
Abstract
To clarify the effect of deep rooting on grain yield in rice (Oryza sativa L.) in an irrigated paddy field with or without fertilizer, we used the shallow-rooting IR64 and the deep-rooting Dro1-NIL (a near-isogenic line homozygous for the Kinandang Patong allele of DEEPER ROOTING 1 (DRO1) in the IR64 genetic background). Although total root length was similar in both lines, more roots were distributed within the lower soil layer of the paddy field in Dro1-NIL than in IR64, irrespective of fertilizer treatment. At maturity, Dro1-NIL showed approximately 10% higher grain yield than IR64, irrespective of fertilizer treatment. Higher grain yield of Dro1-NIL was mainly due to the increased 1000-kernel weight and increased percentage of ripened grains, which resulted in a higher harvest index. After heading, the uptake of nitrogen from soil and leaf nitrogen concentration were higher in Dro1-NIL than in IR64. At the mid-grain-filling stage, Dro1-NIL maintained higher cytokinin fluxes from roots to shoots than IR64. These results suggest that deep rooting by DRO1 enhances nitrogen uptake and cytokinin fluxes at late stages, resulting in better grain filling in Dro1-NIL in a paddy field in this study.
Similar content being viewed by others

Introduction
Plant roots are important organs for the uptake of water and nutrients from the surrounding soil. The physiological and morphological traits of roots affect shoot growth. In rice (Oryza sativa L.), a well-developed root system increases biomass and yield in different treatments and cultivars in paddy fields1,2,3,4,5,6,7,8. Higher uptake of nitrogen and water during the grain filling stage allows the plant to attain higher dry matter production by maintaining a higher photosynthetic rate in paddy fields1,5,6,7,9. High flux of cytokinins, which are synthesized in the roots and transported through the xylem to the shoot10,11,12,13,14, contributes to the maintenance of high photosynthetic rate owing to increasing leaf nitrogen and ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) which is a key enzyme in photosynthesis in rice6,9,15,16. High-yielding rice plants often have a larger volume of downward-growing nodal roots, which increases the proportion of roots in the lower soil layer in paddy fields17,18,19,20. However, the importance of these root ideotypes has been suggested on the basis of comparison among cultivars with different root and shoot morphologies and it is not clear whether the deep root system contributes to high yield and dry matter production in paddy fields. To accurately evaluate the effect of deep rooting on rice grain yield, homogeneous genetic materials such as near-isogenic lines (NILs) with differences only in the root growth angle are needed.
Recently, Uga et al.21 cloned and characterized a quantitative trait locus (QTL) for root growth angle in rice, DEEPER ROOTING 1 (DRO1). DRO1 functions downstream of the auxin signaling pathway and controls the gravitropic curvature in rice roots. The authors also developed Dro1-NIL, carrying a functional allele of DRO1 derived from the deep-rooting cultivar Kinandang Patong (a traditional tropical japonica upland cultivar of Philippine origin) in the genetic background of the shallow-rooting parent variety IR64 (a modern lowland indica cultivar, widely grown in South and Southeast Asia), which has a non-functional allele of DRO1. Under upland conditions, Dro1-NIL showed deeper rooting than IR64, although the two lines did not show any marked differences in other root and shoot traits21,22. Under upland conditions with drought stress, Dro1-NIL maintained better grain filling and thus showed higher yield than IR64 because the plants avoided drought stress by collecting water from deep soil. This clearly demonstrated that deep rooting is a beneficial trait for stable rice production under drought conditions. Thus, we assumed that Dro1-NIL might be a useful material to clarify the effect of deep roots in paddy fields.
Lodging also affects grain yield and dry matter production23. Although previous studies examined the relationship between deep rooting and lodging resistance, they compared different cultivars or the effects of root pruning24,25,26,27. Such comparisons cannot exclude the effects of genotype differences or root injury, respectively, on the observed effects.
The objective of this study was to examine the effect of deep rooting on grain yield in paddy fields by using the deep-rooting Dro1-NIL and the shallow-rooting IR64. We evaluated not only yield and its components, but also physiological traits such as nitrogen uptake from roots and cytokinin fluxes in xylem exudates and leaves. We also investigated the effect of deep-rooting Dro1-NIL on lodging resistance.
