
Abstract
Studies have shown that bone marrow-derived cells play an important role in tumor recurrence after chemotherapy and radiotherapy. In this study, we examined the relationship between the accumulation of Gr-1+CD11b+ cells and tumor recurrence after irradiation in tumor-bearing mice. By transplanting bone marrow cells into whole body-irradiated mice depleted of bone marrow, we assessed the role of Gr-1+CD11b+ cells in lung carcinoma models after local irradiation (LI). 20 Gy local irradiation could recruit CD11b+CXCR4+ cells into the irradiated tissues and the recruited CD11b+CXCR4+ cells could promote tumor recurrence. Further 6 Gy whole body irradiation (WBI6Gy) could decrease tumor recurrence by inhibiting the accumulation of Gr-1+CD11b+ cells and then suppressing tumor vasculogenesis and angiogenesis. Our results suggest that the accumulation of CD11b+Gr-1+ cells promote tumor re-growth after local irradiation by enhancing tumor neovascularization and low dose of whole body irradiation or irradiation of enlarged spleen may provide a new alternative for anti-angiogenesis therapies.
Similar content being viewed by others
Introduction
Epidemiological, clinical and experimental studies have demonstrated a link between chronic inflammation and cancer progression1,2,3. Tumor angiogenesis is a crucial step in tumor development since tumors have to establish a blood supply to grow and metastasize4,5. Mounting evidence has shown that tumor-infiltrating myeloid cells, such as tumor-associated macrophages (TAMs)6,7,8, Tie-2 expressing monocytes (TEMs)9,10, endothelial progenitor cells (EPCs)11,12,13, play a pivotal role in tumor angiogenensis. In some tumors, upon recruitment to tumors, these cells contribute to the revascularization by incorporating into the pre-existing vessels or secreting pro-angiogenic factors5,14. The two processes are referred to as vasculogenesis and angiogenesis, but their exact mechanisms have been poorly understood5.
Gr-1+CD11b+ myeloid cells of myeloid macrophage/DC (dendritic cells) lineage are significantly increased in spleen and bone marrow of tumor-bearing animals and elevated Gr-1+CD11b+ myeloid cells are associated with impaired immune reactivity and tumor progression15,16,17. Tumor tissues tend to express high levels of chemo-tactic factors such as CXCL12, CCL2 and stem cell factor (SCF), which recruit Gr-1+CD11b+ myeloid cells to the tumors2,18,19. The recruited cells are then involved in the tumor vasculature by secreting MMP9 (Matrix Metalloproteinases 9) or differentiating into endothelium-like cells or other vascular cells20,21,22. It has been well documented that Gr-1+CD11b+ myeloid cells play a crucial role in the regulation of tumor neo-vasculature and tumor recurrence23. TAMs and TEMs have been reported to play a key role in the tumor recurrence after irradiation24,25,26, but the underlying mechanism by which Gr-1+CD11b+ myeloid cells promote tumor recurrence after local irradiation (LI) remain unclear. In this study, we examined the roles of various Gr-1+CD11b+ myeloid cells in the post-LI re-growth in a lung tumor model.
Results
Accumulation of Gr-1+CD11b+ myeloid cells with the progression of Lewis lung carcinoma in tumor-bearing mice
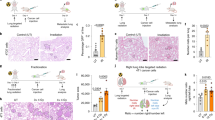
The presence of Gr-1+CD11b+ cells was flow cytometrically evaluated in the bone marrow, spleen and peripheral blood (Fig. 1A–C). First, we examined the Gr-1+CD11b+ cells in the bone marrow (Fig. 1A). It was found that two weeks after tumor inoculation, over 40% of the bone marrow cells expressed Gr-1+CD11b+ makers in LCC tumor-bearing mice, but only 21% of the bone marrow cells did so in normal mice. The proportion of Gr-1+CD11b+ cells was also significantly increased to over 15% in spleen and 20% in the peripheral blood in tumor-bearing mice, against a ratio of less than 5% in normal mice (Fig. 1B and C). More importantly, the weight of the spleen of tumor-bearing mice were 7 times more than that of the control mice 35 days after tumor inoculation(Fig. 1D and E). The inflammatory cells in the lung also increased with the accumulation of Gr-1+CD11b+ myeloid cells (Fig. 1F). These findings suggest that tumor progression affects the environment of the host, as manifested by the accumulation of Gr-1+CD11b+ cells and, especially, the infiltration of GR-1+CD11b+ cells into the spleen was a major contributor to this accumulation (Fig. 1E and F).

Accumulation of Gr-1+CD11b+cells in LCC tumor-bearing mice.
Flow cytometry analysis revealed that the proportion of Gr-1+CD11b+cells in the bone marrow (A) peripheral blood (B) and spleen (C) of tumor-bearing mice were increased with tumor progression. Increase in spleen weight followed the same pattern (D&E). HE staining showed that the inflammatory cells in the lung also increased with the accumulation of Gr-1+CD11b+ myeloid cells (F), Scale bar, 50 μm. n = 10 mice for each group. All the experiments were repeated three times.
