, open bar, n = 3) and Vnut−/− mice (KO, filled bar, n = 4). (B) and (C) OGTT. Serum glucose (B) and insulin (C) levels were measured at indicated times after oral injection of glucose in WT (open squares, n = 7) and KO mice (filled squares, n = 10). (D) and (E) Serum glucose (D) and glucagon (E) levels during ITT in WT (n = 8) and KO mice (n = 17). Error bars represent mean ± S.E. *: p < 0.05.
The role of ATP signaling in glucose homeostasis in vivo
Finally, we investigated the mechanism for the altered glucose homeostasis in Vnut−/− mice. Since Akt functions as an intracellular regulator of insulin sensitivity27, we evaluated phosphorylation level of Akt (pAkt) in the liver. As shown in Fig. 5A, the relative amount of pAkt under basal conditions was significantly increased in Vnut−/− mice even though the total amount of Akt and pAkt was not different from controls. Addition of exogenous insulin further increased the relative proportion of pAkt in livers of both wild-type and Vnut−/− mice. A significant difference was also found in the amount of pAkt after insulin stimulation between Vnut−/− and wild-type mice. However, such a significant difference was not observed in skeletal muscle (Fig. S9). Glycogen contents in the livers of Vnut−/− mice were lower than that of wild-type mice under fed conditions, whereas there was no difference under fasting conditions (Fig. 5B). In wild-type mice, glycogen content was highest under fed conditions. In Vnut−/− mice, however, the differences in glycogen content between the fed and fasting conditions were not significant. Hepatic lipid content was increased after fasting in wild-type mice, whereas no increase was observed in Vnut−/− mice (Fig. 5C).

Glucose metabolism in Vnut−/− mice.
(A) Western blot analyses of liver homogenates (20 μg protein) from wild-type (WT, open bar, n = 3) and Vnut−/− (KO, filled bar, n = 3) mice using antibodies against Akt and pAkt (lower panel). The results of quantitative analysis are shown in the upper panel. The full-length blotting images are presented in Supplementary Fig. S8. (B) Glycogen contents in livers of WT (open bar, n = 3) and KO mice (filled bar, n = 4) before and after 6 h fast. (C) Total lipids in livers of WT (open bar, n = 6) and KO mice (filled bar, n = 6) before and after 6 h fast. Error bars represent mean ± S.E. *: p < 0.05. NS: not significant.
Discussion
In the present study we generated Vnut−/− mice and examined the changes in the properties of vesicular storage and release of ATP in neurons and two kinds of neuroendocrine cells. Previous studies have shown the wide physiological functions of purinergic chemical transmission, that VNUT is broadly expressed in the brain, including the cerebellar cortex, olfactory bulb and hippocampus and that it is responsible for vesicular storage and release of ATP23. Consistent with these studies, Vnut−/− mice lost vesicular ATP release from hippocampal neurons due to the lack of VNUT protein. It is noteworthy that VNUT is a member of SLC17 family and no isoform is presented in this family. Results show VNUT is an essential component of vesicular ATP release.
Although our behavioral analyses did not discriminate between wild-type and Vnut−/− mice, it doesn't imply less physiological relevance of VNUT in the central nervous system. Since purinergic signaling in brain is involved in cognition, learning, memory and sleep, more detailed behavioral analyses should be required to reveal physiological relevance of VNUT in brain28.
In contrast to the central nervous system, significant effects of VNUT gene knockout were observed in neuroendocrine cells. In adrenal chromaffin cells, ATP and catecholamine are co-stored in chromaffin granules through VNUT and VMAT-mediated active transport and are co-secreted through exocytosis. Various peptides such as encephalin, chromogranins and their degradation products are also secreted as intercellular messengers1,3,4. In Vnut−/− mice, it was found that vesicular ATP transport was absent, while reserpine-sensitive dopamine uptake was unchanged compared to controls. The secretion of catecholamines in the gland was not radically affected, although the content of catecholamine in the glands was decreased in Vnut−/− mice. The results are in line with the long-standing hypothesis that vesicular ATP is involved in the formation of an osmotically inactive complex with catecholamine and thereby plays an important role in synthesis and secretion of catecholamine1,3,29,30.