Results
Root length density
There was no significant difference in total root length between IR64 and Dro1-NIL, irrespective of fertilizer treatment (Table 1). In both lines, root length density decreased with increasing depth. Root length density differed significantly between the lines when measured in specific soil layers, although there was no interaction between the effects of the line and fertilizer. Root length density of Dro1-NIL was 0.79 times that of IR64 in the upper layer, 1.54 times in the middle layer and 1.24 times in the lower layer, irrespective of fertilizer treatment.
Dry matter production, yield and yield components
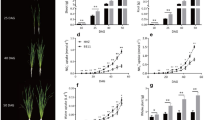
The dry weight of the above-ground parts was similar in both lines at heading, but was higher in Dro1-NIL at maturity, irrespective of fertilizer treatment (Table 2). Grain yield and HI were also higher in Dro1-NIL than in IR64, irrespective of fertilizer treatment. The number of panicles per m2, the number of spikelets per panicle and the number of total spikelets per m2 were similarly increased by fertilizer treatment in both lines (Table 3). In contrast, the 1000-kernel weight and percentage of filled spikelets were higher in Dro1-NIL than in IR64, irrespective of fertilizer treatment.
Nitrogen concentration in leaves and nitrogen content of plants
At heading, nitrogen concentration in flag leaves and total nitrogen content of plants were similar in the two lines (Tables 4 and 5). On the other hand, at 20 days after heading, nitrogen concentration in flag leaves was higher in Dro1-NIL than in IR64, irrespective of fertilizer treatment (Table 4). At maturity, total nitrogen content of plants was also higher in Dro1-NIL than in IR64, irrespective of fertilizer treatment (Table 5). As a result, nitrogen accumulated during grain filling was higher in Dro1-NIL than in IR64 with or without fertilizer treatment, although nitrogen concentration in leaves and nitrogen content of plants were increased by fertilizer treatment in both lines (Tables 4 and 5).
Xylem exudation rate, cytokinin flux in xylem exudates and cytokinin concentration in flag leaves
At 14 days after heading, the rate of xylem exudates and the flux of cytokinins (determined as trans-zeatin, cis-zeatin, trans-zeatin riboside, N6(Δ2-isopentenyl)adenine, N6(Δ2-isopentenyl)adenosine, dihydrozeatin and dihydrozeatin riboside combined) in xylem exudates, as well as concentrations of these cytokinins in flag leaves, were higher in Dro1-NIL than in IR64, irrespective of fertilizer treatment, although both parameters were increased by fertilizer treatment in both lines (Table 6).
Lodging resistance
At 14 days after heading, pushing resistance was higher in Dro1-NIL than in IR64 (Table 7). The difference between the two lines was no longer significant at 35 days after heading.
Discussion
The roots of Dro1-NIL were distributed in the lower soil layer than those of IR64 in a paddy field (Table 1, this study), similar to previous observations in upland fields21. Recently, we have demonstrated that Dro1-NIL produces more filled grains than IR64 under drought stress conditions, but not under non-drought conditions21. These results were obtained in upland fields. Thus, we wondered whether yield performance differs between shallow- and deep-rooting varieties in paddy fields.
In a paddy field, grain yield of the Dro1-NIL was higher than that of IR64, irrespective of fertilizer treatment (Table 2). Under drought conditions, deeper rooting caused by DRO1 led to a lower percentage of seed sterility than in shallow-rooted IR64 and thus resulted in higher yield, because plants avoided drought stress by increasing water capture from deep soil21. In a paddy field, higher grain yield of the Dro1-NIL was mainly due to the higher 1000-kernel weight and percentage of ripened grains, resulting in better grain filling and higher HI (Tables 2 and 3). This suggests that the Dro1-NIL may have greater ability to supply carbohydrates to the panicle after heading.
