
Abstract
The Aptian Jiufotang Formation of northeast China is a Konservat Lagerstätte particularly rich in pterosaurs, notably azhdarchoids. Here we describe a new genus and species of toothed pteranodontoid pterosaur, Ikrandraco avatar gen. et sp. nov., based on two laterally flattened specimens. Ikrandraco avatar is diagnosed by a suite of features, including a very low and elongate skull, strongly inclined quadrate and a deep, blade-like bony mandibular crest with a hook-like process on its posterior edge, an unusual structure so far unique to this taxon. The particular skull shape hints at a distinct feeding habit for pterosaurs that potentially includes temporary skimming and an extensible skin acting as a throat pouch that was more developed than in any other pterosaur known so far. The presence of two other taxa of purported piscivorous pterosaurs in the Jiufotang Formation suggests distinct resource exploitation in this part of China during the Early Cretaceous.
Similar content being viewed by others
Introduction
Pterosaurs represent an important group of Mesozoic reptiles, being the first vertebrates to occupy the aerial niche1. Despite being represented in all continents, their fossil record is very uneven2. Only a few deposits bear complete and well-preserved specimens, thus contributing to the current knowledge on pterosaur anatomy, diversity and evolution3,4. Among those is the Jehol Group of northeastern China, with pterosaurs found in the Jiufotang and Yixian formations playing a crucial role in providing novel information about the evolutionary history of those flying reptiles. In the last decade, more specimens have been recovered from these lacustrine deposits than from any other region in the world, revealing a large diversity of species. Regarding pterodactyloids, the most productive deposit of those two has been the Lower Cretaceous Jiufotang Formation5, with some specimens showing quite distinct morphologies. So far, only dsungaripteroid pterodactyloids6 have been recovered, some closely related to the taxa known from the renowned Early Cretaceous (Aptian-Albian) Romualdo and Crato formations of northeastern Brazil7. Among those are tapejarines8,9 and anhanguerians10,11, thus suggesting a very interesting biogeographical tie between those pterosaur faunas11,12. The Jiufotang Formation, however, has a higher diversity of more inclusive clades, as exemplified by the presence of istiodactylids, chaoyangopterines and one basal dsungaripteroid3,13,14,15, representing therefore a unique pterosaur fauna composition.
Here, we describe a new pterosaur species from this unit with very distinct cranial and mandibular morphology, uniting a low and elongate crestless skull with a lower jaw that has a developed dentary crest with a hook-shaped posterior process. Such combination of morphological features has no parallel among the known pterosaur species and suggests a specialized foraging habit of this new flying reptile.
Results
Systematic paleontology
Pterosauria Kaup, 1834
Pterodactyloidea Plieninger, 1901
Dsungaripteroidea Young, 1964
Pteranodontoidea Marsh, 1876
Ikrandraco gen. nov.
Type species
Ikrandraco avatar, type by monotypy.
Etymology
Ikran, from the fictional flying creature portrayed in the movie Avatar that shows a well developed dentary crest and draco, from the Latin meaning dragon.
Diagnosis
The same for the type species.
Ikrandraco avatar sp. nov.
Etymology
Avatar, in allusion to the homonymous science fiction movie.
Holotype
A partial skeleton including complete skull and mandible, atlas, axis, three mid-cervical vertebrae, part of the sternal plate, some ribs, partial right and left wings and part of the foot (IVPP V18199), deposited at the Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences in Beijing, China.
Referred specimen
A complete skull, mandible, atlas, axis and third cervical vertebra (IVPP V18406), deposited at the Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences in Beijing, China.
Horizon and locality
Jiufotang Formation, Early Cretaceous (Aptian) 120 Ma5. IVPP V18199 was collected in Lamadong, Jianchang and IVPP V18406 comes from Sihedang, Lingyuan, western Liaoning, China.
Diagnosis
Ikrandraco avatar can be distinguished from all other pteranodontoid pterodactyloids by the following autapomorphies: slightly arched dorsal margin of the skull above the nasoantorbital fenestrae; lateral depression on the nasal; median hook-like process on the posterior edge of the dentary crest; two well-developed pneumatic foramina piercing the lateral surface of the axis. It can further be separated from other pteranodontoids by the following combination of characters: very low skull (height over quadrates about 18.7% of skull length); strongly inclined quadrate (150°); lack of a premaxillary crest; presence of a deep, blade-like bony mandibular crest with deepest point at mid-length; and a ventral pneumatic foramen on the proximal portion of the second phalanx and third phalanges of the wing finger.