Egress of CD11b+ myeloid cells to the peripheral blood and the recruitment of CD11b+ CXCR4+ myeloid cells to the tumor induced by local irradiation
In order to know whether local irradiation could induce the migration of Gr+CD11b+ cells into the tumors, in vivo imaging system was used to monitor the local infiltration of mononuclear cells from the bone marrow and spleen of tumor-bearing mice. The imaging showed that DIL-stained cells could infiltrate into the 20 Gy-irradiated tumors but not into the non-irradiated tumors (Fig. 2A and B). Some of the infiltrating DIL-stained cells expressed CD11b marker (Fig. 2A). Flow cytometry showed that 20 Gy LI could significantly promote the migration of CD11b+Gr-1+ cells into the peripheral blood one week after irradiation as compared with LLC-bearing mice without irradiation (Fig. 2C) (44.73% vs 26.53%, P < 0.001). The CD11b+Gr-1+ cells were also recruited to the irradiated tumor tissues, accounting for 9% of the total irradiated tumor cells against a less than 5% ratio in their non-irradiated counterparts (Fig. 2D). HE staining showed the area of tumor necrosis was significantly larger in 20 Gy LI-irradiated tumors (Fig. 3A and B) and the number of recruited CD11b+ cells adjacent to the necrotic tissues was also increased in LLC mice (Fig. 3A and C) 1 week after 20 Gy local irradiation. These results indicated that irradiation altered the microenvironment of the tumor and thereby recruited Gr-1+CD11b+ cells into the tumor tissues.

Recruitment of mononuclear cells from the bone marrow of tumor-bearing mice to the irradiated tumors.
DIL-stained mononuclear cells were recruited to the irradiated tumor (A), as shown by in vivo bioluminescent imaging (BLI) (B). The experiment was repeated three times. LI20Gy induced Gr-1+CD11b+cell migration into the peripheral blood (C) and the irradiated tumor tissues (D). Mice in A were drawn by T.L. n = 10 mice for each group.

Recruitment of the CD11b+ mononuclear cells to tumor necrotic tissues by local irradiation.
HE staining showed a marked increase in necrotic regions (N) (A&B) 1 week after 20 Gy local irradiation. Immunohistochemical staining demonstrated that he recruitment of CD11b+ cells in Lewis lung carcinomas was also enhanced (A&C).  : P < 0.01. n = 10 mice for each group. All the experiments were repeated three times.
: P < 0.01. n = 10 mice for each group. All the experiments were repeated three times.
20 Gy irradiated tumors were immunofluorescently examined for the co-expression of CD11b+CXCR4+ cells and results showed that number of cells expressing both CD11b+ and CXCR4+_were increased in the tumor tissues 7 days after irradiation (Fig. 4). Then AMD3100 was used to block the interaction between CXCR4 and SDF-1 to further confirm the role of CXCR4 in the recruitment of Gr-1+CD11b+ cells into the irradiated tumors34. Tumor-bearing mice were given AMD3100 two days before irradiation and the treatment lasted for 9 days. The infiltration of CD11b+CXCR4 cells was reduced upon AMD3100 treatment (Fig. 4B). It was found that AMD3100 treatment abolished the effect of LI on the recruitment of CD11b+ cells into the tumors, confirming the involvement of the CXCR4-expressing cells in the recruitment. Significantly, the infiltration of CD11b+CXCR4+ cells into the irradiated tumors was also decreased by WBI6Gy, which destroyed the bone marrow and depleted leukocytes temporarily (for 1.5–2 weeks) from the blood circulation (Fig. 4A and B). The finding suggested that WBI6Gy could effectively inhibit the recruitment of Gr-1+CD11b+ cells to the irradiated tumors. Transplantation of bone marrow from tumor-bearing mice into the WBI mice could restore the recruitment of Gr-1+CD11b+ cells in the WBI mice. Taken together, the above findings suggested that LI could increase the size of necrotic areas and the infiltration of CD11b+CXCR4+ cells into the microenvironment of the tumors and SDF -1α-CXCR4 axis plays an important part in the recruitment of CD11b+cells. Consistent with our results, other studies have shown that recurrent tumors following irradiation or tumors growing in an irradiated normal tissue were more hypoxic25,38 and therefore had elevated HIF-1 levels, which could induce the expression of SDF-1α and impact the SDF -1α-CXCR4 axis.

Decreased recruitment of bone marrow cells to the irradiated tumor tissues by whole body irradiation of 6 Gy (WBI6Gy) and AMD3100.
WBI6Gy and AMD3100 (an antagonist of CXCR4) could block the infiltration of CD11b+CXCR4+ cells after 20 Gy irradiation by immunofluorescent staining (A&B). Bone marrow reconstitution of the WBI mice with BM from tumor-bearing mice without irradiation could resume the infiltration of Gr-1+CD11b+ cell.  : P < 0.01; not significant (n.s.). n = 10 mice for each group. All the experiments were repeated three times.
: P < 0.01; not significant (n.s.). n = 10 mice for each group. All the experiments were repeated three times.