Plant carbohydrates are produced by photosynthesis. Nitrogen is essential for the production of photosynthetic enzymes and there is a positive relationship between leaf nitrogen content and the rate of photosynthesis6,9,28. Recently, Takai et al.29 provided genetic evidence suggesting that photosynthetic rate is positively related to leaf nitrogen content through the cloning and characterization of GPS (GREEN FOR PHOTOSYNTHESIS), a rice QTL that regulates carboxylation efficiency and thus controls the photosynthetic rate. In this study, nitrogen concentration in flag leaves (Table 4) and the other leaves (data not shown) after heading were higher in Dro1-NIL than in IR64, irrespective of fertilizer treatment, which may conduce better grain filling in Dro1-NIL. We hypothesize that increase in nitrogen uptake by DRO1 enhances photosynthetic rate, resulting in higher grain filling ability. However, we did not investigate the photosynthesis of leaves in both lines of this study. Therefore, the relationship between photosynthesis and grain filling remains to be investigated.
Nitrogen is supplied to rice from both fertilizer and mineralized soil nitrogen30. Because nitrogen in the upper soil layer is often insufficient at the late growth stage, efficient uptake of nitrogen from the lower layer is important for vigorous growth at this stage31. A significant difference in plant dry weight and nitrogen uptake between IR64 and Dro1-NIL at maturity, but not at heading, suggests that higher root density of Dro1-NIL in the lower soil layer in comparison with IR64 was probably advantageous in capturing soil nitrogen from these layers during the grain filling period. However, the rate of nitrogen uptake from the lower soil layer still needs to be accurately measured to clarify how nitrogen absorbed from the lower soil layer in rice carrying DRO1 assists increasing grain yield. We observed several roots of Dro1-NIL penetrated through the plow pan layer (data not shown), although we did not quantify roots and nitrogen existed below 20 cm in the soil layer. We also have to elucidate effect of deep roots below 20 cm of soil layer on nitrogen uptake.
Cytokinins are involved in the control of leaf senescence32,33. The delay in leaf senescence is considered to be important for increased dry matter and grain production in paddy rice1,5,6,7,9,34. Higher transport of cytokinins from roots to shoots during grain filling is also associated with the maintenance of higher leaf nitrogen and Rubisco and thus higher photosynthetic rate6,9,15,16. In this study, both cytokinin flux in exudates and cytokinin concentration in flag leaves were higher in the Dro1-NIL than in IR64, irrespective of fertilizer treatment (Table 6). These results indicate that DRO1 plays an important role in elevating the cytokinin level. We suppose that this higher cytokinin level of Dro1-NIL was associated with maintenance of higher leaf nitrogen and delaying leaf senescence. How is Dro1-NIL able to maintain higher cytokinin levels during grain filling? The roots distributed in the lower layer were white and fresh in comparison with the roots distributed in the upper soil layer, which implies a non-reducing environment around the plow pan layer. We also observed that roots of Dro1-NIL penetrated through the plow pan layer were white and fresh (data not shown). Therefore, we hypothesize that elevated cytokinin synthesis in Dro1-NIL after heading was due to its vigorous roots in the lower soil layer and the layer beneath the plow pan. The mechanisms of high cytokinin flux in rice plants with roots distributed in the lower layer and its effects on photosynthesis and yield performance remain to be clarified.
Lodging reduces grain yield and dry matter production in paddy rice35. In this study, Dro1-NIL showed higher pushing resistance as an index of lodging resistance than IR64 before the soil hardness increased as a result of the drainage of residual water in the paddy field, although lodging no longer occurred after heading in either line (Table 7). This suggests that roots distributed in the lower layer contributed to the increased root lodging resistance in wet soil. Because the position of the shoot base of rice plants directly sown in the soil is shallower than that of transplanted rice plants, lodging easily occurs after heading in direct-sowing cultivation27, resulting in a reduction in grain yield and dry matter production23. The roots penetrating into the lower soil layer with a higher bulk soil density have been proposed to be an important determinant of lodging resistance in rice in direct sowing cultivation25,26,27. We have now demonstrated that introduction of the deep rooting trait into shallow-rooting cultivars can form the basis of a breeding strategy for lodging resistance.