Description
Both pterosaur specimens are laterally compressed, a common condition among vertebrates from the Jiufotang Formation. The holotype (Figs. 1, 2a,b, 3) shows that the tip of the cranium and the first and second right wing finger phalanges slightly displaced from the rest of the skeleton by a fracture. A piece of fossil wood is associated with the skeleton. Based on the fusion of the extensor tendon process this specimen can be considered an adult individual. In the referred specimen (Figs. 2c,d) some of the cortical bone, particularly of the vertebrae, is missing and the bones are not as well preserved as in the holotype.

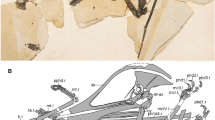
Ikrandraco avatar, holotype (IVPP V18199), nearly complete skeleton.
(a) Photograph; (b) respective line drawing. Abbreviations: art, articular; at, atlas; ax, axis; cv, cervical vertebra; d, dentary; dca, distal carpal; dcr, dentary crest; dlca, distal lateral carpal; fopn, pneumatic foramen; hu, humerus; m, maxilla; mcI-IV, metacarpal I-IV; mt I-IV, metatarsal I-IV; naof, nasoantorbital fenestra; or, orbit; ph1-3d4, first to third phalanx of manual digit IV; pla, plant; pm, premaxilla; prca, proximal carpal; ptd, pteroid; ra, radius; ri, rib; sri, sacral rib; st, sternum; te, teeth; ul, ulna; un, ungual; l, left; r, right. Scale bar: 50 mm. Line drawing by S.J.

Ikrandraco avatar, skull and mandible.
(a–b) Photograph and line drawing of the holotype (IVPP V18199), (c–d) Photograph and line drawing of the referred specimen (IVPP V18046). Abbreviations: art, articular; ax, axis; bo, basioccipital; bpp, basisphenoid process; bs, basisphenoid; ch, choanae; cv, cervical vertebra; d, dentary; dcr, dentary crest; dep, depression; f, frontal; fon, foramen nasalis; fola, foramen lacrimale; foprn, foramen of nasal process; hy, hyoid; ios, interorbital septum; ipf, interpterygoid fenestra; j, jugal; la, lacrimal; ltf, lower temporal fenestra; m, maxilla; n, nasal; naof, nasoantorbital fenestra; oc, occipital condyle; op, opisthotic; or, orbit; p, parietal; pcf, posterior cranial fenestra; pl, palatine; pm, premaxilla; po, postorbital; pplf, postpalatinal fenestra; prid, palatal ridge; prn, processus nasalis; psm, pseudomesethmoid; ptf, posttemporal fenestra; pty, pterygoid; ptyf, pterygoid fenestra; q, quadrate; qj, quadratojugal; scl, sclerotic ring; soc, supraoccipital; socr, supraoccipital crest; sq, squamosal; stf, subtemporal fenestra; te, teeth; ul, ulna; utf, upper temporal fenestra; vo, vomer; l, left; r, right. Scale bars: 50 mm. Line drawing by S.J.

Ikrandraco avatar, holotype IVPP V18199.
(a) Anterior tip of the skull; arrows point the palatal ridge. (b) Left maxilla; arrows point a small pathology. (c) Nasal process and nasal. (d) Cervical vertebrae; arrows point the two foramina of axis. (e) Humerus. (f) Occipital region; the arrow points the basioccipital. Scale bars: 10 mm.
The skull is exposed in left lateral view in both specimens. It is very elongate and low, with a long postorbital region compared to other pteranodontoids. The orbit is oval, more rounded dorsally than ventrally (Supplementary Fig S1). The nasoantorbital fenestra occupies about one third of the skull length, with an elongate triangular profile and the anterior margin slightly more pointed in the holotype, possibly due to taphonomic compression. The anterior-most point of the nasoantorbital fenestra lies closer to the dorsal margin of the skull than to its ventral margin, as seen in Tropeognathus cf. T. mesembrinus7. The lower temporal fenestra has a tear-drop shape, with the dorsal part broader than the ventral.
As usual in pterodactyloids, the premaxillae and maxillae are fused. The premaxilla occupies the anterodorsal part of the skull, forming the dorsal margin of the nasoantorbital fenestra and shows no premaxillary sagittal crest. In the holotype, there is a sign of pathology in the left maxilla, where the bone is more rugose (Fig. 3b). The maxillary ventral margin shows some rounded depressions between the alveoli, which might have accommodated the lower teeth during occlusion. Although such features have occasionally been observed in other pterosaurs, in Ikrandraco avatar they are more numerous.