Delay of post-irradiation tumor recurrence by whole body irradiation via decreasing the accumulation of Gr-1+CD11b+ cells
In view of the fact that WBI significantly decreased the accumulation of Gr-1+CD11b+ cells, especially in the spleen, we further examined the effect of WBI on tumor growth. Tumor-bearing mice were treated with a sub-lethal dose of 6 Gy WBI and then the efficacy of the same radiation dose (20 Gy) administered either locally to tumors alone (LI20Gy) or LI14Gy in combination with 6 Gy WBI was compared. The growth of LCC tumors in C57BL/6 mice was initially arrested in both groups, while the tumor started to re-grow about 8 days later (Fig. 5A). It was noteworthy that the tumor growth was significantly delayed (12 days) in the mice treated with both LI and WBI as compared to LI alone (Fig. 5B). Moreover, WBI did not change the weight, appearance, or behavior of the mice (data not shown).

Effect of whole body irradiation of 6 Gy on the re-growth of LLC after local irradiation.
Whole body irradiation of 6 Gy could significantly delay the re-growth of LLC after local irradiation, while bone marrow transplantation one day after WBI6Gy could restore the re-growth (A&B). Effect of WBI6Gy on the size and weight of the spleen in tumor-bearing mice also was tested(C&D). Flow cytometry analysis revealed that the percentages of the Gr-1+CD11b+cells in the spleens remained unchanged (E) while the spleen weight was significantly reduced after WBI6Gy (C). The proportions of the Gr-1+CD11b+cells in the peripheral blood (F) and irradiate tumor tissues (G) were also decreased. Meanwhile, the transplantation of bone marrow cells could restore the spleen weight and accumulation of Gr-1+CD11b+cells.  : P < 0.01; not significant (n.s.). n = 10 mice for each group. All the experiments were repeated three times.
: P < 0.01; not significant (n.s.). n = 10 mice for each group. All the experiments were repeated three times.
We then examined if post-irradiation tumor recurrence was correlated with accumulation of Gr-1+CD11b+ cells in the spleen (Fig. 5C). LI reduced the spleen weight, but WBI6Gy decreased the spleen weight more significantly (Fig. 5D), although the proportions of CD11b+Gr-1+ cells didn't experience significant change (Fig. 5E). More importantly, flow cytometry exhibited that the percentage of Gr-1+CD11b+ cells in the peripheral blood and irradiated tumor tissues were also reduced significantly (Fig. 5F and G).
These results suggested that Gr-1+CD11b+ cells in the spleen might play an important role in the post-irradiation recurrence. One day after WBI6Gy, the irradiated mice were reconstituted with bone marrow cells from LLC-bearing mice without irradiation. BM reconstitution one day after LI plus WBI resumed the accumulation of CD11b+Gr-1+ cells in the spleen and infiltration of CD11b+Gr-1+cells into the irradiated tissues and thereby accelerated the tumor growth. We were led to conclude that WBI can significantly decrease the accumulation of CD11b+Gr-1+cells in the spleen, reduce the infiltration of Gr1+CD11b+ cells in the tumor and finally delay the tumor re-growth after irradiation.
Elimination of CD11b+ myelomonocytic cells-derived neovasculature by WBI6Gy
MVD in the harvested tumors of different groups was examined. LI14Gy plus WBI6Gy significantly decreased MVD counts (P < 0.05) in the Lewis-lung carcinoma models as compared with LI20Gy alone (Fig. 6E). Moreover, bone marrow transplantation (with the proportion of CD11b+Gr-1+ cells from LLC-bearing mice being over 50%) one day after WBI6Gy abrogated the decrease and restored the CD31-expressing neo-vasculature (Fig. 6E). Monocytes/macrophages have been known to promote tumor neo-vasculature by direct incorporation into tumor endothelia or by paracrine secretion of pro-angiogenesis factors such as MMP-9, iNOS. One week after LI20Gy alone, frozen tumor sections were examined for co-expression of CD11b with CD31, α-SMA, MMP-9 and iNOS. Histological examination showed that the proportions of vessels co-expressing CD11b+CD31+ or CD11b+α-SMA+ to total CD31- or α-SMA-expressing vessels were 14.31% ± 2.43% and 12.50% ± 4.12% (Fig. 6A and C). On the other hand, the proportions of cells co-expressing CD11b+MMP-9+ or CD11b+ iNOS+ to MMP-9- or iNOS-expressing cells were 65.44% ± 11.18% and 93.50% ± 4.12% (Fig. 6B and D). These findings showed that pro-angiogenesis factors may play a more important role in neovasculature and irradiated tumors were able to recruit CD11b+ cells, which were involved in neovasculature more through an angiogeneic way than vasculogenenic way.

The role of CD11b+ BMDCs in the angiogenesis and vasculogenesis processes of Lewis tumors one week after 20 Gy local irradiation.
Double immunostaining of CD11b with CD31, a-SMA, MMP-9 and iNOS for tumor sections were used to demonstrate the role of CD11b+ BMDCs in tumor neovasculature. Quantification of CD31-positive vessels per HPF in the tumor sections was used to identify tumor microvessel density (E). Immunostaining revealed the vessels co-expressing CD11b+CD31+/α-SMA+ in tumor sections (A).Immunostaining showed the proportions of vessels co-expressing CD11b+CD31+ or CD11b+α-SMA+ to total CD31- or α-SMA-expressing vessels in tumors sections(C). Immunostaining revealed the cells co-expressing CD11b+MMP-9+/iNOS+ in tumors sections (B). Immunostaining showed the proportions of cells co-expressing CD11b+MMP-9+ or CD11b+ iNOS+ to MMP-9- or iNOS-expressing cells in tumors sections (D). Not significant (n.s.). n = 10 mice for each group. All the experiments were repeated three times.