We confirmed that Vnut−/− mice lost vesicular storage and release of ATP from the chromaffin cells, resulting in impairment of purinergic chemical transmission. As expression of purinoceptors in the adrenal glands was not altered in Vnut−/− mice, stimulation of purinoceptors upon addition of exogenous ATP facilitated depolarization-dependent secretion of NA and Ad (Figs. 2H and I). Thus, it is concluded that vesicular ATP release in the adrenal gland is involved in the positive feedback regulation of catecholamine secretion through enhanced synthesis and exocytosis of catecholamines in vivo. Although contradictory secondary responses have been reported regarding catecholamine secretion from chromaffin cells7,8,9,10,11,12,13, the action by the P2Y2 and P2X4-7 receptors may dominate in adrenal chromaffin cells in vivo.
In the islets of Langerhans, we found that VNUT was associated with insulin granules of β-cells and glucose-stimulated ATP secretion from β-cells was not detected in Vnut−/− mice. These results indicated that insulin and ATP are co-stored and co-secreted upon glucose stimulation through a VNUT-dependent mechanism. Recently, Geisler et al. reported that suppression of VNUT gene expression in isolated islets and MIN6 clonal cells by RNA interference decreased secretion of both ATP and insulin; they concluded that ATP signaling enhances the secretion of insulin through P2X receptor signaling31. On the other hand, in the islets of Vnut−/− mice, glucose dependent insulin release was stimulated. Contradictory results may partly due to the expression level of VNUT protein and cells used in the experiments. Although shRNA treatment decreased VNUT mRNA level, it retained significant level of VNUT protein. In addition, they used MIN6 cells as a model of β cell. Alternatively, there may be some modification in Vnut−/− islet during development. Such methodological differences may cause the discrepancy between these studies.
In the islets from Vnut−/− mice, the content of insulin and insulin/proinsulin ratio were comparable to those of control mice, indicating that vesicular ATP is not involved in the biosynthesis and processing of insulin. As shown previously, β-cells secrete ATP with insulin when glucose concentration increases and the released ATP leads to complex effects on insulin secretion that are dependent on species and receptor type24,25. For example, ATP and its derivatives enhance insulin secretion in different species including rodents and humans14,15,16,17,18. It was also reported that ATP inhibits insulin secretion through the P2Y1 receptor and/or A1 receptor in mice14,32. It should be stressed that in rats and dogs, ATP or ATP analogues, such as 2-methylthio ATP and ADPβS, cause opposite effects to that in mice and facilitating insulin secretion and improving glucose tolerance33. A recent study using specific ligands showed that stimulation of P2Y13 receptor inhibits insulin secretion, whereas stimulation of P2Y1 receptor increases insulin secretion34. The P2Y13 antagonist, MRS2211, reduced plasma glucose concentrations by increased plasma insulin level34. Taken together, these observations indicate that the multiple effects of nucleotides on insulin secretion are dependent on species and receptor type. In this study, we have provided evidence that glucose-dependent ATP release is impaired and that glucose-dependent secretion of insulin is enhanced in Vnut−/− mice. Thus, ATP release from β-cells is involved in the negative feedback regulation of insulin release in vivo and loss or decreased purinergic chemical transmission blocks the feedback loop, resulting in increased insulin secretion in mice. The inhibitory effect of P2Y13 is probably dominant to the stimulatory effect of P2Y1 on insulin secretion in mice.
This conclusion was collaborated by our observations that Vnut−/− mice were hypoglycemic after 24 h fasting and showed improved glucose tolerance after glucose challenge. These results provide compelling evidence that vesicular ATP actually participates in the purinergic chemical transmission in chromaffin cells and islets and that its regulation is involved in glucose homeostasis in vivo. Under fasting, serum glucose concentration is controlled by increased glucagon and adrenaline secretion. Interestingly, both glucagon and adrenaline levels were not altered in Vnut−/− mice suggesting a defective glucagon and adrenalin secretion in response to hypoglycemia in Vnut−/− mice.