The jugal process of the maxilla ends between the postpalatine and the subtemporal fenestrae (Fig. 2). A thin palatal ridge extends between the 5th and 11th tooth sockets with maximum depth between the 9th and 10th (in the holotype; Figs. 2a,b). The nasal is elongate, pierced by a foramen and has a marked depression. The nasal process is very thin and long, located medially, inclined anteriorly and bears a small foramen on its dorsal portion, differing from the large foramen observed in some anhanguerids16,17. A well-defined suture separates the nasal and the nasal process in the holotype (Fig. 3c), suggesting that this process is a distinct ossification and potentially not homologous with the extension of the nasal that forms the maxillae-nasal bar separating the external naris and the antorbital fenestra of non-pterodactyloid pterosaurs. Both frontals and parietals bear no crests. The lacrimal has a small foramen that is positioned more dorsally than in anhanguerids16,17,18 and is also potentially not homologous with the foramen described in other pterosaurs. The jugal has a broad and slightly anteriorly inclined lacrimal process and, differing from other pteranodontoids, shows no lateral ridge. The quadrate has a helical jaw joint and a reduced medial flange, smaller than in other Dsungaripteroidea. It is further inclined for about 150°, similar to archaeopterodactyloids but differing from other pteranodontoids6.
Due to taphonomic crushing, part of the palatal region became visible on both specimens, but elements are better observed in the holotype. As usual in pterodactyloids, the palatines are firmly fused to the premaxillae and maxillae and form the anterior margin of the choanae. The choanae occupy approximately 29% of the skull length (82.3 mm) and are divided by a thin process of the vomers. Their anterior margin is coincident with that of the nasoantorbital fenestra. The interpterygoid fenestra is well-developed, unlike the reduced condition seen in Pteranodon19. The pterygoid fenestra shows an oval shape. The postpalatinal fenestra (7.5 mm) is smaller than the subtemporal fenestra (17.3 mm). The pterygoid is elongate and forms the posterior and most of the lateral margins of the choanae.
The occipital region is better observed in the holotype (Fig. 3f). The supraoccipital crest is well-developed, much larger than in the Anhangueridae16,17,18,19. The basioccipital has a small foramen on its lateral flange. The occipital condyle has one developed lateral depression on each side. The basisphenoid has broad and short processes, a well-developed tuber and separates the large posterior cranial fenestrae. The opisthotic bears a small foramen. The pseudomesethmoid, seen in both specimens, is well-developed and has a thick lateral margin, similar to the condition reported in Tapejara20.
The dentary is the largest element of the lower jaw and shows a large mandibular symphysis (Table 1), with a pointed process on the tip (Supplementary Figs S2,S3). It is probable that the distal end of the mandibular symphysis showed some expansion, but due to flattening this cannot be confirmed in either specimen. On the ventral margin the dentary bears a well-developed, deep, blade-shaped and elongate bony sagittal crest that begins at the tip and extends to the end of the symphysis. It is larger than in any known pterosaur species (Figs. 2,4). This structure further differs from the crest observed in some tapejarids like Sinopterus, where it is less ventrally projected and less laterally compressed8. The crest is symmetrical, with the deepest point about half its length (between the 9th and 10th alveoli). The ventral edge is extraordinarily thin and might have been covered by a horny sheath. The crest also has a hook-like process on its posterior margin (Fig. 2). This quite unusual structure, unique to Ikrandraco avatar, can be observed in both specimens and is not an artifact.

Comparison between the dentary crest length in relation to the mandibular length in (a) Ikrandraco avatar, holotype IVPP V18199; (b) Ikrandraco avatar, referred specimen IVPP V18046; (c) Anhanguera piscator [after 17]; (d) Anhanguera robustus [after 17]; (e) Anhanguera spielbergi [after 18].
Not to scale. Line drawing by S.J.
There are at least 21 teeth on each side of the upper jaw and 19 in the lower. Alveoli are small and oval, without expressive size variation, contrary to several other toothed pteranodontoids, including species from the Jiufotang Formation10,11. Upper teeth 1–3 and lower 1–2 are closer to each other with the alveolar distance increasing gradually towards the posterior end. The enamel is smooth and dark-brown. The crowns are small, differing in this respect from anhanguerians21, with the anterior-most ones rounder and gently curved and posterior ones smaller and more flattened laterally. Teeth are considerably smaller than those of Guidraco and Liaoningopterus10,11 and those of any anhanguerid17. They are comparable in size with istiodactylids, but those pterosaurs have quite distinctive triangular and laterally compressed teeth3,13 not present in the new taxon.