Role of iNOS in CD11b+ myelomonocytic cells-derived neovasculature
iNOS expression in irradiated tumor tissues were then examined. IHC confirmed that LI20Gy increased the iNOS expression in the irradiated tumors while WBI6Gy abrogated the increase of iNOS expression (Fig. 7A and B). However, bone marrow transplantation one day after WBI6Gy could restore the expression of iNOS in locally-irradiated tumor tissues. Then, tumor tissues were flow cytometrically separated into tumor cells and stromal cells. Moreover, the NO immunofluorescence density (DAF-FM DA probe) were found to be significantly higher in the stromal tissues than in the tumor tissues (P < 0.05) (Fig. 7C–G). These data suggested that after irradiation iNOS from stromal cells was the major pool for NO and the infiltrating CD11b+ cells highly expressing iNOS had the potential to mediate tumor recurrence by enhancing tumor neovasculature after local irradiation.

iNOS expression in the Lewis tumor tissues.
IHC staining showed that iNOS-stained cells accumulated in the mesenchymal tissues and necrotic areas (A). IHC staining revealed that the Lewis tumor cells expressed iNOS protein after irradiation (B). In situ flow cytometrical determination of the nitric oxide in tumors also was done. The tumor tissues were separated into mesenchymal cells (P1) and parenchymal cells (P2) (C&D). Flow cytometric analysis showed the percentage of DAF-FM DA+ cells of the total cells in P1 and P2 gates (E&F). Fluorescence density of DAF-FM DA probes in tumor tissues (G).  : P < 0.001;
: P < 0.001;  : P < 0.01; not significant (n.s.). n = 10 mice for each group. All the experiments were repeated three times.
: P < 0.01; not significant (n.s.). n = 10 mice for each group. All the experiments were repeated three times.
Discussion
Tumor vasculature is derived from sprouting of local vessels (angiogenesis) and bone marrow (BM)-derived circulating cells (vasculogenesis)14. Owing to the tumor-bed effect37,38, tumor angiogenesis is abrogated due to the destruction of the pre-existing blood vessels after irradiation. Several studies have demonstrated that CD11b+ bone marrow-derived cells, such as TAMs and TEMs, play a crucial role in promoting tumor recurrence after irradiation by promoting vasculogenesis rather than angiogenesis24,25. TEMs, TAMs and MDSCs are all subpopulations of the total tumor infiltrating CD11b+ myeloid cells and they could be distinguished from each other by their specific surface marker profile14. Therefore, Gr-1+CD11b+ myeloid cells, which were also of bone marrow origin, were believed to promote tumor neovasculature after irradiation22,23,24,25 but the detailed mechanisms were not fully understood.
CD11b + Gr-1 + myeloid cells can expand systemically in bone marrow, spleen and peripheral blood20,21. In this study, by using the Lewis lung carcinoma model, we demonstrated that accumulation of Gr-1+CD11b+ myeloid cells in tumor-bearing mouse, especially in spleen, were associated with the progression of tumors. More importantly, we also revealed that Gr-1+CD11b+ cells could promote tumor recurrence by increasing MVD of irradiated LLC. An array of chemokines, growth factors and adhesion molecules are required to work in concert with bone marrow cells to be mobilized to the peripheral bloods and recruited to tumor tissues. In particular, CXCR4 plays an important part in the recruitment of inflammatory cells to the irradiated tumor tissues33. SDF-1/CXCR4 axis is also implicated in the recruitment of hematopoietic stem cells and progenitor cells, supporting our observation that Gr-1+CD11b+ cells had differentiation potential and were progenitor-like EPCs18,19. Our data also showed that infiltration of Gr-1+CD11b+ cells into the irradiated tumor tissues could be significantly reduced by using the CXCR4 antagonist AMD3100 and 6 Gy whole body irradiation. More importantly, 6 Gy whole body irradiation could significantly decrease the accumulation of Gr-1+CD11b+ cells in an enlarged spleen, which have been demonstrated to be associated with tumor progression. Finally, we showed that 6 Gy whole body irradiation could delay the recurrence of Lewis lung carcinoma after local irradiation by reducing the accumulation of Gr-1+CD11b+ cells.
We exhibited that CD11b+ myeloid cells in the irradiated tumors could functionally mimic CD31-expressiong endothelial cells and α-SMA-expressing mural cells and thereby increased the microvessel density and, in the same way, promoted tumor recurrence. These results indicated that Gr-1+CD11b+ cells could enhance vasculogenesis to promote MVD in the irradiated tumors and might play an important role in the tumor recurrence after irradiation. Different studies yielded different results concerning the roles of bone marrow-derived cells in tumor neo-vasculature and the discrepancies might partly be due to different models used5,14. Our data also showed that Gr-1+CD11b+ myeloid cells were mostly the bone-marrow cells and spleen cells in tumor bearing mice (Fig. 1A), suggesting that Gr-1+CD11b+ myeloid cells may play a more important role than other CD11b+cells in tumor neo-vasculature. This speculation warrants further study.