Vnut−/− mice also exhibited hypoglycemia under glucose tolerance test indicating increased insulin sensitivity. It should be stressed that we also observed elevated phosphorylation of hepatic Akt, a key protein in the insulin signaling cascade that is downstream of the insulin receptor and insulin receptor substrate (IRS)27, in Vnut−/− mice. Activation of the insulin receptor promotes phosphorylation of Akt and leads to metabolic responses, such as activation of glucose uptake and glycogen and fatty acid synthesis. Thus, increased phosphorylation of Akt in Vnut−/− mice may explain the improved glucose tolerance and insulin sensitivity. It is interest to note that Akt phosphorylation in hepatocytes is inhibited by binding of ADP to the P2Y13 receptor, suggesting that loss of purinergic signaling stimulates Akt phosphorylation, in good agreement with our observations in Vnut−/− mice35. Interestingly, gene disruption of Cd39/Entpd1, an ecto-ATPase, resulted in hyperglycemia and impaired glucose tolerance, a phenotype that was almost the opposite of that in Vnut−/− mice36. Cd39/Entpd1−/− mice also showed decreased glucose uptake and inhibited IRS. These results support our hypothesis that purinergic signaling modulates glucose metabolism through inhibition of the insulin signaling cascade. In the wild-type liver, a reduction of glycogen content and an increase of lipid content were observed after fasting. However, these changes in glycogen and lipid metabolism were not observed in Vnut−/− mice, suggesting roles of purinergic signaling in glucose and lipid metabolism in the liver. On the other hand, a statistical difference in the Akt phosphorylation was not found in skeletal muscle, the other major insulin target organ. This may be due to the lower expression level in muscle compared to the liver. Alternatively, the experimental condition using high dose insulin may not be a good system to detect the effect of VNUT on insulin signaling in the muscle. Recently, ATP released from skeletal muscle by exercise has shown to increase glucose uptake in rat muscle fibers37, a further study on the VNUT role in muscle will be guaranteed. Nonetheless, impairment of vesicular ATP release induces hypoglycemia, which may correlate with the upregulation of Akt-dependent energy metabolism and suggests that VNUT is a potential and novel target of antidiabetic agents.
In conclusion, we demonstrated that VNUT-mediated ATP transport is essential to vesicular storage and subsequent release of ATP in neuroendocrine cells. Vnut−/− mice lose purinergic signal output due to defective ATP vesicular storage and release. As a consequence, a profound relationship between vesicular ATP and glucose metabolism was revealed. Vnut−/− mice are a potential tool to investigate the overall mechanism and physiological relevance of ATP release from purinergic cells.
Methods
Generation of Vnut−/− mice
Mouse experiments were performed according to guidelines set by the animal ethics committees of Kyushu University, Okayama University and Ajinomoto Co. Ltd. All experiments were carried out in accordance with the approved institutional guidelines.
Vnut−/− mice were generated by gene disruption (Fig. 1). Briefly, 4.0 kb and 4.7 kb DNA fragments that flank exons 2 and 3 and which were used as the 5′ and 3′ arms, respectively and were amplified by PCR from 129/Sv mouse genomic DNA and subcloned into a pPGKneobpAlox2PGKDTA vector containing the neomycin resistance gene (Neo) as a positive selection marker and the diphtheria toxin A fragment gene (DT-A) as a negative selection marker. The targeting vector was linearized with Xho I and transfected into mouse embryonic stem (ES) cells. Targeted Vnut+/− ES clones were injected into C57BL/6J blastocysts to obtain chimeric mice. Male chimeric mice were crossed with female C57BL/6J to generate Vnut+/− mice. The heterozygous offspring were intercrossed to generate homozygous knockout (KO, Vnut−/−) mice. Homozygous Vnut−/− mice were obtained at the expected Mendelian frequency. Genomic DNA was isolated from tail biopsies and was used as the template for genotyping PCR. The primers used for the wild-type allele were: WT forward, 5′-CTATGTGTAGCCCTGGATGG-3′ and WT reverse, 5′-GTGTACCCTTCGGGGAAAGT-3′. The primers for the knockout allele were: KO forward, 5′-CTTGTGTTTGGGATCCTGGT-3′ and KO reverse, 5′-GGGAGGATTGGG AAGACAAT-3′. These primers amplified a 217 bp fragment from the endogenous Vnut wild-type allele and a fragment of 314 bp from the Vnut targeted allele.
