
Abstract
Toll-like receptors (TLRs) are innate receptors critical for host defense and play a role in normal biological processes. For example, host DNA, a TLR9 ligand, stimulates epithelial repair following skin wounding. TLR signaling also plays a crucial role in regulating intestinal homeostasis. We therefore asked whether TLR9 is important for intestinal wound repair using a dextran sulfate sodium (DSS)-induced intestinal damage and repair model. We showed that TLR9-deficient mice are more susceptible to DSS and exhibited delayed wound repair at both the clinical and histologic levels. TLR9-deficient mice showed reduced gene expression of hairy enhancer of split 1, an intestinal progenitor cell differentiation factor and vascular endothelial growth factor, a growth factor important for epithelial cell restitution. Therefore, we conclude that TLR stimulation may play a normal role in regulating intestinal homeostasis and could potentially be a novel therapeutic target to enhance intestinal wound repair in inflammatory bowel diseases.
Similar content being viewed by others

Introduction
The intestinal mucosa is composed of a simple columnar epithelium (enterocytes), covering the crypt/villus units and crypt units of the small intestine and colon, respectively1,2,3. This mucosal surface is continuously exposed to a multitude of factors derived from dietary4,5,6, foreign6,7,8,9,10 and self sources6,11 that can result in disruption of the physical barrier7,9. Therefore, the intestinal mucosa must perform its physiologic function of nutrient absorption2 while maintaining the ability to sense and respond to intestinal injury6,7,8,9,10,12. The enterocytes that comprise the intestinal mucosa express several different families of innate immune receptors that recognize dietary, foreign and self antigens6,7,8,9,10,12. Microbial antigen recognition is important because the intestinal mucosa is colonized with approximately 1014 commensal bacteria13. These commensal bacteria are a vital component of intestinal development14,15, physiology16,17 and immunology7,18. A complex, dynamic balance, or homeostasis, is continuously maintained between the commensal bacteria and the immune response of the intestinal mucosa7,19,20. Disruption of homeostasis, termed dysbiosis, can result in several types of inflammatory bowel diseases (IBD), such as Crohn's disease and ulcerative colitis19,20.
Crohn's disease and ulcerative colitis are characterized by recurrent episodes of autoimmune activity, resulting in ulceration of the intestinal mucosa and remission, with no or subclinical disease21,22. Crohn's disease primarily affects the small intestine and colon, while ulcerative colitis affects only the colon21,22. Recent work showed that development of IBD requires multiple components, consisting of genetic10,23,24, environmental24,25, microbial26,27 and immunological factors28,29. One important component involved in the recognition of commensal bacteria is a family of ten innate immune receptors called Toll-like receptor (TLR) 1–1030. TLRs recognize conserved molecular patterns of commensals or self resulting in pro-inflammatory responses and activation of immune cells via induction of specific cellular signaling pathways30. Rakoff-Nahoum et al. showed that myeloid differentiation factor 88 (MyD88), an adaptor molecule used for signaling by all TLRs except TLR330, was required for commensal bacteria recognition and initiation of colonic enterocyte proliferation following radiation or dextran sulfate sodium (DSS)-induced injury7, a chemically-induced model for an ulcerative colitis-like disease state in mice31,32,33. MyD88-deficient mice exhibited increased susceptibility to DSS and reduced colonic enterocyte proliferation following induction of colitis7. These data provided the first indication that TLR signaling might be involved in the development of IBD.
Specific TLR-deficient mice were utilized to evaluate the potential role of TLRs in the development of intestinal injury using the DSS colitis model7,8,9,10,34,35. Cario et al. showed that DSS treated, TLR2-deficient mice were more susceptible to injury due to increased tight junction permeability, allowing for increased commensal bacteria translocation across the intestinal mucosa and induction of severe inflammatory responses9. Similar to the MyD88-deficient mice7, TLR4-deficient mice were more susceptible to DSS-induced colitis and had less colonic enterocyte proliferation8. Interestingly, TLR5-deficient mice developed spontaneous colitis in the absence of DSS treatment, likely due to increased bacterial burdens and reduced anti-inflammatory cytokine secretion10,36. Data on the nucleic acid-sensing TLRs in the intestinal mucosa is limited and two published studies show differing results for the role of TLR934,35. While the role of TLRs in the development of intestinal injury has been characterized7,8,9,10,34,35, the contribution of TLRs to the intestinal wound repair process in vivo is still not well understood.
Intestinal progenitor cells, which are present in the intestinal crypts, continually proliferate and differentiate into enterocytes, goblet cells, or enteroendocrine cells to replace the mature cells that are lost due to rapid turnover (3–5 days) or injury1,2,3. Differentiation into enterocytes is controlled by expression of hairy enhancer of split 1 (HES1) and this transcription factor controls expression of ATOH1 (MATH1 in mice), which induces differentiation into goblet or enteroendocrine cells1,2,3. Following intestinal injury, progenitor cell proliferation and differentiation into mature epithelial cells increased expression of other factors involved in initiating the wound repair process, such as vascular endothelial growth factor (VEGF) and pro-inflammatory cytokines6,37. Recent research using murine skin wounding models has implicated TLR9 in the wound healing process38,39,40. Recognition of self DNA, via TLR9 expressed on plasmacytoid dendritic cells, induced a rapid pro-inflammatory cytokine response, consisting of type I interferons, interleukin-6 and tumor necrosis factor-alpha, which was important for initiation of the wound healing process38,39. Additionally, Sato et al. showed that application of CpG DNA, a TLR9 agonist30, accelerated wound healing in a murine skin biopsy-wounding model40. Interestingly, TLR9-deficient mice treated with or without CpG showed a delay in the wound healing process starting at 4 days post- wounding40.