Under upland conditions, the maximum root depth of the Dro1-NIL was more than 40 cm while that of IR64 was around 20 cm21. The plow pan was 20 cm below the soil surface in our paddy field. The root growth of Dro1-NIL might be affected by the plow pan but some roots penetrated through the plow pan layer. Since we did not measure a penetration ability of roots in both lines, whether DRO1 improved the penetration ability of roots is unclear. It is necessary to further investigate the effect of DRO1 on penetration ability to determine if roots that penetrate through the plow pan have some benefits with respect to nitrogen uptake and cytokinin fluxes.
Our results suggest that deep rooting caused by the functional allele of DRO1 improves grain yield in a paddy field by enhancing the higher 1000-kernel weight and percentage of ripened grains. DRO1 predominantly influences root growth angle rather than other root morphological traits such as root length and number21. Physiological and yield traits improved in the Dro1-NIL should be attributable to change of vertical root distribution by DRO1, although we have to elucidate relationships between higher nitrogen uptake and cytokinin fluxes by DRO1 and grain yield in the Dro1-NIL. Previous studies have reported QTLs associated with photosynthesis and grain yield near the DRO1 region on chromosome 936,37. Multiple QTLs for net photosynthesis rate were found around SSR marker RM410 at flowering and grain filling under both drought and well-watered field conditions36. Dixit et al.37 detected promising QTL, qDTY9.1B, for grain yield under paddy fields with drought stress in the interval between SSR markers RM24350 (15.4 Mb) to RM24390 (15.9 Mb), whereas this QTL did not show positive effect on grain yield under non-drought conditions. They supposed that qDTY9.1B was due to pleiotropic effect of DRO1. The IR64 allele of DRO1 was identified in several IR64 progenies showing shallow rooting but not in the ancestors of IR6421. Introducing the Kinandang Patong allele of DRO1 into these modern varieties might be improved their grain yield in the paddy fields, although potential effects of DRO1 on grain yield in other genetic backgrounds remain to be clarified. Thus, further analysis using the other genetic backgrounds will be needed to understand relationship between deep rooting by DRO1 and its pleiotropic effect.
In the developing countries lacking agricultural infrastructure, farmers may not be able to use sufficient amounts of nitrogen fertilizers in paddy fields. DRO1 may be useful in these areas because nitrogen uptake from the lower soil layer due to deep rooting would contribute to increasing grain yield. However, these effects by DRO1 would depend on the soil fertility and the method of fertilizer application in each paddy field. Multi-location trials for the Dro1-NIL will be needed to conclude whether deep rooting by DRO1 shows universally effective for grain yield in the paddy fields because our results were obtained from the one location with relatively small plot size.
Methods
Materials and plant cultivation
IR64 and Dro1-NIL were cultivated in a paddy field at the NICS in Tsukubamirai (lat 36°02′N, long 140°04′E), Ibaraki, Japan, in 2010 and 2011. Dro1-NIL was developed from a cross between IR64 and Kinandang Patong (as a donor of the functional allele of DRO1) by repeated backcrossing with IR64 and marker-assisted selection to eliminate non-target DRO1 regions21. The Dro1-NIL had only homozygous for the Kinandang Patong allele between 15.9 Mb (SSR marker RM24386) to 18.8 Mb (SSR marker RM242) on chromosome 9, indicating that more than 99% of genome in the Dro1-NIL was derived from IR64. Plants were grown at a density of 22.2 hills (15 cm × 30 cm) per m2, with one plant per hill. Germinated seeds were sown in nursery boxes in April and grown in a vinyl shelter. In May, when the fourth leaves were expanding, seedlings were transplanted into an irrigated paddy field after puddling. The experimental plants were laid out in a split-plot arrangement with or without chemical fertilizer, with three replications (9 m2 in 2010 and 28 m2 in 2011 for each replicate, with fertilizer treatment as the main plot and line as the sub-plot). For the fertilizer plot, chemical fertilizers were applied as a basal dressing at 9 g m−2 each of P2O5 and K2O and 12 g m−2 N (6 g LP40 and 6 g LP100); LP40 and LP100 are controlled release fertilizers releasing 80% of the total nitrogen content at a uniform rate up to 40 and 100 days after application, respectively, at 20–30°C. The soil type was the Gleyic Fluvisols and the soil contained 5.0 mg 100 g−1 available nitrogen in the upper layer (0–10 cm) and 4.4 mg 100 g−1 in the lower layer (10–20 cm), as determined by hot water extraction. The plow layer was approximately 20 cm from the soil surface and the lower layer formed a plow pan, which was approximately 10 cm thick. Daily mean, maximum and minimum air temperatures during growth period were 23.4°C, 30.4°C and 13.3°C in 2010 and 23.0°C, 29.7°C and 13.6°C in 2011, respectively. The heading dates (defined as the dates when 50% of plants were at heading) of both lines were August 16, 2010 and August 15, 2011 with fertilization and August 14, 2010 and August 13, 2011 without fertilization. Plants were harvested on September 26, 2010 and on September 25, 2011.