Uptake of ATP and dopamine by chromaffin granule membrane vesicles
Membrane vesicles of chromaffin granules were prepared from adrenal glands of 30 wild-type and 30 Vnut−/− mice each as described previously38. The membrane vesicles were suspended in reaction mixture consisting of 20 mM MOPS-Tris (pH 7.0), 0.3 M sucrose, 2 mM magnesium acetate, 4 mM KCl and incubated for 2 min at 30°C. The assay was initiated by the addition of 2 mM [2,8-3H] ATP (20 MBq/μmol) and 100-μl aliquots were taken and filtered through nitrocellulose filters (Millipore, USA). After washing with 6 ml of cold reaction buffer, the radioactivity remaining on the filters was counted.
For dopamine uptake, the membrane vesicles were suspended in reaction mixture consisting of 20 mM MOPS-Tris (pH 7.0), 0.1 M KCl, 0.1 M sucrose and 5 mM magnesium acetate in the presence or absence of 1 μM reserpine and incubated for 2 min at 30°C. The assay was initiated by the addition of 5 mM ATP and 10 μM [ring-2, 5, 6-3H] dopamine (0.5 MBq/μmol).
ATP, noradrenaline and adrenaline release from isolated chromaffin cells
Adrenal chromaffin cells were isolated from wild-type and Vnut−/− mice as described previously39. Primary culture cells were washed three times with Krebs-Ringer bicarbonate buffer composed of 128 mM NaCl, 1.9 mM KCl, 1.2 mM KH2PO4 1.3 mM MgSO4, 26 mM NaHCO3, 10 mM D-glucose, 10 mM HEPES-Tris, pH 7.4, 2.4 mM CaCl2 and 0.2% (w/v) bovine serum albumin (BSA). After the cells had been incubated in Krebs-Ringer bicarbonate buffer at 37°C for 20 min, ATP, noradrenaline (NA) and adrenaline (Ad) release were stimulated by the addition of 50 mM KCl. After incubation at 37°C for 20 min, aliquots were taken and the amount of ATP was measured using the Kinshiro ATP luminescence kit (TOYO INK, Japan) and the amounts of NA and Ad were measured by 3-CAT Research ELISA™ (Labor Diagnostika Nord GmbH & Co. KG, Nordhorn, Germany).
To determine the NA and Ad contents of whole adrenal glands, four glands per assay were homogenized in buffer consisting of 10 mM MOPS-Tris (pH 7.0), 0.3 M sucrose and 5 mM EDTA and centrifuged at 1,300 × g, for 8 min. NA and Ad contents in the supernatant were measured. To measure plasma catecholamine levels, blood samples (100 μl) were collected from the tail vein of mice fasted for 18 h or randomly fed. Samples were centrifuged and the plasma used to determine the catecholamine concentration.
ATP and insulin secretion from isolated islets
Pancreatic islets were isolated from Vnut−/− and wild-type mice as described previously40. Each group of five size-matched islets (100–200 μm in diameter) was preincubated for 1 h at 37°C in Krebs-Ringer bicarbonate buffer composed of 135 mM NaCl, 3.6 mM KCl, 1.5 mM CaCl2, 0.5 mM MgCl2, 2 mM NaHCO3, 0.5 mM NaH2PO4 and 10 mM HEPES containing 2.5 mM D-glucose. After preincubation, the buffer composition was changed to 20 mM glucose or 2.5 mM glucose plus 0.2% (w/v) BSA. After 60 min, the medium from each group was collected. Insulin concentrations in the medium were measured using the Mouse Insulin ELISA Kit (Morinaga Institute of Biological Science, Japan). ATP content in the medium was measured as described above.