Based on the role for TLR9 recognition of self DNA and induction of epithelial restitution following skin wounding38,39,40, we hypothesized that TLR9 promotes intestinal wound repair. Herein, we show that TLR9-deficient mice were more susceptible to DSS-induced colonic injury as measured by clinical and histologic parameters and exhibited delayed wound repair. TLR9-deficient mice had reduced expression of HES1 and VEGF. Our data support a model wherein TLR9 is important for protecting the intestinal epithelium from damage and for promoting wound repair. Understanding the mechanisms of intestinal injury and repair will provide key insights into normal host biology and provide novel intervention points that could be utilized to prevent or treat IBD.
Results
TLR9-deficient mice have increased susceptibility to colonic injury
MyD88-deficient mice have defects in intestinal homeostasis and repair7, TLR9 signaling requires MyD8830 and TLR9 is important for restitution following skin wounding38,39,40. We therefore asked whether TLR9 promotes intestinal wound repair using a mouse model of DSS-induced intestinal damage31,32,33. We administered a concentration of DSS that caused intestinal damage, but not so severe that wild-type mice could not repair the damage rapidly after removal of the DSS treatment. Wild-type and TLR9-deficient mice were treated with 1% DSS in the drinking water for seven days and monitored for disease. During the acute phase of DSS-induced intestinal damage, only a few wild-type mice tested positive for blood in the stool on the last day of treatment (Figure 1A). Two wild-type mice developed changes in stool consistency by day 6 post-DSS treatment (Figure 1B). However, under these mild treatment conditions, wild-type mice did not lose weight (Figure 1C). This is in contrast to the earlier and more severe disease observed in TLR9-deficient mice. TLR9-deficient mice showed significantly (p < 0.05; Mann Whitney test) increased occult blood scores, which developed earlier (Figure 1D). Furthermore, TLR9-deficient mice showed significantly (p < 0.001; Mann Whitney test) increased changes in stool consistency as early as day 2 post-DSS treatment, with more mice affected than the wild-type cohort (Figure 1E). Additionally, TLR9-deficient mice showed significant (p < 0.01; Student's t-test) weight loss over the course of treatment (Figure 1C). These data support previous studies showing that TLR9 is involved in protection against initial intestinal injury35.

TLR9-deficient mice showed increased clinical parameter severity following 1% DSS treatment.
Wild-type (A, B) and TLR9-deficient (D, E) mice were given normal water (untreated) or 1% DSS in the drinking water for seven days, then switched to regular drinking water. Fecal samples were screened daily for occult blood, by Hemoccult® test (A, D) and consistency (B, E), as described in materials and methods. (C) Mice were weighed daily to determine the percent weight change during the treatment (Day 0–7) and recovery (Day 7–14) phases. (F) Colon length was measured at day 14 for 1% DSS- treated wild-type and TLR9-deficient mice. n = 6 mice/group for each genotype, two independent experiments = 12 mice per group analyzed. *p < 0.05, **p < 0.01, Student's t-test.
To examine intestinal repair processes, mice were returned to regular drinking water and monitored for an additional seven days. Once removed from DSS, only one wild-type mouse tested positive and only for one day, for occult blood in the stool (Figure 1A), yet TLR9-deficient mice tested positive up to day 4 after cessation of DSS treatment (Figure 1D). Even more pronounced was the change in stool consistency with only one wild-type mouse showing very soft or loose stool on days 8 and 9 (Figure 1B), yet all TLR9-deficient mice continued showing clinical signs seven days after being returned to regular drinking water (Figure 1E). Wild-type mice maintained consistent weight throughout the 14 days, suggesting that the mild clinical parameters were not severe enough to produce weight loss. In contrast, TLR9-deficient mice showed a significant (p < 0.05; Student's t-test) increase in weight during the recovery period following the loss during the treatment (Figure 1C). Improvement of occult blood scores in these mice by day 10–11 suggest that enough of the intestine has been repaired to restore nutrient uptake, which correlates with a weight gain during the recovery period (Figure 1C). It is important to note, that the TLR9-deficient mice have not returned to their baseline weight by day 14 (net 5% loss over the experiment). Measurement of colon length, another indicator of intestinal damage, at day 14 showed that 1% DSS-treated, TLR9-deficient, mice had significantly (p < 0.05; Student's t-test) shorter colons than similarly treated, wild-type mice (Figure 1F). We conclude that TLR9-deficient mice have delayed intestinal repair following induction of injury, which indicates a potential role for this receptor in initiating epithelial cell restitution.