Measurements of root length density
To evaluate root length density of rice plants in paddy field, we used core sampler method, because the core sampler can minimize destruction of the soil structure of paddy field compared to other methods such as monolith sampler. Four hills with the normal number of panicles were collected from each replicate at 20 days after heading in 2010 and 2011. A soil core (50 mm in diameter and approximately 200 mm in depth from the soil surface to the plow pan) was collected from position between the rows by using a core sampler (DIK-110C, Daiki Rika Kogyo Co., Saitama, Japan). The soil cores were divided into three segments corresponding to the upper (0–5 cm), middle (5–10 cm) and lower (10–20 cm) layers. The segments were washed carefully to separate roots from soil. The root length was determined by using a root scanner (WinRHIZO, Regent Instruments, Inc., Quebec, Canada). The root length density in each segment was calculated as root length divided by volume of each segment.
Measurements of yield, yield components and plant dry weight
Plants were harvested from an area of 1.1 m2 in 2010 and 2.6 m2 in 2011 for each replicate. Fully ripened grains were selected by sieving through a 1.6-mm mesh and the moisture content was adjusted to 15%. The harvest index (HI) was calculated as the panicle dry weight divided by the total plant dry weight. To determine plant dry weight, three (in 2010) or eight (in 2011) hills were selected for each replicate. Each plant was separated into leaves, culm and leaf sheaths and panicles. Samples were dried in an oven at 80°C for 5 days and weighed.
Measurement of nitrogen in flag leaves and whole plants
Measurements were performed in 2011. Flag leaves were collected from 8 hills per replicate at heading and at 20 days after heading. Whole plants with the normal number of panicles were collected from 8 hills per replicate at heading and at maturity. Samples were dried in an oven at 80°C for 5 days, weighed and powdered in a mill (W-100; Ikeda Rika, Tokyo, Japan). The nitrogen concentration on a dry-weight basis in each sample was determined with the Sumigraph NCH-22 analyzer (Sumika Chemical Analysis Service Center, Tokyo, Japan) and nitrogen content was calculated from the dry weight and nitrogen concentration.
Measurement of xylem exudation rate and cytokinins in xylem exudate and flag leaves
Five hills with the normal number of panicles were collected per replicate at 14 days after heading in 2011 and flag leaves were collected from the main tillers. To obtain exudates, shoots were cut 10 cm above the soil surface and a polyethylene bag containing cotton wool was attached to each cut end with tape. Bags were detached 3 h later, sealed and weighed. The weight of the exudate was calculated by subtracting the weight of the bag and the cotton wool. Exudates were collected by squeezing the cotton wool using syringe and immediately stored at −80°C. Cytokinins were extracted using an automatic liquid handling system (MICROLAB STAR, Hamilton, NV, USA) for solid phase extraction and quantified by ultra-performance liquid chromatography (UPLC) coupled with a tandem quadrupole mass spectrometer (qMS/MS) equipped with an electrospray interface (ESI; UPLC-ESI-qMS/MS, UPLC-Xevo TQ-S, Waters, MA, USA) as described previously38. Cytokinin flux was calculated from the exudation rate and cytokinin concentration.