To determine their insulin content, the islets were sonicated in 0.2 mM hydrochloric acid and 75% ethanol on ice and incubated overnight at 4°C. After a brief centrifugation (800 × g, 15 min, 4°C), the supernatant was used for the measurement of insulin content using the insulin ELISA kit named above.
Antibodies
Primary antibodies for immunoblotting were purchased from the following commercial sources: anti-VMAT1 rabbit polyclonal antibody (Abcam, UK), anti-VMAT2 rabbit polyclonal antibody (Abcam, UK), anti-insulin guinea pig polyclonal antibody (Abcam, UK), anti-glucagon mouse monoclonal antibody (Sigma, USA), anti-somatostatin rat monoclonal antibody (Chemicon, USA) and anti-β actin mouse monoclonal antibody (Abcam, UK). Antibodies against mouse VNUT, VGLUT1, VGLUT2, VEAT, VGAT, SV2, VAMP2, NSF and subunit A of V-ATPase for immunoblotting and immunohistochemistry were prepared as described previously22,23,41,42,43,44,45.
Immunohistochemistry
Indirect immunofluorescence microscopy was performed as described41. Mouse VNUT antibody was diluted 1:200 (v/v) with PBS containing 0.5% (w/v) BSA and used for immunostaining. Anti-insulin guinea pig polyclonal antibody (1:2000), anti-glucagon mouse monoclonal antibody (1:500000) and anti-somatostatin rat monoclonal antibody (1:2000) were used for immunostaining of the respective antigens.
Electron microscopy
Conventional and immunoelectron microcopies were carried out as described previously41,46. Adult Vnut−/− and wild-type male mice were anesthetized with ether and then perfused intracardially with saline, followed by 2.5% or 4% (w/v) glutaraldehyde in 0.1 M phosphate buffer (pH 7.4). Ultrathin sections of adrenal glands and pancreas were observed under a Hitachi H-7600 electron microscope.
Real-time PCR
Total RNA from the adrenal glands of wild-type and Vnut−/− mice was extracted with Isogen II (Nippon Gene, Toyama, Japan). cDNA was generated from 1 μg of total RNA with High Capacity RNA-to-cDNA Kit (Applied Biosystems, Foster City, CA). The resulting cDNA pool was used for real-time PCR, which was carried out with TaqMan® Gene Expression Master Mix and the respective probe sets (Applied Biosystems). Forty cycles were performed with denaturation at 95°C for 15 s and annealing/extension at 60°C for 30 s. The comparative CT method was used for relative quantification.
Oxygen consumption measurements
Mice were subjected to oxygen consumption measurements using a computer-controlled open-circuit indirect calorimeter (Oxymax; Columbus Instruments, USA) and the data were analyzed using Oxymax Windows software47.
Behavior-based test
Each animal was set on the open field (60 cm × 60 cm) or the plus maze apparatus (30 cm × 5 cm closed arm, with a 25 cm high wall, 47 cm above the floor) and its behavior was recorded on video for 10 min48.
Oral glucose tolerance test and insulin tolerance test
For the oral glucose tolerance test (OGTT), mice fasted for 18 h were injected orally with 2 g of glucose/kg body weight dissolved in saline. To measure plasma insulin levels, blood samples (100 μl) were collected from the tail vein and centrifuged. For insulin tolerance tests (ITT), mice fasted for 4 h were injected intraperitoneally with 0.75 units of human regular insulin/kg body weight. In each test, blood glucose levels were measured using a Glutest Sensor (Sanwa Kagaku Inc., Tokyo, Japan).
Insulin signaling
After an overnight fast, mice were anesthetized and the abdominal cavity was opened to inject either 5 units regular human insulin or insulin diluent, diluted in 0.9% saline (final volume 125 μl) into the inferior vena cava. Samples of liver were collected 2 min after injection and analyzed by Western blotting49.