TLR9-deficient mice have delayed repair of epithelial damage, which correlates with increased leukocyte infiltration
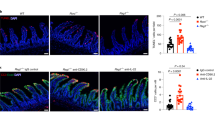
To determine whether gross clinical disease parameters correlated with histologic intestinal injury, we examined colon sections from untreated wild-type and TLR9-deficient mice (day 0) and 1% DSS treated mice at days 7 and 14 of the experiment. Untreated wild-type and TLR9-deficient mice showed no signs of intestinal damage in H&E-stained colon sections (Figures 2A, D). Treatment of wild-type or TLR9-deficient mice with 1% DSS resulted in partial or complete crypt loss and increased leukocyte infiltration into the mucosa by day 7 (Figures 2B, E). However, seven days after cessation of DSS treatment, wild-type mice showed no epithelial damage and only very minor resolving leukocyte infiltration (Figure 2C). TLR9-deficient mice treated with 1% DSS for seven days, followed by regular drinking water for seven days, still showed loss of colonic crypts and leukocyte infiltration (Figure 2F). Blinded scoring of histologic sections for all mice showed that wild-type mice had no epithelial damage, but all TLR9-deficient mice still had significant (p < 0.01; Mann Whitney test) damage at day 14 (Figure 2G). Furthermore, TLR9-deficient mice showed significantly (p < 0.01; Mann Whitney test) more leukocyte infiltration at day 14 than wild-type mice (Figure 2H). We conclude that TLR9 deficient mice have delayed epithelial restitution and delayed clearance of intestinal inflammation following DSS-induced intestinal damage.

1% DSS-treated, TLR9-deficient mice showed delayed intestinal wound healing with increased leukocyte infiltration.
Wild-type and TLR9-deficient mice were treated as in Figure 1. Colons were collected from untreated (day 0) and 1% DSS- treated (days 7, or 14) wild-type (A–C) and TLR9-deficient (D–F) mice. Representative H&E-stained sections (200X magnification) are shown (scale bars = 100 μm). The slides from all mice in the experiment were scored, in a blinded manner, for epithelial damage (G) and leukocyte infiltration (H) at day 14. n = 6 mice/group for each genotype. **p < 0.01, Mann Whitney test.
TLR9-deficient mice exhibit increased enterocyte proliferation
The observed differences in intestinal wound repair between the two genotypes (Figure 2) could potentially be due to inherent differences in the intestinal mucosa due to lack of TLR signaling. Since MyD88-deficient mice show increased epithelial cell proliferation in the absence of DSS treatment7, we next asked whether the homeostatic environment of the colon was altered in the absence of TLR9. Similar to MyD88-deficient mice, the absence of TLR9 resulted in significantly (p < 0.05; Student's t-test) increased enterocyte proliferation, as indicated by positive Ki-67 labeling, in the basal state compared to wild-type mice (Figure 3A). Proliferating cells were distributed similarly to the wild-type mice but were numerous and extended farther up the crypt past the transient amplifying zone (Figures 4A, B). There are several possible explanations for the increased number of proliferative cells in the TLR9-deficient mice, some of which include: increased crypt length, increased numbers of differentiated epithelial cell types such as goblet cells, or increased apoptosis of differentiated cells. Quantification of crypt length and goblet cell numbers per crypt showed no significant (p > 0.05; Student's t-test) differences between the wild-type and TLR9-deficient mouse colons (Figures 3B, C; Figures 4C, D). There was an average of one TUNEL-positive cell per wild-type crypt, while TLR9-deficient crypts varied from 1–3 TUNEL-positive cells (p > 0.05, not significant; Student's t-test, 50 crypts each from six independent mice) (Figure 3D; Figures 4E, F). Since there was an increase in proliferating cells without a compensatory increase in crypt length or number of apoptotic cells, we conclude that TLR9-deficient mice may have a defect in the differentiation of precursors into enterocytes.

Untreated, TLR9-deficient mice have increased numbers of proliferating colonic epithelial cells, but not increased crypt length.
50 crypts from six independent untreated mice (300 total crypts) of each genotype were analyzed for: (A) number of Ki-67+ cells per colonic crypt, (B) colonic crypt length, (C) goblet cells per colonic crypt and (D) number of TUNEL+ cells per colonic crypt. Error bars represent ± standard error of the means (n = 6). *p < 0.05, not significant; Student's t-test.

Representative images for quantitative histology.
Images from colons of untreated wild-type (A, C, E) and TLR9-deficient (B, D, F) mice. (A, B) Sections were stained for Ki-67 (red), counterstained with DAPI (blue) and imaged at 630X. (C, D) Sections were stained with PAS for goblet cells and imaged at 200X. (E, F) Sections were TUNEL stained for apoptotic cells (green), counterstained with DAPI (blue) and imaged at 630X. White arrows indicate examples of Ki-67, or TUNEL, positive cells. All scale bars = 50 μm.