Some studies reported that myeloid inflammatory cells, after being recruited to tumor tissues, could directly incorporate into the existing blood vessels and/or secrete pro-angiogenetic factors, such as MMP-9 and iNOS14,20,22, to promote tumor recurrence after irradiation. CD11b+ cells could express high level of matrix metalloproteinase-9 (MMP-9), which has been found to play a crucial role in tumor vasculogenesis in a pre-irradiated model22 and promote tumor angiogenesis39,40. CD11b-positive myelomonocytic cells from the transplanted BM were responsible for tumor growth and the development of immature blood vessels in MMP-9 knockout mice receiving wild-type BM22. This suggests that MMP-9 also plays an important role in tumor neo-vasculature after irradiation. More interestingly, our data showed that more than 95% infiltrated CD11b+cells expressed iNOS, partly owing to the high impression of NF-κβ after irradiation. As a signaling molecule, NO (Nitric oxide) regulates various vascular functions, including angiogenesis, blood flow, vascular permeability and micro-lymphatic flow35. The elevated expression of iNOS (inducible nitric oxide synthase) in infiltrating CD11b+cells after irradiation showed that iNOS might play an important role in the revasculation36. Various studies also showed that NO could both promote and inhibit tumor progression and metastasis41,42,43. The effects of NO in tumors seem to depend on the activity and localization of NOS isoforms, concentration, duration of NO exposure and cellular sensitivity to NO43. The effect of altered NO levels on tumor growth and angiogenesis has also been well studied by pharmacological manipulation with either NO donors or NOS inhibitors. Treatment of mice bearing mammary and squamous carcinomas with NG-nitro-L-arginine methyl ester, a universal NOS inhibitor, as well as specific inhibition of iNOS by either 1400 W or L-nil, reduced subcutaneous tumor growth and vessel development44,45,46. A single dose of the NOS inhibitor NG-nitro-L-arginine (L-NNA) was shown to induce an acute and sustained reduction in human tumor blood volume, providing clinical evidence that inhibition of NOS has anti-tumor vascular effects47. The importance of host-derived NO for tumor growth has also been genetically investigated in iNOS knockout mice, in which tumor growth and vascularization were decreased48. Although tumor cells are also a major source of NO, which can affect angiogenesis and modulate tumor progression, our data showed that, after irradiation, most of the NO came from the mesenchymal cells.
To sum up, our results showed that Gr-1+CD11b+ cells were associated with tumor progression and could promote tumor recurrence after irradiation by enhancing tumor neovascularization. Moreover, recruited CD11b+cells highly expressing iNOS may play an angiogenic role in tumor neovascularization. Then whole body irradiation of 6 Gy was proven to be an effective way to delay the recurrence in LLC-bearing mice after local irradiation by depleting the accumulation of Gr-1+CD11b+ cells, especially in the spleen. More interestingly, irradiation of enlarged spleen might provide a novel alternative of anti-angiogenesis therapies.
Methods
Mice
Male C57BL/6 mice, aged 8–10 weeks, were purchased from Beijing University (Beijing, China). The mice were maintained on a chow diet on a 12-hour light/12-hour dark cycle at 25°C in the Animal Care Facility of Tongji Medical College by strictly following the institutional guidelines.
Cell culture
Lewis lung carcinoma (LLC) cells were obtained from the American Type Tissue Collection (Manassas, Virginia, USA) and maintained by employing standard cell culture techniques. LLC cells were cultured in RPMI 1640 supplemented with 10% FBS (Gibco, Grand Island, NY).
Tumor models
Lewis lung caner cells (5 × 105) were injected subcutaneously (s.c.) onto the back of the mouse approximately 1 cm proximal to the base of the tail. Tumors were measured with a caliper and tumor volumes (mm3) were calculated by the following formula: Volume = length × width2 × π/6.
Treatment of animals
Treatments were initiated when the mean diameter of tumor reach 0.75 cm. The tumor-bearing mice were given 20 Gy local irradiation (LI) or 14 Gy local irradiation in combination with 6 Gy sublethal whole body irradiation (WBI) to suppress the bone marrow and temporarily deplete leukocytes24,28. Some mice were injected AMD3100 (Sigma-Aldrich, at 5 mg/kg i.p., twice a day), a specific CXCR4 inhibitor. On the third day, mice were administered 20 Gy LI 2 hours after the first dose of AMD-3100. AMD3100 was injected until the mice were sacrificed26. One day after the WBI6Gy, bone marrow was harvested from the femurs of the tumor-bearing mice. Then cells (1 × 107) were injected i.v. through the tail vein of WBI-irradiated recipient mice. Therapeutic efficacy was measured by the time taken for tumors to grow from 7.5 mm to 10 mm in diameter.