Glycogen and lipid measurement
To measure glycogen levels, livers were isolated from mice fed and then fasted for 6 h. The tissues were homogenized in distilled water and boiled for 10 min. After centrifugation at 16400 × g for 10 min, the supernatant was measured using glycogen assay kit (BioAssay Systems, Hayward, USA). The total lipid levels were measured after Folch lipid extraction.
Statistical analysis
All numerical values are shown as the mean ± S.E. for 3 to 17 experiments. Statistical significance was determined by Student's t test.
References
Winkler, H. & Carmichael, S. W. in The Secretory granule, Poisner, A. M. & Trifaro, J. M. Eds. (Elsevier Biomedical, Amsterdam, 1982), pp 3–79.
Johnson, R. G., Carty, S. E. & Scarpa, A. Catecholamine transport and energy-linked function of chromaffin granules isolated from a human pheochromocytoma. Biochim. Biophys. Acta 716, 366–176 (1982).
Johnson, R. G. Accumulation of biological amines into chromaffin granules: a model for hormone and neurotransmitter transport. Physiol. Rev. 68, 232–307 (1988).
Laslop, A. & Mahata, S. K. Neuropeptide and chromogranins: session overview. Ann. NY Acad. Sci. 971, 294–299 (2002).
Schuldiner, S., Shirvan, A. & Linia, M. Vesicular neurotransmitter transporters: From bacteria to humans. Physiol. Rev. 75, 369–392 (1955).
Burnstock, G. Physiology and pathophysiology of purinergic neurotransmission. Physiol. Rev. 87, 659–797 (2007).
Currie, K. P. & Fox, A. P. ATP serves as a negative feedback inhibitor of voltage-gated Ca2+ channel currents in cultured bovine adrenal chromaffin cells. Neuron 16, 1027–1036 (1996).
Harkins, A. B. & Fox, A. P. Activation of purinergic receptors by ATP inhibits secretion in bovine adrenal chromaffin cells. Brain Res. 885, 231–239 (2000).
Ennion, S. J., Powell, A. D. & Seward, E. P. Identification of the P2Y12 receptor in nucleotide inhibition of exocytosis from bovine chromaffin cells. Mol. Pharmacol. 66, 601–611 (2004).
Reichsman, F., Santos, S. & Westhead, E. W. Two distinct ATP receptors activate calcium entry and internal calcium release in bovine chromaffin cells. J. Neurochem. 65, 2080–2086 (1995).
Tome, A. R., Castro, E., Santos, R. M. & Rosario, L. M. Functional distribution of Ca2+-coupled P2 purinergic receptors among adrenergic and noradrenergic bovine adrenal chromaffin cells. BMC Neurosci. 8, 39 (2007).
Afework, M. & Burnstock, G. Changes in P2Y2 receptor localization on adrenaline- and noradrenaline containing chromaffin cells in the rat adrenal gland during development and ageing. Int. J. Dev. Neurosci. 23, 567–573 (2005).
Afework, M. & Burnstock, G. Distribution of P2X receptors in the rat adrenal gland. Cell Tissue Res. 298, 449–456 (1999).
Petit, P., Lajoix, A. D. & Gross, R. P2 purinergic signalling in the pancreatic beta-cell: control of insulin secretion and pharmacology. Eur. J. Pharm. Sci. 37, 67–75 (2009).
Novak, I. Purinergic receptors in the endocrine and exocrine pancreas. Purinergic Signaling 4, 237–253 (2008).
Jacques-Silva, M. C. et al. ATP-gated P2X3 receptors constitute a positive autocrine signal for insulin release in the human pancreatic beta cell. Proc. Natl. Acad. Sci. USA 107, 6465–6470 (2010).
Silva, A. M. et al. Electrophysiological and immunocytochemical evidence for P2X purinergic receptors in pancreatic beta cells. Pancreas 36, 279–283 (2008).
Leon, C. et al. The P2Y1 receptor is involved in the maintenance of glucose homeostasis and in insulin secretion in mice. Purinergic Signaling 1, 145–151 (2005).