TLR9 signaling is important for the induction of VEGF and intestinal progenitor cell differentiation
Based on the importance of TLR9-induced production of VEGF in healing of skin wounds38,39,40, we next asked whether homeostatic levels of VEGF were altered in the absence of TLR9. We quantified VEGF expression in distal colon sections from untreated wild-type and TLR9-deficient control mice using real-time RT-PCR. Untreated TLR9-deficient mice showed markedly lower (5-fold) VEGF expression compared to the wild-type mice (Figure 5A). We conclude that the colonic environment may be altered in the absence of TLR9 and this might provide a potential mechanism by which TLR9 signaling could mediate intestinal wound repair and play a role in normal host biology.

TLR9-deficient mice have reduced VEGF and HES1 expression in the distal colon.
Relative expression of VEGF (A) and HES1 (B) in distal colons isolated from untreated wild-type and TLR9-deficient mice analyzed using relative quantitative real-time RT-PCR. VEGF or HES1 expression is shown for untreated wild-type or TLR9-deficient mice relative to untreated wild-type mice. Each point was normalized first to a housekeeping gene and then to the wild-type level. Each dot represents one mouse (n = 8).
Because we observed increased numbers of proliferating cells extending past the transient amplifying zone (Figures 4A, B), we next asked whether expression of factors important for differentiation of progenitor cells into mature enterocytes was reduced in the absence of TLR9. HES1 induces differentiation of progenitor cells through Notch signaling2,41,42. In distal colon segments from TLR9-deficient mice (n = 6 of 8), HES1 expression was 5-fold reduced compared to wild-type mice (Figure 5B). There was some variability in expression of HES1 in the TLR9-deficient animals suggesting that TLR9 is not the only mechanism by which HES1 expression is regulated. From these results we conclude that, although there are compensatory TLR9-independent mechanisms, TLR9 is necessary for optimal expression of differentiation factors necessary for mature enterocyte production. These studies provide insight into the mechanism by which TLR9 promotes resolution of DSS-induced intestinal epithelial damage.
Discussion
Although the roles of innate immune receptors and their signaling pathways in acute intestinal epithelial cell injury have been well-characterized7,8,9,10,34,35, the contribution of these receptors to wound repair during the recovery phase is less understood. Because IBD produce periodic episodes of injury and repair21,22, understanding the role of innate immune receptors in repair is of critical importance for people affected by IBD. Using a mouse model of IBD, we showed that mice deficient in TLR9 exhibited significantly increased susceptibility to intestinal damage and delayed wound healing (Figures 1 & 2). Untreated TLR9-deficient mice had increased numbers of proliferating enterocytes located not only within, but also above, the transient amplifying zones of colonic crypts (Figures 3, 4), similar to previous observations with MyD88-deficient mice7. Increased proliferation did not result in increased crypt length, goblet cell numbers, or numbers of apoptotic cells (Figure 3), indicating the possibility of reduced intestinal progenitor cell differentiation. Interestingly, TLR9-deficient colons displayed dramatically reduced expression of HES1, an intestinal progenitor cell differentiation factor important for enterocyte production and VEGF, a growth factor important for epithelial restitution (Figure 5). Embryonic lethality of HES1-deficient mice precludes our ability to directly address the role of this protein in intestinal repair43. Based on our results, absence of TLR9 contributed to increased sensitivity to chemically-induced damage and reduced ability to repair the intestine. Together these studies suggest that TLR9 plays a key role in homeostatic processes in the colon.
Previously published studies present conflicting data on the role of TLR9 in acute intestinal injury following DSS treatment34,35. Rachmilewitz et al. treated mice with 1.75% DSS and saw no differences between wild-type and TLR9-deficient mice34. In contrast, Lee et al. observed increased sensitivity in TLR9-deficient mice treated with 1.5% DSS35. We also observed increased susceptibility at 1% DSS treatment, supporting the later findings35. It is possible that the observed differences were due to mice that had been backcrossed to C57BL/6 for different numbers of generations, or variability in microbiota composition34,35. Another potential explanation for the differences among these studies is that higher concentrations of DSS caused intestinal injury that was severe enough to be indistinguishable between wild-type and TLR9-deficient mice. In support of this model, we observed that both wild-type and TLR9-deficient mice treated with ≥ 2% DSS exhibited a similar severity of clinical and histologic parameters (data not shown). Therefore, small differences in DSS concentration can dramatically affect development of colitis and we propose that the use of lower DSS concentrations (< 1.5%) is necessary when designing studies to evaluate intestinal repair.
TLR9 may play an important role in normal biological processes, such as wound repair38,39,40. Previous studies have shown that there are few viral infection models in which TLR9-deficient mice are more susceptible44. In contrast, in models of acute DSS-induced experimental colitis, pre-treatment of mice with CpG ODN induces type I interferon that attenuates disease severity via a TLR9-dependent mechanism45. More recently, Sato et al. proposed a role for TLR9 signaling in accelerated repair in a murine model of skin biopsy wounding40. In that study, wild-type and TLR9-deficient mice showed equal magnitude and kinetics of early repair, but after several days, TLR9-deficient mice showed a delayed repair40. We observed a similar delay in intestinal wound healing (Figures 1 & 2) suggesting that TLR9 might play a role in normal repair processes. In the skin, CpG ODN treatment induced production of VEGF and neutralization of VEGF with specific antibody treatment delayed wound healing40. Similarly, we observed that TLR9-deficient mice produced greatly reduced VEGF expression levels compared to wild-type mice (Figure 5). These data, along with those demonstrating delayed wound repair in the absence of TLR9, indicate that TLR9-dependent regulation of expression of specific genes likely plays a critical role for intestinal repair.