Only a small part of the neural spine of the atlas is preserved, closely attached to the axis, suggesting that they were fused. The axis presents a tall and blade-like neural spine and the centrum pierced by two lateral pneumatic foramina (Fig. 3d), thus differing from anhanguerids, which have a spike-like spine and one large lateral pneumatic foramen16,17. The third cervical vertebra shows a well-developed lateral pneumatic foramen, a high and spike-like neural spine, large postexapophyses and no cervical ribs. Two remaining cervical elements, probably cervical vertebrae four and five, show no significant variation in centrum length compared to the third cervical.
The holotype of Ikrandraco avatar preserves partial wings. Although not articulated, they are closely associated. The left humerus is exposed ventroposteriorly (Fig. 3e). When compared to the length of the skull, it is smaller than in all other toothed pteranodontoids, but still larger than in the Nyctosauridae (Table 2). It shows a warped deltopectoral crest, with the proximal margin rounded, a typical pteranodontoid feature6. Both ulnae and the left radius are incomplete, but the right radius is well preserved and shows a diameter about half that of the ulna. The carpals are fused into proximal and distal syncarpals, the latter with a triangular outline (Supplementary Fig. S4). The pteroid is long and thin, with a preserved portion over half the length of the ulna. Both wing metacarpals are present. The left metacarpal I is an elongate element preserved in articulation with the distal syncarpal. A partially preserved thin bone could be right metacarpal II or III, indicating that metacarpals II and III were reduced, a common feature within derived pterodactyloids6,19. One manual ungual, preserved displaced inside the nasoantorbital fenestra, is comparatively high and short and is covered distally by a brown matter, likely the remains of a horny sheath. Both extensor tendon processes of the first wing phalanges are fused. A well-developed pneumatic foramen close to the anterior margin in both the second and third wing phalanges is observed as has been reported in Pteranodon19, showing a high degree of pneumatization in the wing skeleton of the new species, possibly by subcutaneous diverticulae of an air sac system22.
The only preserved elements of the hind limbs are the metatarsals present in the holotype (IVPP V18199). Despite being incomplete, it is clear that the metatarsal IV is stouter than the others, with two very unusual depressions that might be puncture marks.
Phylogenetic position
A phylogenetic analysis was performed (see Methods and Supplementary Information) and recovered Ikrandraco avatar among the Pteranodontoidea, outside but closely related to the Anhangueria (Fig. 5). The new species belongs to the Pterodactyloidea based on the presence of a confluent naris and antorbital fenestra (also present in wukongopterids) and several postcranial characters including the lack of ribs on mid-cervical vertebrae6. Although the inclination of the quadrate is more similar to the Archaeopterodactyloidea6, the new species possesses features typical of pteranodontoids such as a warped deltopectoral crest of the humerus and can be confidently assigned to that clade.

The strict consensus tree of the 18 equally parsimonious trees with 211 steps each.
Discussion
Both specimens were found in two localities about 20–25 km apart and their exact stratigraphic level is not known. Therefore, there is a potential that the strata from which they belong were of distinct ages, which could suggest that they represent distinct species, what would have to be supported by anatomical differences23. The referred specimen (IVPP V18406) is about 6% smaller than the holotype, but both have similar proportions of the nasoantorbital fenestra and of the rostrum relative the skull length. The referred specimen has comparatively deeper dentary crest, but in both specimens, the deepest point of the crest lies between the 9th and 10th lower jaw alveoli. Other distinctions are the height of the skull and the height of the mandibular rami, which are lower in the holotype than in the referred specimen and here mainly attributed as the result of taphonomic compression.
One of the most outstanding features of Ikrandraco avatar is the presence of a blade-like bony median crest at the anterior portion of the lower jaw. Several pterosaur species are known to bear crests, either bony or formed by a combination of bone and soft tissue. However, Ikrandraco avatar is unique by possessing a crest only on the mandible and none on the skull. Until now, blade-like crests were known exclusively in the Anhangueria and in Cimoliopterus cuvieri21. Ludodactylus sibbicki was reported to bear a dentary crest24, however, upon personal examination of the holotype, no crest could be observed. Cearadactylus atrox possesses a crest only on the premaxilla25 and the presence of a head crest, but none in the mandible, is more widely distributed within pterosaurs [e.g.3,11,26,27,].