Flow cytometry
Single-cell suspension was prepared from resected tumors which were lysed with 1% collagenase (Sigma-Aldrich). Cells from spleens, bone marrows and peripheral blood were collected by mechanical disruption. Red blood cells were removed by hypotonic lysis. Cells were then washed and stained for 30 minutes on ice with antibodies in PBS with 1% inactivated fetal calf serum by following instructions. The antibodies, CD11b(130-081-201)29, Gr-1(130-091-931)30, used for flow cytometry were procured from Miltenyi Biotec.
Cells tracking
Mononuclear cells (BM-MNCs) were obtained by flushing tibia and spleen from the LLC-bearing mice. Briefly, cells were labeled with the lipophilic membrane dye Dil (Invitrogen, Carlsbad, CA) according to the manufacturer's protocol at 37°C for 1 h. The cells were then collected and observed under a confocal microscope for labeling rate. Bioluminescent imaging (BLI) was performed by utilizing the IVIS Imaging System (Xenogen, Baltimore, MD) 3 and 7 days after the Dil-Staining cells (1 × 106 cells) were injected via the tail veins.
Western blot analysis
For Western blotting, tumor tissue lysates were loaded onto 10% SDS-PAGE, transferred to nitrocellulose membrane and then incubated for immunoblotting by using primary antibodies. Secondary antibodies conjugated to alkaline phosphatase were purchased from Jackson ImmunoResearch Laboratories Inc. and used at 1:10,000 dilutions. The primary antibodies against NF-kB p65 (ab31481, Abcam) or iNOS (ab15323, Abcam) were purchased from Abcam. β-actin was from Santa Cruz Biotechnology Inc.
Immunohistochemical and immunofluorescent staining
For necrosis assay, LLC tumor sections were HE-stained and histopathologically examined to identify necrotic areas in tumor tissues. CD11b+ cells accumulation was immunohistochemically assessed in formalin-fixed, paraffin-embedded 4 μm thick tumor serial sections by using a rabbit anti-mouse CD11b antibody (Bioss, Beijing). Frozen tissue sections were co-immunostained with rat anti-mouse CD11b-FITC (ab24874, Abcam) and rabbit anti-CD31 (SAB1302548, sigma), CXCR4 (ab2074, Abcam), Cy3-conjugated mouse anti-α-SMA (1:400; Sigma) antibodies, anti-iNOS (ab15323, Abcam) or anti-MMP-9 (Bioss, Beijing). Then staining with anti-CD 31 antibody, anti-CXCR4 antibody, anti-iNOS antibody and anti-MMP-9 antibody was followed by staining with a DyLight 488-conjugated goat anti-rabbit secondary antibody (1:200; Jackson ImmunoResearch).
Microvessel density
To evaluate tumor angiogenesis, microvessel density (MVD) was determined by immunohistochemical staining of CD3131. The generally accepted criteria for determining a vessel profile32 were used. Any stained endothelial cell or endothelial cell cluster that was separate from adjacent microvessels was counted as a microvessel. Vessel lumens were not essential for the identification of a microvessel.
Histological assessment
For CD11b+ cells in tumors, only DAPI-positive (viable tissues) areas were subjected to confocal microscopy22,24. The ratios of MMP-9+, iNOS +, CD31+, α-SMA+ cells to CD11b+ cells in the selected fields were calculated as percentage of CD11b-positive area. Images were captured using a LMS510 confocal microscope (ZEISS, Germary) and were analyzed by using Image-Pro Plus 6.0 software package (Media Cybernetics, MD). For each tumor, five fields were randomly selected from immunohistochemically or immunofluorescently stained slices, and, with each group, 5 to 6 tumors were studied.
Flow cytometrical analysis of NO
Single-cell suspension was made from resected tumors. The cells were incubated with 5 μM fluorescent probe DAF-FM DA (3-amino, 4-aminomethyl-2′, 7′-difluorescein, diacetate; Beyotime, China) in the dark at 37°C for 30 minutes. The cells were rinsed with HEPES-NaOH (pH 7.5) three times (2 min each time) to remove excess fluorescence and further incubated in the dark in probe-free medium (30 min, 37°C) before flow cytometrical analysis (FACSCalibur, BD).
Ethics statement
All experiments were approved by the local ethics committee for animal welfare (Animal Care and Utilization Committee of Huazhong University of Science and Technology, Wuhan, China) and complied with the guidelines for the welfare of animals for experimental cancer research27. All surgeries and irradiation procedures were performed under sodium pentobarbital anesthesia and efforts were made to minimize the animal suffering.
Statistical analysis
All data were expressed as mean ± SEM. Differences among groups were evaluated by ANOVA or the unpaired Student's t test by using GraphPad Prism5 software. A P value <0.05 was considered to be statistically significant.
References
Grivennikov, S. I., Greten, F. R. & Karin, M. Immunity, inflammation and cancer. Cell 140, 883–899 (2010).
Ostrand-Rosenberg, S. & Sinha, P. Myeloid-derived suppressor cells: linking inflammation and cancer. J Immunol 182, 4499–4506(2009).
Tennyson, A. G. & Lippard, S. J. Generation, translocation and action of nitric oxide in living systems. Chem Biol 18, 1211–1220 (2011).
Folkman, J. Tumor angiogenesis: therapeutic implications. N Engl J Med 285, 1182–1186 (1971).