We observed that HES1 expression was reduced, but not completely absent, in TLR9-deficient mice. Previous studies on intestinal development show that targeted disruption of HES1 resulted in increased numbers of goblet cells and decreased numbers of enterocytes in day E17 embryos41. HES1 is a repressor of MATH146 and deletion of MATH1 resulted in absence of secretory cells (Paneth, goblet and enteroendocrine) in the fetal intestine47. Intestinal-specific deletion of MATH1 also resulted in the absence of secretory cells in the adult intestine48. Although MATH1-deficient crypts from these mice displayed delayed restitutive responses, heterozygous mice showed normal pancreatic development suggesting that small amounts of HES1 were sufficient for development. Similar studies have not been performed on the intestine41, leaving open the possibility that reduced, but not absent, HES1 (and thus altered MATH1) expression, as we see in the absence of TLR9, might result in subtle phenotypic differences in adult intestine.
HES1 expression was reduced, but not completely absent, in TLR9-deficient mice; therefore, it is possible that low levels of HES1 expression are sufficient to prevent increased MATH1 expression. We did not observe differences in MATH1 expression between untreated wild-type and TLR9-deficient mice (not shown). Since there is no change in goblet cell numbers in TLR9-deficient mice, there may be enough HES1 and MATH1 expression to support goblet cell development, but when the intestine is stressed and damaged, this level may not be enough to support wild-type kinetics of reconstitution.
Whether the TLR9 ligands in the intestine are derived from the host or microbes is an open question. The intestine is colonized with commensal bacteria13 and maintains a delicate balance with the host immune response. Loss of this balance has been implicated in the development of IBD19,20. Identifying the individual contributions of bacterial or host DNA is difficult because antibiotic treatment can reduce the number of bacteria present in the intestine significantly, but not completely29 and therefore these types of studies are inconclusive. While gnotobiotic mice allow investigation of responses in the absence of microbial contributions, these mice have defects in intestinal and immune development7,14,15,16,17,18. Host DNA can contribute to autoinflammation by complexing with LL-37, a host-derived antimicrobial peptide and inducing TLR9-dependent type I interferon from plasmacytoid dendritic cells in the skin of psoriasis patients38,49. Regardless of the source of DNA, therapeutic approaches to improve wound repair or reduce intestinal damage via alteration of TLR9 signaling would be the same.
Pre-treatment of wild-type mice with probiotics containing CpG DNA reduced the severity of DSS-induced colitis in a TLR9-dependent manner50. Beneficial effects of probiotic pretreatment are likely from apical stimulation of enterocytes35. Signaling from the basolateral surface, upon barrier disruption, results in an NF-kB-mediated inflammatory response35. This differential signaling from apical or basolateral surfaces could explain why treatment of CpG DNA once intestinal damage occurs, exacerbates inflammatory pathology. Further studies are required to determine if intestinal repair mediated by TLR9 is due to apical or basolateral signaling.
Collectively, we have demonstrated a role for TLR9 in protection against intestinal injury and provide the first report for the importance of TLR9 in intestinal wound repair. We also showed that TLR9 deficiency, similar to MyD88 deficiency, results in increased numbers of proliferating crypt cells. In the absence of TLR9, expression of HES1 was reduced. These data are consistent with published observations of TLR9 and MyD88 importance for resistance to DSS-induced intestinal injury7,35 and the contribution of MyD88-dependent signaling pathways to intestinal wound repair7. Additionally, our results show that TLR9 signaling is involved in intestinal wound repair similar to observations in skin wounding models38,39,40. Because of the recurrent nature of IBD21,22, understanding the role of TLR9 in intestinal wound healing is of critical importance for people affected by IBD. Our results provide important insights into the mechanisms involved in intestinal wound repair following activation of innate immune signaling pathways and provide novel targets for potential therapeutics. Our data also suggest future studies are necessary to determine the role of TLR signaling in intestinal epithelial crypt progenitor cell differentiation.
Methods
Mice
C57BL/6 (WT) and TLR9 receptor-deficient mice were bred and housed under specific pathogen-free conditions in the Cornell University transgenic mouse core facility. TLR9-deficient mice B6.129P2-TLR9tmAki were obtained from S. Akira via the National Institutes of Health. All animal experiments were approved by Cornell University's Institutional Animal Care and Use Committee (Animal Welfare Assurance A3347-01). Cornell University is accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International. Female mice used for experiments were aged 8–10 weeks and weighed between 15–20 grams at the start of experiments.