With mounting evidence of the presence of crests in pterosaurs both basal and derived, their function is presently a contentious and hotly debated subject. Proposals range from sexual dimorphism (either presence × absence28,29 and size differences between genders23,30), mutual sexual selection, thermoregulation, species recognition, aerodynamics and biomechanical functions [e.g.30,31,32,33,34].
The potential functions of cranial crests that are limited to the rostral ends have been far less discussed than, for instance, the extremely enlarged cranial structures present in the Pteranodontidae and the Tapejaridae19,30. Admittedly, to infer functions on structures found in fossils that have not left recent descendants is a difficult task, particularly if modern analogs are also lacking. This is the case of the odd cranial structures of several pterosaurs, including the one observed in Ikrandraco avatar. Regardless of the primary force responsible for the evolution of head crests, it is likely that they played multiple roles30. Even if biomechanical purposes were not the driving force for the development of these structures, once present it is conceivable that they might have helped the animal in its daily activities.
The dentary crest in Ikrandraco is unusually long and deep. Although it cannot be ruled out that it might have been used as a display structure, involved in sexual display32,33 or in species recognition34, it seems odd for an animal to evolve such a bony projection in the lower jaw and not on the dorsal surface of the skull, where it would be far more visible. Perhaps if the animal would raise its head, this large dentary crest, covered with skin of bright colors might have had a display function, a difficult scenario to test when it comes to a fully extinct and distinctive group of flying animals such as pterosaurs. Furthermore, the dentary sagittal crest is very elongate, deep and laterally compressed. It also has a very sharp ventral margin, which is not a taphonomic artifact and differs from the more blunt condition of some other crested pterosaurs, particularly the Anhangueridae (Fig. 4). It further presents a unique hook-shaped process at the posterior margin, a novel and unusual structure, to our knowledge never documented in any vertebrate before. The medial position of this structure is consistent with the notion that it could have served as an anchor for soft tissues. Being under the chin and just in front the hyoid bones, it could be possibly related to some sort of throat sac.
The presence of throat sacs in pterosaurs has been proposed several times before, based primarily on soft tissue impressions in specimens of Rhamphorhynchus and Pterodactylus35,36. In all described cases, the pouch starts at the posterior ventral part of the mandible and extends until the level of the third or fourth cervical vertebrae, but its exact properties and morphology are disputed. While some researchers have suggested a structure similar to that seen in pelicans37, others have used the more neutral term “loose extensible skin”, arguing that this gular structure might have helped them to swallow large prey items, as seen in the ostriches38. None of the pterosaurs where they are thought to be present show the hook-shaped dentary process present in Ikrandraco avatar. It is possible that a gular structure potentially made of extensible skin was more developed in the new species.
These anatomical features, allied with a low and elongate fusiform and streamline skull, suggests that Ikrandraco avatar could have foraged by flying low over the water and, upon visually locating a prey item near the surface, could have lowered the jaw partially immersing it. The deep and laterally compressed dentary crest with a sharp ventral margin would break the water resistance, allowing a temporary skimming activity. When the prey item was closer, the animal could open the jaws and capture the submersed prey, emerging the jaws from the water surface. It is conceivable that this crest might also have been covered by a smooth horny sheath, a structure already reported in other pterosaurs [e.g.24], which could potentially reduce drag. The presence of an analog to the avian throat pouch, anchored in the hook-shaped medial process of the dentary crest, could have acted as a storing device for the prey items, allowing the animal to perform subsequent captures before swallowing, and/or help to expel the water that might have gotten with the recently captured prey. It should be noted that the skimming-like behavior suggested here would not have been used extensively during foraging because Ikrandraco avatar lacks the extensive adaptations reported for recent skimmers39 and also for the tapejarid Thalassodromeus sethi, that is also regarded to have been able to skim [30, but see40]. Ikrandraco lived near large freshwater bodies represented by the Jiufotang Formation, where several small fishes have been found41 and could potentially have been the prey item of this pterosaur.
Lastly, it is interesting to point out that Ikrandraco avatar comes from the same deposit as the anhanguerid Liaoningopterus gui and Guidraco venator, the latter from the same locality as the referred specimen IVPP V18406. Both are regarded as piscivorous but differ from the new species by the possession of much larger and more robust teeth10,11. Therefore, the distinct morphologies and potential different foraging habits of these species suggest different prey preferences and therefore distinct resource exploitation.