Kerbel, R. S. Tumor angiogenesis. N Engl J Med 358, 2039–2049 (2008).
Allavena, P., Sica, A., Solinas, G., Porta, C. & Mantovani, A. The inflammatory micro-environment in tumor progression: the role of tumor-associated macrophages. Crit Rev Oncol Hematol 66, 1–9 (2008).
Coffelt, S. B., Hughes, R. & Lewis, C. E. Tumor-associated macrophages: effectors of angiogenesis and tumor progression. Biochim Biophys Acta 1796, 11–18 (2009).
Solinas, G., Germano, G., Mantovani, A. & Allavena, P. Tumor-associated macrophages (TAMs) as major players of the cancer-related inflammation. J Leukoc Biol 86, 1065–1073(2009).
Venneri, M. A. et al. Identification of proangiogenic TIE2-expressing monocytes (TEMs) in human peripheral blood and cancer. Blood 109, 5276–5285(2007).
De Palma, M. et al. Tie2 identifies a hematopoietic lineage of proangiogenic monocytes required for tumor vessel formation and a mesenchymal population of pericyte progenitors. Cancer Cell 8, 211–226(2005).
Vajkoczy, P. et al. Multistep nature of microvascular recruitment of ex vivo-expanded embryonic endothelial progenitor cells during tumor angiogenesis. J Exp Med 197, 1755–1765(2003).
Orimo, A. et al. Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion. Cell 121, 335–348(2005).
Duda, D. G. et al. Evidence for incorporation of bone marrow-derived endothelial cells into perfused blood vessels in tumors. Blood 107, 2774–2776 (2006).
Murdoch, C., Muthana, M., Coffelt, S. B. & Lewis, C. E. The role of myeloid cells in the promotion of tumour angiogenesis. Nat Rev Cancer 8, 618–631 (2008).
Marx, J. Cancer immunology. Cancer's bulwark against immune attack: MDS cells. Science 319, 154–156 (2008).
Yan, H. H. et al. Gr-1+CD11b+ myeloid cells tip the balance of immune protection to tumor promotion in the premetastatic lung. Cancer Res 70, 6139–6149 (2010).
Sinha, P. & Clements, V. K. Ostrand-Rosenberg S. Reduction of myeloid-derived suppressor cells and induction of M1 macrophages facilitate the rejection of established metastatic disease. J Immunol 174, 636–645 (2005).
Duda, D. G. et al. CXCL12 (SDF1alpha)-CXCR4/CXCR7 pathway inhibition: an emerging sensitizer for anticancer therapies? Clin Cancer Res 17, 2074–2080 (2011).
Petit, I., Jin, D. & Rafii, S. The SDF-1-CXCR4 signaling pathway: a molecular hub modulating neo-angiogenesis. Trends Immunol 28, 299–307 (2007).
Yang, L. et al. Expansion of myeloid immune suppressor Gr+CD11b+ cells in tumor-bearing host directly promotes tumor angiogenesis. Cancer Cell 6, 409–421 (2004).
Suzuki, E., Kapoor, V., Jassar, A. S., Kaiser, L. R. & Albelda, S. M. Gemcitabine selectively eliminates splenic GR-1+/CD11b+ myeloid suppressor cells in tumor-bearing animals and enhances antitumor immune activity. Clin Cancer Res 11, 6713–6721 (2005).
Ahn, G. O. & Brown, J. M. Matrix metalloproteinase-9 is required for tumor vasculogenesis but not for angiogenesis: role of bone marrow-derived myelomonocytic cells. Cancer Cell 13, 193–205 (2008).
Xu, J. et al. CSF1R Signaling Blockade Stanches Tumor-Infiltrating Myeloid Cells and Improves the Efficacy of Radiotherapy in Prostate Cancer. Cancer Res 73, 2782–2794(2013).
Kozin, S. V. et al. Recruitment of myeloid but not endothelial precursor cells facilitates tumor regrowth after local irradiation. Cancer Res 70, 5679–5685 (2010).
Kioi, M. et al. Inhibition of vasculogenesis, but not angiogenesis, prevents the recurrence of glioblastoma after irradiation in mice. J Clin Invest 120, 694–705(2010).
Welford, A. F. et al. TIE2-expressing macrophages limit the therapeutic efficacy of the vascular-disrupting agent combretastatin A4 phosphate in mice. J Clin Invest 121, 1969–1973 (2011).
Workman, P. et al. Guidelines for the welfare and use of animals in cancer research. Br J Cancer 102, 1555–1577(2010).
Seung, L. P., Weichselbaum, R. R., Toledano, A., Schreiber, K. & Schreiber, H. Radiation can inhibit tumor growth indirectly while depleting circulating leukocytes. Radiat Res 146, 612–618 (1996).
Li, H. et al. Different neurotropic pathogens elicit neurotoxic CCR9-or neurosupportive CXCR3-expressing microglia. The Journal of Immunology 177, 3644–3656 (2006).
Fleming, T., Fleming, M. & Malek, T. Selective expression of Ly-6G on myeloid lineage cells in mouse bone marrow. RB6-8C5 mAb to granulocyte-differentiation antigen (Gr-1) detects members of the Ly-6 family. J Immunol 151, 2399–2408 (1993).