Measurement of the pushing resistance of hills
Pushing resistance can be used as an index of lodging resistance of rice plants27,39,40,41,42. A prostrate tester (DIK7400; Daiki Rika Kogyo Co., Saitama, Japan) was set perpendicularly to the shoot at 15 cm above the soil surface and pushing resistance was measured by pushing the plants to an angle of 45° from the vertical. Eight hills (with fertilization only) with the normal number of panicles per replicate were selected at 14 and 35 days after heading in 2011. The average number of panicles was 13.5 hill−1 for IR64 and 13.6 hill−1 for Dro1-NIL at 14 days after heading and 13.8 hill−1 for IR64 and 13.7 hill−1 for Dro1-NIL at 35 days after heading. Water in the paddy field was drained starting from 25 days after heading.
Statistical analysis
Statistical analyses were performed by using a general linear model in SPSS version 17.0 (IBM, Tokyo, Japan). Analysis of variance (ANOVA) was used to test the effects of line and fertilizer treatment on root traits, yield, yield components, dry weight of the above-ground parts and parameters of nitrogen and cytokinin. Line and fertilizer treatment were considered as fixed effects. Year and replication were considered as random effects for the former four parameters. ANOVA was also used to test the effects of line on pushing resistance. Replications were considered as random effects. Significant treatment effects (P <0.05 or 0.01) were explored by using Fisher's protected least significant difference test.
References
Jiang, C.-Z., Hirasawa, T. & Ishihara, K. Physiological and ecological characteristics of high yielding varieties in rice plants. II Leaf photosynthetic rates. Jpn. J. Crop Sci. 57, 139–145 (1988).
Kang, S.-Y., Morita, S. & Yamazaki, K. Root growth and distribution in some Japonica-Indica hybrid and Japonica type rice cultivars under field conditions. Jpn. J. Crop Sci. 63, 118–124 (1994).
Kang, S.-Y. & Morita, S. Comparative analysis of root and shoot growth between Tongil and Japonica type rice. Korean J. Crop Sci. 43, 161–167 (1998).
Kumura, A. Physiology of high-yielding rice plants from the viewpoint of dry matter production and its partitioning. In:: Matsuo, T., Kumazawa, K., Ishii, R., Ishihara, K., Hirata, H. eds, Science of the rice plant 2. Physiology. Food and Agriculture Policy Research Center, Tokyo, pp 704–736 (1995).
San-oh, Y., Mano, Y., Ookawa, T. & Hirasawa, T. Comparison of dry matter production and associated characters between direct-sown and transplanted rice plants in a submerged paddy field and relationships to planting patterns. Field Crops Res. 87, 43–53 (2004).
San-oh, Y., Sugiyama, T., Yoshita, D., Ookawa, T. & Hirasawa, T. The effect of planting pattern on the rate of photosynthesis and related processes during ripening in rice plants. Field Crops Res. 96, 113–124 (2006).
San-oh, Y., Kondo, M., Ookawa, T. & Hirasawa, T. Ecophysiological analysis on effect of planting pattern on biomass production and grain yield in rice. JARQ 42, 79–89 (2008).
Taylaran, R. D. et al. Performance of a high-yielding modern rice cultivar Takanari and several old and new cultivars grown with and without chemical fertilizer in a submerged paddy field. Plant Prod. Sci. 12, 365–380 (2009).
Ookawa, T., Naruoka, Y., Yamazaki, T., Suga, Y. & Hirasawa, T. A comparison of the accumulation and partitioning of nitrogen in plants between two rice cultivars, Akenohoshi and Nipponbare, at the ripening stage. Plant Prod. Sci. 6, 172–178 (2003).
Kamada-Nobusada, T., Makita, N., Kojima, M. & Sakakibara, H. Nitrogen-dependent regulation of de novo cytokinin biosynthesis in rice: the role of glutamine metabolism as an additional signal. Plant Cell Physiol. 54, 1881–1893 (2013).
Letham, D. S. Cytokinins. in: Letham, D. S., Goodwin, P. B. & Higgins, T. J. V. (Eds.), Phytohormones and Related Compounds: A Comprehensive Treatise, Volume I. Elsevier/North-Holland Biomedical Press, Amsterdam, pp. 205–263 (1978).