Ikrandraco avatar is a new genus and species of a pteranodontoid pterosaur from the Jiufotang Formation of China and is one of the few pterosaur species known by more than one specimen. It is closely related to the Anhangueria, a clade formed by purported piscivorous pterosaurs. The new species is diagnosed by a suite of features, but most notably by a blade-like, deep and elongate dentary crest, which possesses an unusual hook-shaped process at the posterior margin and a low and elongate skull that lacks a premaxillary crest. We propose that this pterosaur fed on fishes from nearby freshwater lakes by flying low over the water, capturing its prey by lowering the mandible in the water, being capable of a reduced and temporary skimming. We also propose that it had a more developed throat pouch then in other pterosaur species. Ikrandraco avatar foraging habit increases the potential feeding modes of pterosaurs.
Methods
Specimens IVPP V18199 and IVPP V18406 are deposited at the Institute of Vertebrate Paleontology and Paleoanthropology in Beijing, China, where they were first-hand analyzed. In order to assess the phylogenetic position of Ikrandraco avatar, an analysis was performed based on a modified version of a previously published matrix21. Analysis was performed on T.N.T. (Tree Analysis Using New Technology), version 1.1 for Windows with a menu interface – standard version42. All characters were unordered and given the same weight. Ornithosuchus longidens was considered the primary outgroup. Analysis was performed using the traditional search, with the TBR algorithm (although the software uses both TBR and SPR), with 5,000 random addition sequence replicates. Branches were collapsed when their minimum length was equal to zero. As some replications overflowed after a first round, traditional search was again employed, this time on trees from RAM. The analysis resulted in 18 equally parsimonious trees with 211 steps each. From these, a strict consensus tree was obtained and is here figured.
Change history
11 August 2020
An amendment to this paper has been published and can be accessed via a link at the top of the paper.
References
Kellner, A. W. A. Pterossauros: Os Senhores Dos Céus Do Brasil. (Vieira & Lent, Rio de Janeiro, 2006).
Barrett, P. M., Butler, R. J., Edwards, N. P. & Milner, A. R. Pterosaur distribution in time and space: an atlas. Zitteliana B 28, 61–107 (2008).
Wang, X., Kellner, A. W. A., Zhonghe, Z. & Campos, D. A. Pterosaur diversity and faunal turnover in Cretaceous terrestrial ecosystems in China. Nature 437, 875–879 (2005).
Butler, R. J., Barrett, P. M., Nowbath, S. & Upchurch, P. Estimating the effects of sampling biases on pterosaur diversity patterns: implications for hypotheses of bird/pterosaur competitive replacement. Paleobiology 35, 432–446 (2009).
He, H. et al. Timing of the Jiufotang Formation (Jehol Group) in Liaoning, northeastern China and its implications. Geophys. Res. Lett. 31, L12605 (2004).
Kellner, A. W. A. Pterosaur phylogeny and comments on the evolutionary history of the group. Evolution and Palaeobiology of Pterosaurs. Buffetaut, E. & Mazin, J.-M. (eds.) 105–137 (Geological Society, London, 2003).
Kellner, A. W. A. et al. The largest flying reptile from Gondwana: a new specimen of Tropeognathus cf. T. mesembrinus Wellnhofer, 1987 (Pterodactyloidea, Anhangueridae) and other large pterosaurs from the Romualdo Formation, Lower Cretaceous, Brazil. An. Acad. Bras. Cienc. 85, 113–135 (2013).
Wang, X. & Zhou, Z. A new pterosaur (Pterodactyloidea, Tapejaridae) from the Early Cretaceous Jiufotang Formation of western Liaoning, China and its implications for biostratigraphy. Chin. Sci. Bull. 47, 1521–1528 (2002).
Lü, J., Gao, Y., Xing, L., Li, Z. & Ji, Q. A new species of Huaxiapterus (Pterosauria: Tapejaridae) from the Early Cretaceous of western Liaoning, China. Acta Geol. Sin. 81, 683–687 (2007).
Wang, X. & Zhou, Z. Two new pterodactyloid pterosaurs from the Early Cretaceous Jiufotang Formation of western Liaoning, China. Vert. PalAsiat. 41, 34–41 (2003).
Wang, X., Kellner, A. W. A. Jiang, S. & Cheng, X. New toothed flying reptile from Asia: close similarities between early Cretaceous pterosaur faunas from China and Brazil. Naturwissenschaften 99, 249–257 (2012).
Sayão, J. M., Vila Nova, B. & Romano, P. S. R. Considerações sobre os padrões de distribuição geográfica dos Pterodactyloidea (Archosauria, Pterosauria). Paleontologia De Vertebrados: Relações Entre América Do Sul E África. Gallo, V., Silva, H. M. A., Brito, P. M. & Figueiredo, F. J. (eds) 281–300 (Interciência, Rio de Janeiro, 2012).