Weidner, N. Current pathologic methods for measuring intratumoral microvessel density within breast carcinoma and other solid tumors. Breast Cancer Res Treat 36, 169–180(1995).
Weidner, N. et al. Tumor angiogenesis: a new significant and independent prognostic indicator in early-stage breast carcinoma. J Natl Cancer Inst 84, 1875–1887(1992).
Tseng, D., Vasquez-Medrano, D. & Brown, J. Targeting SDF-1/CXCR4 to inhibit tumour vasculature for treatment of glioblastomas. Br J Cancer 104, 1805–1809(2011).
Hatse, S., Princen, K., Bridger, G., De Clercq, E. & Schols, D. Chemokine receptor inhibition by AMD3100 is strictly confined to CXCR4. FEBS Lett 527, 255–262(2002).
Singh, S. & Gupta, A. K. Nitric oxide: role in tumour biology and iNOS/NO-based anticancer therapies. Cancer Chemother Pharmacol 67, 1211–1224 (2011).
Cianchi, F. et al. Heterogeneous expression of cyclooxygenase-2 and inducible nitric oxide synthase within colorectal tumors: correlation with tumor angiogenesis. Dig Liver Dis 42, 20–27 (2010).
Rofstad, E. K., Mathiesen, B., Henriksen, K., Kindem, K. & Galappathi, K. The tumor bed effect: increased metastatic dissemination from hypoxia-induced up-regulation of metastasis-promoting gene products. Cancer Res 65, 2387–2396 (2005).
Zips, D. et al. Impact of the tumour bed effect on microenvironment, radiobiological hypoxia and the outcome of fractionated radiotherapy of human FaDu squamous-cell carcinoma growing in the nude mouse. Int J Radiat Biol 77, 1185–1193(2001).
Kong, D., Li, Y., Wang, Z., Banerjee, S. & Sarkar, F. H. Inhibition of angiogenesis and invasion by 3,3′-diindolylmethane is mediated by the nuclear factor-kappaB downstream target genes MMP-9 and uPA that regulated bioavailability of vascular endothelial growth factor in prostate cancer. Cancer Res 67, 3310–3319 (2007).
Pozzi, A., LeVine, W. F. & Gardner, H. A. Low plasma levels of matrix metalloproteinase 9 permit increased tumor angiogenesis. Oncogene 21, 272–281 (2002).
Kostourou, V. et al. The role of tumour-derived iNOS in tumour progression and angiogenesis. Br J Cancer 104, 83–90(2011).
Cullis, E. R. et al. Tumour overexpression of inducible nitric oxide synthase (iNOS) increases angiogenesis and may modulate the anti-tumour effects of the vascular disrupting agent ZD6126. Microvasc Res 71, 76–84(2006).
Fukumura, D., Kashiwagi, S. & Jain, R. K. The role of nitric oxide in tumour progression. Nat Rev Cancer 6, 521–534 (2006).
Jadeski, L. C. & Lala, P. K. Nitric oxide synthase inhibition by N(G)-nitro-L-arginine methyl ester inhibits tumor-induced angiogenesis in mammary tumors. Am J Pathol 155, 1381–1390 (1999).
Thomsen, L. L. et al. Selective inhibition of inducible nitric oxide synthase inhibits tumor growth in vivo: studies with 1400 W, a novel inhibitor. Cancer Res 57, 3300–3304(1997).
Sikora, A. G. et al. Targeted inhibition of inducible nitric oxide synthase inhibits growth of human melanoma in vivo and synergizes with chemotherapy. Clin Cancer Res 16, 1834–1844(2010).
Ng, Q. S. et al. Effect of nitric-oxide synthesis on tumour blood volume and vascular activity: a phase I study. Lancet Oncol 8, 111–118 (2007).
Konopka, T. E. et al. Nitric oxide synthase II gene disruption: implications for tumor growth and vascular endothelial growth factor production. Cancer Res 61, 3182–3187(2001).
Acknowledgements
This work was supported by the Hubei Provincial Natural Science Foundation. (Project No2009CDA061).
Author information
Authors and Affiliations
Contributions
G.W. and T.L. designed the experiments. T.L. and C.X.Y. performed the experiments and analysed the data. S.Z., Y.C.L., L.L.S. and Y.M.F. help in the experiments. T.L., C.Y.X. and G.W. co-wrote the manuscript. D.D.Y., T.Z. and H.M. help in data collection, data analysis and manuscript writing. All authors discussed the results and commented on the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 3.0 Unported License. The images in this article are included in the article's Creative Commons license, unless indicated otherwise in the image credit; if the image is not included under the Creative Commons license, users will need to obtain permission from the license holder in order to reproduce the image. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article

Cite this article
Liu, T., Xie, C., Ma, H. et al. Gr-1+CD11b+ cells facilitate Lewis lung cancer recurrence by enhancing neovasculature after local irradiation. Sci Rep 4, 4833 (2014). https://doi.org/10.1038/srep04833
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep04833
This article is cited by
-
BMP9 signaling promotes the normalization of tumor blood vessels
Oncogene (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