Gachet, C. P2 receptors, platelet function and pharmacological implications. Thromb. Haemost. 99, 466–472 (2008).
Fabre, J. E. et al. Decreased platelet aggregation, increased bleeding time and resistance to thromboembolism in P2Y1-deficient mice. Nat. Med. 5, 1199–1202 (1999).
Kim, S. & Kunapuli, S. P. P2Y12 receptor in platelet activation. Platelets 22, 54–58 (2011).
Sawada, K. et al. Identification of a vesicular nucleotide transporter. Proc. Natl. Acad. Sci. USA 105, 5683–5686 (2008).
Larsson, M. et al. Functional and anatomical identification of a vesicular transporter mediating neuronal ATP release. Cereb. Cortex 22, 1203–1214 (2012).
Suckale, J. & Solimena, M. The insulin secretory granule as a signaling hub. Trend Endocrinol. Metab. 21, 599–609 (2010).
Eliasson, L. et al. Novel aspects of the molecular mechanisms controlling insulin secretion. J. Physiol. 586, 3313–3324 (2008).
Harada, Y. & Hiasa, M. Immunological identification of vesicular nucleotide transporter in intestinal L cells. Biol. Pharm. Bull. 37, 1090–1095 (2014).
Samuel, V. T. & Shulman, G. I. Mechanism for insulin resistance: common threads and missing links. Cell 148, 852–871 (2012).
Burnstock, G., Krügel, U., Abbracchio, M. P. & Illes, P. Purinergic signalling: from normal behaviour to pathological brain function. Prog. Neurobiol. 95, 229–274 (2011).
Kopell, W. N. & Westhead, E. W. Osmotic pressures of solutions of ATP and catecholamines relating to storage in chromaffin granules. J. Biol. Chem. 257, 5707–5710 (1982).
Phillips, J. H. & Morton, A. G. Adenosine triphosphate in the bovine chromaffin granule. J. Physiol. 74, 503–508 (1978).
Geisler, J. C. et al. Vesicular nucleotide transporter-mediated ATP release regulates insulin secretion. Endocrinology 154, 675–684 (2013).
Johansson, S. M. et al. A1 receptor deficiency causes increased insulin and glucagon secretion in mice. Biochem. Pharmacol. 74, 1628–1635 (2007).
Hillaire-Buys, D. et al. Stimulation of insulin secretion and improvement of glucose tolerance in rat and dog by the P2Y-purinoreceptor agonist, adenosine-5′-O-(2-thiodiphosphate). Br. J. Pharmacol. 109, 183–187 (1993).
Amisten, S. et al. ADP mediates inhibition of insulin secretion by activation of P2Y13 receptors in mice. Diabetologia 53, 1927–1934 (2010).
Chatterjee, C. & Sparks, D. L. Extracellular nucleotides inhibit insulin receptor signaling, stimulate autophagy and control lipoprotein. PLoS ONE 7, e36916 (2012).
Enjyoji, K. et al. Deletion of Cd39/Entpd1 results in hepatic insulin resistance. Diabetes 57, 2311–2320 (2008).
Osorio-Fuentealba, C. et al. Electrical stimuli release ATP to increase glut4 translocation and glucose uptake via PI3Kγ-Akt-AS160 in skeletal muscle cells. Diabetes 62, 1519–1526 (2013).
Bankston, L. A. & Guidotti, G. Characterization of ATP transport into chromaffin granule ghosts: Synergy of ATP and serotonin accumulation in chromaffin granule ghosts. J. Biol. Chem. 271, 17132–17138 (1996).
Gilabert, J. A., Montalvo, G. B. & Artalejo, A. R. Rat chromaffin cells primary cultures: Standardization and quality assessment for single-cell assays. Nat. Protoc. 2006; 10.1038/nprot.2006.294.
Lacy, P. E. & Kostianovsky, M. Method for the isolation of intact islets of Langerhans from the rat pancreas. Diabetes 16, 35–39 (1967).