Induction of colitis
DSS at 1% weight per volume (36,000–50,000 Da; MP Biomedical, Solon, OH) was administered ad libitum in drinking water with fresh DSS supplied every three days. Control mice received regular drinking water for all fourteen days of the experiment. There were 6 mice per group in two independent experiments, for a total of 12 total mice per group.
Clinical scoring
Mice were monitored daily for weight, stool consistency and presence of occult blood. Net weight change during treatment was calculated as: % Weight Change = (weight at day 7 – weight at day 0/weight at day 0) x 100. Net weight change during recovery was calculated similarly: (day 14 – day 7/day7) x 100. Stool consistency and occult blood (Hemoccult® SENSA®, Beckman Coulter, Brea, CA) were scored as previously described51. Briefly, stool consistency was scored as: 1) soft but still formed; 2) very soft stool; or 3) diarrhea. Occult blood was scored as: 1) a positive Hemoccult® test; 2) visible blood in the stool; or 3) rectal bleeding.
Tissue collection and histopathologic scoring
Mice were euthanized at days 7 or 14 of the experiment and whole colons were excised. Length was measured and a segment (0.5 cm) from the distal colon was collected for RNA preparation. Following fixation (10% formalin, Azer Scientific, Morgantown, PA), the remaining colon was paraffin-embedded, sectioned (5 μm) and stained by hematoxylin and eosin (H&E) or periodic acid-Schiff (PAS) at the Cornell University Animal Health Diagnostic Center. H&E-stained sections were scored, by a board-certified pathologist, in a blinded manner as previously described52.
Immunofluorescent staining and histology
Sections cut from the paraffin-embedded colons were deparaffinized using xylene (Sigma, St. Louis, MO) then rehydrated through a graded ethanol series (Sigma). Heat-induced epitope retrieval was performed by microwaving the slides at 800 watts for 20 minutes in a 10 mM Sodium Citrate (Sigma) pH 6.0 solution with 0.05% Tween-20 (Sigma). Apoptotic cells were stained using the In Situ Cell Death Detection kit, Fluorescein (Roche, Indianapolis, IN) according to manufacturer's instructions. Proliferating cells were stained using a Mouse on Mouse kit (Vector, Burlingame, CA) with a monoclonal Ki-67 (Clone MM1; Leica, Buffalo Grove, IL) primary antibody and a streptavidin-conjugated Alexa Fluor 555 (Life Technologies, Grand Island, NY) detection antibody. After mounting in Prolong Gold Anti-fade Reagent with DAPI (Life Technologies), slides were imaged with an Axio Imager M1 microscope (Zeiss, Thornwood, NY) and an Axiocam MRm (Zeiss). For histological analysis, sections were stained for H&E or PAS and images were taken with an Axio Imager M1 microscope and an Axiocam HRc (Zeiss). Crypt length was quantified from the H&E stained sections by measuring from the basolateral surface of the basal crypt cells to the apical cell surface at the top of intact crypts using the length function in the AxioVision Rel. 4.8 software (Zeiss). PAS-stained sections were used to quantify the number of goblet cells per crypt. Measurements for each quantitative outcome were collected from 50 crypts analyzed from the distal to proximal colon of six independent mice (300 total crypts per parameter).
Real-time RT-PCR
The distal colon segment (0.5 cm) from each mouse was collected in 600 μl of RLT buffer (Qiagen), homogenized with a Power Gen 125 (Thermo Fisher Scientific, Pittsburg, PA) and total RNA was isolated using the RNeasy Mini kit (Qiagen) according to the manufacturer's instructions. RNA from the murine primary colonic epithelial cells was also isolated using the RNeasy Mini kit. RNA was quantified using a NanoDrop® ND-1000 spectrophotometer (NanoDrop, Wilmington, DE) and 1 μg of RNA was reverse-transcribed using SuperScript® III First-Strand Synthesis (Life Technologies). RNA abundance was compared using SYBR Green Supermix (Life Technologies) on an ABI PRISM 7500 (Life Technologies). The 2−ΔΔCT method53 was used to calculate relative expression of HES1 or VEGF following normalization to the housekeeping gene glyceraldehyde 3-phsophate dehydrogenase and then normalization to the wild-type level. Primer sequences are listed in Table 1.
Statistical analysis
All statistical analyses were performed as described in the figure legends using Prism software package v5 (Graphpad, La Jolla, CA).
References
Sancho, E., Batlle, E. & Clevers, H. Live and let die in the intestinal epithelium. Curr. Opin. Cell Biol. 15, 763–770 (2003).
van der Flier, L. G. & Clevers, H. Stem cells, self-renewal and differentiation in the intestinal epithelium. Annu. Rev. Physiol. 71, 241–260 (2009).
Shaker, A. & Rubin, D. C. Intestinal stem cells and epithelial-mesenchymal interactions in the crypt and stem cell niche. Transl. Res. 156, 180–187 (2010).
Joo, Y. E. et al. Tomato lycopene extract prevents lipopolysaccharide-induced NF-kappaB signaling but worsens dextran sulfate sodium-induced colitis in NF-kappaBEGFP mice. PLoS ONE 4, e4562 (2009).