Sakakibara, H. Cytokinins: Activity, biosynthesis and translocation. Annu. Rev. Plant Biol. 57, 431–449 (2006).
Short, K. C. & Torrey, J. G. Cytokinins in seedling roots of pea. Plant Physiol. 49, 155–160 (1972).
Van Staden, J. & Davey, J. E. The synthesis, transport and metabolism of endogenous cytokinins. Plant Cell Environ. 2, 93–106 (1979).
Ookawa, T., Naruoka, Y., Sayama, A. & Hirasawa, T. Cytokinin effects on ribulose-1,5-bisphosphate carboxylase/oxygenase and nitrogen partitioning in rice during ripening. Crop Sci. 44, 2107–2115 (2004).
Soejima, H., Sugiyama, T. & Ishihara, K. Changes in the chlorophyll contents of leaves and in levels of cytokinins in root exudates during ripening of rice cultivars Nipponbare and Akenohoshi. Plant Cell Physiol. 36, 1105–1114 (1995).
Morita, S., Iwabuchi, A. & Yamazaki, K. Relationships between the growth direction of primary roots and yield in rice plants. Jpn. J. Crop Sci. 56, 530–535 (1986).
Morita, S., Suga, T. & Yamazaki, K. The relationship between root length density and yield in rice plants. Jpn. J. Crop Sci. 57, 438–443 (1988).
Morita, S. Root system distribution and its possible relation to yield in rice. Low-input sustainable crop production systems in Asia. KSCS, Korea 371–377 (1993).
Kawata, S., Soejima, M. & Yamazaki, K. The superficial root function and yield of hulled rice. Jpn. J. Crop Sci. 47, 617–628 (1978).
Uga, Y. et al. Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions. Nat. Genet. 45, 1097–1102 (2013).
Uga, Y., Okuno, K. & Yano, M. Dro1, a major QTL involved in deep rooting of rice under upland field conditions. J. Exp. Bot. 62, 2485–2494 (2011).
San-oh, Y., Ookawa, T., Aizawa, N. & Hirasawa, T. Varietal differences in growth, lodging and related characters of rice plants broadcasted in submerged paddy field at different densities. Jpn. J. Crop Sci. 70, 515–524 (2001).
Sakata, I., Kagiya, T., Kawai, Y. & Oyanagi, A. Effects of pruning of roots growing at various angles on pushing resistance of rice cultivars. Jpn. J. Crop Sci. 73, 1–5 (2004).
Terashima, K., Ogata, T. & Akita, S. Eco-physiological characteristics related with lodging tolerance of rice in direct sowing cultivation II. Root growth characteristics of tolerant cultivars to root lodging. Jpn. J. Crop Sci. 63, 34–41 (1994).
Terashima, K., Akita, S. & Sakai, N. Eco-physiological characteristics related with lodging tolerance of rice in direct sowing cultivation III. Relationship between the characteristics of root distribution in the soil and lodging tolerance. Jpn. J. Crop Sci. 64, 243–250 (1995).
Terashima, K. Eco-physiological study of root lodging tolerance in direct-seeded rice cultivars. JARQ 31, 155–162 (1997).
Makino, A., Mae, T. & Ohira, K. Differences between wheat and rice in the enzymic properties of ribulose-1,5-bisphosphate carboxylase/oxygenase and the relationship to photosynthetic gas exchange. Planta 174, 30–38 (1988).
Takai, T. et al. A natural variant of NAL1, selected in high-yield rice breeding programs, pleiotropically increases photosynthesis rate. Sci. Rep. 3, 2149 (2013).
De Datta, S. K. Chemical changes in submerged rice soils. In: Principles and practices of rice production. John Wiley & Sons, New York, pp 89–145 (1981).
Toriyama, K. Soil and plant science now developed from field study – Acquisition and analysis of data from the new view point. 1. Growth of rice plant in the larger size paddy field and the variability in soil nitrogen fertility. Jpn. J. Soil Sci. Plant Nut. 72, 453–458 (2001).
Buchanan-Wollaston, V. et al. The molecular analysis of leaf senescence — A genomics approach. Plant Biotech. J. 1, 3–22 (2003).