Hayashi, M. et al. Secretory granule-mediated cosecretion of L-glutamate and glucagon triggers glutamatergic signal transmission in islets of Langerhans. J. Biol. Chem. 278, 1966–1974 (2003).
Miyaji, T. et al. Identification of a vesicular aspartate transporter. Proc. Natl. Acad. Sci. USA 105, 11720–11724 (2008).
Juge, N., Muroyama, A., Hiasa, M., Omote, H. & Moriyama, Y. Vesicular inhibitory amino acid transporter is a Cl−/γ-aminobutyrate co-transporter. J. Biol. Chem. 284, 35073–35078 (2009).
Hayashi, M. et al. Synaptic vesicle protein SV2B, but not SV2A, is predominantly expressed and associated with microvesicles in rat pinealocytes. J. Neurochem. 71, 356–365 (1988).
Moriyama, Y., Yamamoto, A., Tagaya, M., Tashiro, Y. & Michibata, H. Localization of N-ethylmaleimide-sensitive fusion protein in pinealocytes. Neuroreport 6, 1757–1760 (1995).
Yamamoto, A. & Masaki, R. Pre-embedding nanogold silver and gold intensification. Methods Mol. Biol. 657, 225–235 (2010).
Fan, W. et al. Androgen receptor null male mice develop late-onset obesity caused by decreased energy expenditure and lipolytic activity but show normal insulin sensitivity with high adiponectin secretion. Diabetes 54, 1000–1008 (2005).
Ansorge, M. S., Zhou, M., Lira, A., Hen, J. & Gingrich, J. A. Early-life blockade of the 5-HT transporter alters emotional behavior in adult mice. Science 306, 879–881 (2004).
Ueki, K. et al. Increased insulin sensitivity in mice lacking p85β subunit of phosphoinositide 3-kinase. Proc. Natl. Acad. Sci. USA 99, 419–424 (2002).
Acknowledgements
This work was supported in part by the following awards or organizations: Grants-in-Aid for Scientific Research (A) (No. 2523008), the Salt Science Foundation (No. 1139) and the Japan Science and Technology Agency for Japan-Israel Scientific Research cooperation to Y.M.; Grants-in-Aid for Young Scientists (B) (No. 40550314), the Smoking Research Foundation, the Uehara Memorial Foundation and the Takada Science Foundation to T.M.; Grants-in-Aid for Research Fellowship for Young Scientists and the Program to Disseminate Tenure Tracking System to M.H.; Grants-in-Aid for Scientific Research (B) (No. 21370057) to H.O.; and Grants-in-Aid for Scientific Research (C) to M.N. The authors declare that they have no conflicting financial interests.
Author information
Authors and Affiliations
Contributions
S.S. and T.M. contributed equally to this work. S.S., T.M., M.H., K.I., H.O., M.N. and Y.M. designed the experiments, analyzed the data and wrote the paper. S.S., T.M., M.H., R.I., A.U., K.I., A.S. and A.Y. performed the experiments. H.U. and R.T. analyzed the data. M.N. and Y.M. are the guarantors of the article.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Supplemental Figs
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/4.0/
About this article

Cite this article
Sakamoto, S., Miyaji, T., Hiasa, M. et al. Impairment of vesicular ATP release affects glucose metabolism and increases insulin sensitivity. Sci Rep 4, 6689 (2014). https://doi.org/10.1038/srep06689
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep06689
This article is cited by
-
Hepatic extracellular ATP/adenosine dynamics in zebrafish models of alcoholic and metabolic steatotic liver disease
Scientific Reports (2024)
-
Clodronate, an inhibitor of the vesicular nucleotide transporter, ameliorates steatohepatitis and acute liver injury
Scientific Reports (2021)
-
Quinacrine is not a vital fluorescent probe for vesicular ATP storage
Purinergic Signalling (2021)
-
Adenosine triphosphate is co-secreted with glucagon-like peptide-1 to modulate intestinal enterocytes and afferent neurons
Nature Communications (2019)
-
The P2X7 receptor and pannexin-1 are involved in glucose-induced autocrine regulation in β-cells
Scientific Reports (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