Iwaya, H. et al. Marginal zinc deficiency exacerbates experimental colitis induced by dextran sulfate sodium in rats. J. Nutr. 141, 1077–1082 (2011).
Iizuka, M. & Konno, S. Wound healing of intestinal epithelial cells. World J. Gastroenterol. 17, 2161–2171 (2011).
Rakoff-Nahoum, S., Paglino, J., Eslami-Varzaneh, F., Edberg, S. & Medzhitov, R. Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis. Cell 118, 229–241 (2004).
Fukata, M. et al. Toll-like receptor-4 is required for intestinal response to epithelial injury and limiting bacterial translocation in a murine model of acute colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 288, G1055–1065 (2005).
Cario, E., Gerken, G. & Podolsky, D. K. Toll-like receptor 2 controls mucosal inflammation by regulating epithelial barrier function. Gastroenterology 132, 1359–1374 (2007).
Vijay-Kumar, M. et al. Deletion of TLR5 results in spontaneous colitis in mice. J. Clin. Invest. 117, 3909–3921 (2007).
Vermeulen, N. et al. Identification of a novel autoantigen in inflammatory bowel disease by protein microarray. Inflamm. Bowel Dis. 17, 1291–1300 (2011).
Cario, E. Toll-like receptors in inflammatory bowel diseases: a decade later. Inflamm. Bowel Dis. 16, 1583–1597 (2010).
Savage, D. C. Microbial ecology of the gastrointestinal tract. Annu. Rev. Microbiol. 31, 107–133 (1977).
Stappenbeck, T. S., Hooper, L. V. & Gordon, J. I. Developmental regulation of intestinal angiogenesis by indigenous microbes via Paneth cells. Proc. Natl. Acad. Sci. U. S. A. 99, 15451–15455 (2002).
Mazmanian, S. K., Liu, C. H., Tzianabos, A. O. & Kasper, D. L. An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell 122, 107–118 (2005).
Hooper, L. V., Xu, J., Falk, P. G., Midtvedt, T. & Gordon, J. I. A molecular sensor that allows a gut commensal to control its nutrient foundation in a competitive ecosystem. Proc. Natl. Acad. Sci. U. S. A. 96, 9833–9838 (1999).
Turnbaugh, P. J. et al. A core gut microbiome in obese and lean twins. Nature 457, 480–484 (2009).
Mazmanian, S. K., Round, J. L. & Kasper, D. L. A microbial symbiosis factor prevents intestinal inflammatory disease. Nature 453, 620–625 (2008).
Hooper, L. V. & Gordon, J. I. Commensal host-bacterial relationships in the gut. Science 292, 1115–1118 (2001).
Abt, M. C. & Artis, D. The intestinal microbiota in health and disease: the influence of microbial products on immune cell homeostasis. Curr. Opin. Gastroenterol. 25, 496–502 (2009).
Kaser, A., Zeissig, S. & Blumberg, R. S. Inflammatory bowel disease. Annu. Rev. Immunol. 28, 573–621 (2010).
Khor, B., Gardet, A. & Xavier, R. J. Genetics and pathogenesis of inflammatory bowel disease. Nature 474, 307–317 (2011).
Hugot, J. P. et al. Association of NOD2 leucine-rich repeat variants with susceptibility to Crohn's disease. Nature 411, 599–603 (2001).
Kaser, A. et al. XBP1 links ER stress to intestinal inflammation and confers genetic risk for human inflammatory bowel disease. Cell 134, 743–756 (2008).
Cadwell, K. et al. Virus-plus-susceptibility gene interaction determines Crohn's disease gene Atg16L1 phenotypes in intestine. Cell 141, 1135–1145 (2010).
Sellon, R. K. et al. Resident enteric bacteria are necessary for development of spontaneous colitis and immune system activation in interleukin-10-deficient mice. Infect. Immun. 66, 5224–5231 (1998).
Rath, H. C. et al. Different subsets of enteric bacteria induce and perpetuate experimental colitis in rats and mice. Infect. Immun. 69, 2277–2285 (2001).
Kuhn, R., Lohler, J., Rennick, D., Rajewsky, K. & Muller, W. Interleukin-10-deficient mice develop chronic enterocolitis. Cell 75, 263–274 (1993).
Elinav, E. et al. NLRP6 Inflammasome Regulates Colonic Microbial Ecology and Risk for Colitis. Cell 145, 745–757 (2011).
Kawai, T. & Akira, S. Toll-like receptors and their crosstalk with other innate receptors in infection and immunity. Immunity 34, 637–650 (2011).
Cooper, H. S., Murthy, S. N., Shah, R. S. & Sedergran, D. J. Clinicopathologic study of dextran sulfate sodium experimental murine colitis. Lab. Invest. 69, 238–249 (1993).
Kitajima, S., Takuma, S. & Morimoto, M. Changes in colonic mucosal permeability in mouse colitis induced with dextran sulfate sodium. Exp. Anim. 48, 137–143 (1999).