Gan, S. & Amasino, R. M. Inhibition of leaf senescence by autoregulated production of cytokinin. Science 270, 1986–1987 (1995).
Jiang, C.-Z., Hirasawa, T. & Ishihara, K. Physiological and ecological characteristics of high yielding varieties in rice plants. I Yield and dry matter production. Jpn. J. Crop Sci. 57, 132–138 (1988).
Kono, M. Physiological aspects of lodging. In:: Matsuo, T., Kumazawa, K., Ishii, R., Ishihara, K., Hirata, H. eds, Science of the rice plant 2. Physiology. Food and Agriculture Policy Research Center, Tokyo, pp 971–982 (1995).
Gu, J. et al. Using chromosome introgression lines to map quantitative trait loci for photosynthesis parameters in rice (Oryza sativa L.) leaves under drought and well-watered field conditions. J. Exp. Bot. 63, 455–469 (2012).
Dixit, S. et al. Fine mapping of QTLs for rice grain yield under drought reveals sub-QTLs conferring a response to variable drought severities. Theor. Appl. Genet. 125, 155–169 (2012).
Kojima, M. et al. Highly sensitive and high-throughput analysis of plant hormones using MS-probe modification and liquid chromatography–tandem mass spectrometry: an application for hormone profiling in Oryza sativa. Plant Cell Physiol. 50, 1201–1214 (2009).
Kashiwagi, T. & Ishimaru, K. Identification and functional analysis of a locus for improvement of lodging resistance in rice. Plant Physiol. 134, 676–683 (2004).
Terashima, K., Akita, S. & Sakai, N. Eco-physiological characteristics related with lodging tolerance of rice in direct sowing cultivation. Jpn. J. Crop Sci. 61, 380–387 (1992).
Terashima, K., Taniguchi, T., Ogiwara, H. & Umemoto, T. Effect of field drainage on root lodging tolerance in direct-sown rice in flooded paddy field. Plant Prod. Sci. 6, 255–261 (2003).
Yoshinaga, S. Improved lodging resistance in rice (Oryza sativa L.) cultivated by submerged direct seeding using a newly developed hill seeder. JARQ 39, 147–152 (2005).
Acknowledgements
We thank the staff of the technical support section of NICS and NIAS for technical assistance with field experiment management and evaluation. This research was supported by Grants-in-Aid from NARO and two grants from the Ministry of Agriculture, Forestry and Fisheries of Japan (QTL-4003 and RBS-2009).
Author information
Authors and Affiliations
Contributions
Y.A.-S., Y.U. and T.T. designed the experiments and wrote the manuscript. Y.U. developed Dro1-NIL. Y.A.-S., Y.U. and T.T. performed yield, biomass and root trails. Y.A.-S. performed nitrogen and exudate trails. H.S. and M.K. performed cytokinin analysis. Y.A.-S. and Y.U. performed lodging resistance. Y.A.-S. and H.N. performed statistical analysis. M.K., Y.S. and H.N. provided advice on the experiments.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International License. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/4.0/
About this article

Cite this article
Arai-Sanoh, Y., Takai, T., Yoshinaga, S. et al. Deep rooting conferred by DEEPER ROOTING 1 enhances rice yield in paddy fields. Sci Rep 4, 5563 (2014). https://doi.org/10.1038/srep05563
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep05563
This article is cited by
-
Exploring root system architecture and anatomical variability in alfalfa (Medicago sativa L.) seedlings
BMC Plant Biology (2023)
-
Protein-protein interaction (PPI) network analysis reveals important hub proteins and sub-network modules for root development in rice (Oryza sativa)
Journal of Genetic Engineering and Biotechnology (2023)
-
Crop nitrogen (N) utilization mechanism and strategies to improve N use efficiency
Acta Physiologiae Plantarum (2023)
-
A comparative analysis of root traits in diverse wheat seedlings screened under optimum and low nitrogen stress in hydroponics
Plant Physiology Reports (2023)
-
Response of potato (Solanum tuberosum L.) cultivars to drought stress under in vitro and field conditions
Chemical and Biological Technologies in Agriculture (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