Andres, B. & Ji, Q. A new species of Istiodactylus (Pterosauria, Pterodactyloidea) from the Lower Cretaceous of Liaoning, China. J. Vertebr. Paleontol. 26, 70–78 (2006).
Lü, J., Unwin, D. M., Xu, L. & Zhang, X. A new azhdarchoid pterosaur from the Lower Cretaceous of China and its implications for pterosaur phylogeny and evolution. Naturwissenschaften 95, 891–897 (2008).
Wang, X., Kellner, A. W. A., Zhou, Z. & Campos, D. A. Discovery of a rare arboreal forest-dwelling flying reptile (Pterosauria, Pterodactyloidea) from China. Proc. Natl. Acad. Sci. U. S. A. 105, 1983–1987 (2008).
Wellnhofer, P. Weitere Pterosaurierfunde aus der Santana–Formation (Apt) der Chapada do Araripe, Brasilien. Palaeontogr. Abt. A 215, 43–101 (1991).
Kellner, A. W. A. & Tomida, Y. Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian–Albian), northeastern Brazil. Natl. Sci. Mus. Monogr. 17, 1–135 (2000).
Veldmeijer, A. J. Description of Coloborhynchusspielbergi sp. nov. (Pterodactyloidea) from Brazil. Scr. Geol. 125, 35–139 (2003).
Bennett, S. C. The osteology and functional morphology of the Late Cretaceous pterosaur Pteranodon. Part I. General description of osteology. Palaeontogr. Abt. A 260, 1–112 (2001).
Kellner, A. W. A. Description of the braincase of two Early Cretaceous pterosaurs (Pterodactyloidea) from Brazil. Am. Mus. Novit. 3175, 1–34 (1996).
Rodrigues, T. & Kellner, A. W. A. Taxonomic review of the Ornithocheirus complex (Pterosauria) from the Cretaceous of England. ZooKeys 308, 1–112 (2013).
Claessens, L. P. A. M., O'Connor, P. M. & Unwin, D. M. Respiratory evolution facilitated the origin of pterosaur flight and aerial gigantism. PLoS ONE 4, e4497 (2009)
Kellner, A. W. A. Comments on the Pteranodontidae (Pterosauria, Pterodactyloidea) with the description of two new species. An. Acad. Bras. Cienc. 82, 1063–1084 (2010).
Frey, E., Martill, D. M. & Buchy, M.-C. A new crested ornithocheirid from the Lower Cretaceous of northeastern Brazil and the unusual death of an unusual pterosaur. Evolution and Palaeobiology of Pterosaurs. Buffetaut, E. & Mazin, J.-M. (eds.) 55–63 (Geological Society, London, 2003).
Vila Nova, B. C., Sayão, J. M., Neumann, V. H. M. L. & Kellner, A. W. A. Redescription of Cearadactylus atrox (Pterosauria, Pterodactyloidea) from the Early Cretaceous Romualdo Formation (Santana Group) of the Araripe Basin, Brazil. J. Vertebr. Paleontol. 34, 126–134 (2014).
Young, C. C. On a new pterosaurian from Sinkiang, China. Vertebrat. PalAsiatic. 8, 239–255 (1964).
Dalla Vecchia, F. M., Wild, R., Hopf, H. & Reitner, J. A crested rhamphorhynchoid pterosaur from the Late Triassic of Austria. J. Vertebr. Paleontol. 22, 196–199 (2002).
Bennett, S. C. Sexual dimorphism of Pteranodon and other pterosaurs, with comments on cranial crests. J. Vertebr. Paleontol. 12, 422–434 (1992).
Lü, J. et al. An egg-adult association, gender and reproduction in pterosaurs. Science 331, 321–324 (2011).
Kellner, A. W. A. & Campos, D. A. The function of the cranial crest and jaws of a unique pterosaur from the Early Cretaceous of Brazil. Science 297, 389–392 (2002).
Bennett, S. C. Soft tissue preservation of the cranial crest of the pterosaur Germanodactylus from Solnhofen. J. Vertebr. Paleontol. 22, 43–48 (2002).
Hone, D. W. E., Naish, D. & Cuthill, I. C. Does mutual sexual selection explain the evolution of head crests in pterosaurs and dinosaurs? Lethaia 45, 139–156 (2012).
Padian, K. & Horner, J. R. Misconceptions of sexual selection and species recognition: a response to Knell et al. and to Mendelson and Shaw. Trends Ecol. Evol. 28, 249–250 (2013).