Kitajima, S., Takuma, S. & Morimoto, M. Histological analysis of murine colitis induced by dextran sulfate sodium of different molecular weights. Exp. Anim. 49, 9–15 (2000).
Rachmilewitz, D. et al. Toll-like receptor 9 signaling mediates the anti-inflammatory effects of probiotics in murine experimental colitis. Gastroenterology 126, 520–528 (2004).
Lee, J. et al. Maintenance of colonic homeostasis by distinctive apical TLR9 signalling in intestinal epithelial cells. Nat. Cell Biol. 8, 1327–1336 (2006).
Carvalho, F. A., Aitken, J. D., Gewirtz, A. T. & Vijay-Kumar, M. TLR5 activation induces secretory interleukin-1 receptor antagonist (sIL-1Ra) and reduces inflammasome-associated tissue damage. Mucosal Immunol. 4, 102–111 (2011).
Krishnan, K., Arnone, B. & Buchman, A. Intestinal growth factors: potential use in the treatment of inflammatory bowel disease and their role in mucosal healing. Inflamm. Bowel Dis. 17, 410–422 (2011).
Gregorio, J. et al. Plasmacytoid dendritic cells sense skin injury and promote wound healing through type I interferons. J. Exp. Med. 207, 2921–2930 (2010).
Guiducci, C. et al. Autoimmune skin inflammation is dependent on plasmacytoid dendritic cell activation by nucleic acids via TLR7 and TLR9. J. Exp. Med. 207, 2931–2942 (2010).
Sato, T., Yamamoto, M., Shimosato, T. & Klinman, D. M. Accelerated wound healing mediated by activation of Toll-like receptor 9. Wound Repair Regen. 18, 586–593 (2010).
Jensen, J. et al. Control of endodermal endocrine development by Hes-1. Nat. Genet. 24, 36–44 (2000).
Okamoto, R. et al. Requirement of Notch activation during regeneration of the intestinal epithelia. Am. J. Physiol. Gastrointest. Liver Physiol. 296, G23–35 (2009).
Yu, X. et al. HES1 inhibits cycling of hematopoietic progenitor cells via DNA binding. Stem Cells 24, 876–888 (2006).
Hirsch, I., Caux, C., Hasan, U., Bendriss-Vermare, N. & Olive, D. Impaired Toll-like receptor 7 and 9 signaling: from chronic viral infections to cancer. Trends Immunol. 31, 391–397 (2010).
Katakura, K. et al. Toll-like receptor 9-induced type I IFN protects mice from experimental colitis. J. Clin. Invest. 115, 695–702 (2005).
Zheng, J. L., Shou, J., Guillemot, F., Kageyama, R. & Gao, W. Q. Hes1 is a negative regulator of inner ear hair cell differentiation. Development 127, 4551–4560 (2000).
Yang, Q., Bermingham, N. A., Finegold, M. J. & Zoghbi, H. Y. Requirement of Math1 for secretory cell lineage commitment in the mouse intestine. Science 294, 2155–2158 (2001).
Shroyer, N. F. et al. Intestine-specific ablation of mouse atonal homolog 1 (Math1) reveals a role in cellular homeostasis. Gastroenterology 132, 2478–2488 (2007).
Lande, R. et al. Plasmacytoid dendritic cells sense self-DNA coupled with antimicrobial peptide. Nature 449, 564–569 (2007).
Rachmilewitz, D. et al. Immunostimulatory DNA ameliorates experimental and spontaneous murine colitis. Gastroenterology 122, 1428–1441 (2002).
Wirtz, S., Neufert, C., Weigmann, B. & Neurath, M. F. Chemically induced mouse models of intestinal inflammation. Nat. Protoc. 2, 541–546 (2007).
Jawhara, S. & Poulain, D. Saccharomyces boulardii decreases inflammation and intestinal colonization by Candida albicans in a mouse model of chemically-induced colitis. Med. Mycol. 45, 691–700 (2007).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402–408 (2001).
Acknowledgements
We thank members of the Leifer lab for helpful discussions and Jeff Mills and Chia-Hsin Ju for review of the manuscript.
Author information
Authors and Affiliations
Contributions
WR and CL designed the experiments, WR performed the experiments, KS performed pathology and WR and CL interpreted the data and wrote the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-No Derivative Works 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
II, W., Sakamoto, K. & Leifer, C. TLR9 is important for protection against intestinal damage and for intestinal repair. Sci Rep 2, 574 (2012). https://doi.org/10.1038/srep00574
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep00574
This article is cited by
-
Engaging natural antibody responses for the treatment of inflammatory bowel disease via phosphorylcholine-presenting nanofibres
Nature Biomedical Engineering (2023)
-
Intestinal serotonergic system is modulated by Toll-like receptor 9
Journal of Physiology and Biochemistry (2022)
-
Epithelial sensing of microbiota-derived signals
Genes & Immunity (2021)
-
Epithelial Toll-like receptors and their role in gut homeostasis and disease
Nature Reviews Gastroenterology & Hepatology (2020)
-
Effect of polyunsaturated fatty acids on postnatal ileum development using the fat-1 transgenic mouse model
Pediatric Research (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