Frey, E. & Martill, D. M. Soft tissue preservation in a specimen of Pterodacylus kochi (Wagner) from the Upper Jurassic of Germany. Neues. Jahrb. Geol. Palaontol.-Abh. 210, 421–441 (1998).
Knell, R. J., Naish, D., Tomkins, J. L. & Hone, D. W. E. Sexual selection in prehistoric animals: detection and implications. Trends Ecol. Evol. 28, 38–47 (2013).
Wellnhofer, P. Die Flughaut von Pterodactylus (Reptilia, Pterosauria) am Beispiel des Wiener Exemplares von Pterodactylus kochi (Wagner). Ann. Naturhist. Mus. Wien. A 88, 149–162 (1987).
Averianov, A. O. Reconstruction of the neck of Azhdarcho lancicollis and lifestyle of azhdarchids (Pterosauria, Azhdarchidae). Paleontol. J. 47, 203–209 (2013).
Bennett, S. C. The osteology and functional morphology of the Late Cretaceous pterosaur Pteranodon. Part II. Size and functional morphology. Palaeontogr. Abt. A 260, 113–153 (2001).
Zhou, Z., Barrett, P. M. & Hilton, J. An exceptionally preserved Lower Cretaceous ecosystem. Nature 421, 807–814 (2003).
Zusi, R. L. Structural adaptations of the head and neck in the black skimmer Rynchops nigra Linnaeus. Nuttal. Ornithol. Club 3, 1–101 (1962).
Humphries, S., Bonser, R. H. C., Witton, M. P. & Martill, D. M. Did pterosaurs feed by skimming? Physical modelling and anatomical evaluation of an unusual feeding method. PLoS Biol. 5, e204 (2007).
Goloboff, P. A., Farris, J. S. & Nixon, K. C. TNT, a free program for phylogenetic analysis. Cladistics 24, 774–786 (2008).
Bennett, S. C. New crested specimens of the Late Cretaceous pterosaur Nyctosaurus. Palaeontology 77, 61–75 (2003).
Lü, J., Xu, L., Chang, H. & Zhang, X. A new darwinopterid pterosaur from the Middle Jurassic of western Liaoning, northeastern China and its ecological implications. Acta Geol. Sin. 85, 507–514 (2011).
Wellnhofer, P. Die Pterodactyloidea (Pterosauria) der Oberjura-Plattenkalke Süddeutschlands. Abh. Bayer. Akad. Wiss., N. F. 141, 1–133 (1970).
Lü, J., Unwin, D. M., Jin, X., Liu, Y. & Ji, Q. Evidence for modular evolution in a long-tailed pterosaur with a pterodactyloid skull. Proc. R. Soc. B, Biol. Sci. 277(1680), 383–389 (2010).
Wang, X. et al. New long-tailed pterosaurs (Wukongopteridae) from western Liaoning, China. An. Acad. Bras. Cienc. 82(4), 1045–1062 (2010).
Lü, J. A new boreopterid pterodactyloid pterosaur from the Early Cretaceous Yixian Formation of Liaoning Province, northeastern China. Acta Geol. Sin. 84, 241–246 (2010).
Acknowledgements
We thank L Xiang and H Zhou for the preparation of the specimens and W Gao for the photos. Brian Choo brought the name Ikran to our attention. This study was supported by the National Science Fund for Distinguished Young Scholars (40825005), the National key Basic Research Program of China (2012CB821900) and the Hundred Talents Project of CAS. TR received funding from the Conselho Nacional de Desenvolvimento Científico e Tecnológico and Fundação de Amparo à Pesquisa do Espírito Santo (CNPq/FAPES # 52986870/2011) and AWAK acknowledges founding from the Fundação Carlos Chagas Filho de Amparo à Pesquisa do Rio de Janeiro (FAPERJ # E-26/102.737/2012) and the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq # 304780/2013-8).
Author information
Authors and Affiliations
Contributions
X.W., T.R. and A.K. wrote the main manuscript; X.W. and S.J. found the specimens, S.J. prepared the figures; X.W., T.R., S.J., X.C. and A.K. contributed to the manuscript and research. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Supplementary Information
Supplementary Information
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/4.0/
About this article

Cite this article
Wang, X., Rodrigues, T., Jiang, S. et al. An Early Cretaceous pterosaur with an unusual mandibular crest from China and a potential novel feeding strategy. Sci Rep 4, 6329 (2014). https://doi.org/10.1038/srep06329
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep06329
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
