
Abstract
Secondary bacterial infection in humans is one of the pathological conditions requiring clinical attention. In this study, we examined the effect of lipopolysaccharide (LPS) on encephalomyocarditis virus (EMCV) infected mice. All mice inoculated with EMCV at 5 days before LPS challenge died within 24 h. LPS-induced TNF-α mRNA expression was significantly increased in the brain and heart at 5 days after EMCV infection. CD11b+/TLR4+ cell population in the heart was remarkably elevated at 5 days after EMCV infection and sorted CD11b+ cells at 5 days after EMCV infection produced a large amount of TNF-α on LPS stimulation in vivo and in vitro. In conclusion, we found that the infiltration of CD11b+ cells into infected organs is involved in the subsequent LPS-induced lethal shock in viral encephalomyocarditis. This new experimental model can help define the mechanism by which secondary bacterial infection causes a lethal shock in viral encephalomyocarditis.
Similar content being viewed by others

Introduction
Polymicrobial infectious diseases show the involvement of 2 or more microbes, including viruses, bacteria, fungi or parasites and these microbes act synergistically to mediate complex disease processes. In particular, a bacterial infection superimposed over an acute viral infection such as influenza is well known as the aggravation factor in the infectious disease1,2. Lipopolysaccharide (LPS), the outer membrane of gram-negative bacteria, causes systemic inflammatory response syndrome, endotoxic shock, disseminated intravascular coagulation and multi-organ failure3,4. LPS is recognized by Toll-like receptor 4 (TLR4)-expressing immunocompetent cells, mainly monocytes and macrophages and induces the production of inflammatory cytokines through NF-κB activation3. Toxic effects of LPS are partially induced by the release and action of macrophage-derived inflammatory cytokines. Especially, the mass production of tumor necrosis factor-α (TNF-α) causes septic shock with an abrupt reduction in blood pressure, leading to rapid aggravation of the disease condition5,6. It is known that LPS-induced lethal shock is caused by a large quantity of TNF-α produced by immune cells activated by some kind of pre-stimulation7,8.
Encephalomyocarditis virus (EMCV), which is a single-stranded RNA virus and a member of Cardiovirus in the Picornaviridae family, causes acute myocarditis and encephalitis in various animal species9,10. In some reports, including our previous study, the death of mice by EMCV infection of high density (500 plaque-forming units (pfu)) occurred after 6 days and the mice that survived for 12 days subsequently recovered. Additionally, remarkable inflammation in the brain and heart occurred at around 6 days after EMCV infection and thereafter the inflammation reduced as the days progressed. Hence, these results suggested that inflammatory cells reach the activating stage around 6 days after EMCV infection11,12,13. It was also demonstrated that various cells under the activating stage augment reactivity to LPS stimulation14,15,16. Previous studies showed that TNF-α is produced in large quantities by subsequent LPS stimulation during adenovirus infection17, lymphocytic choriomeningitis virus and varicella-zoster virus infection18, but the original source of TNF-α is not elucidated. Moreover, it is unknown how LPS affects in the viral encephalomyocarditis. The aim of our study is to examine the effect of LPS on EMCV-infected mice and to characterize the infiltrating cells, which possibly produce TNF-α, into the heart in this model.
Results
Survival of mice after LPS stimulation during EMCV infection
The mice were intraperitoneally inoculated with 20 pfu of EMCV and were intravenously injected with 10 μg LPS at 0, 2 and 5 days after the EMCV inoculation. At 5 days after EMCV infection, all mice died after LPS treatment within 24 h, but the mice subjected to this treatment at 0 and 2 days after EMCV infection were alive (Table 1). Moreover, at 5 days after EMCV infection, LPS-induced lethal shock developed in the mice in an LPS dose-dependent manner (Table 2).
To compare the survival rate between WT and TNF-α KO mice after LPS treatment during EMCV infection, the mice were treated intravenously with 10 μg LPS at 5 days after EMCV infection. All WT mice died after LPS treatment within 24 h at 5 days after EMCV infection, whereas TNF-α KO mice did not die. Moreover, LPS-induced lethality in WT mice was improved by anti-TNF-α antibody (100 μg/mouse) treatment before LPS stimulation (Table 3).
Effect of EMCV infection on LPS-induced TNF-α production
The concentration of serum TNF-α was measured at 0, 2 and 8 h after LPS treatment in each EMCV infection period. LPS-induced TNF-α concentration in the serum was markedly increased at 5 days after EMCV infection than at 0 and 2 days after EMCV infection (Fig. 1A).

Effect of EMCV infection on LPS-induced TNF-α production.
The concentration of serum TNF-α was determined at 0, 2 and 8 h after LPS treatment in each EMCV infection period (A). TNF-α mRNA expression in the brain, heart, liver, lung and spleen at 0, 1 and 8 h after LPS treatment in each EMCV infection period was determined on the basis of 18S rRNA expression using real-time RT-PCR. The data were calculated referring to mRNA levels of the respective tissues in control mice (0 days after EMCV infection, 0 hours after LPS inoculation) (B). The data are represented as means ± SD of the results of 4 mice in each group. *p<0.05
For the localization of TNF-α production, TNF-α mRNA expression was determined in the brain, heart, liver, lung and spleen at 0, 1 and 8 h after LPS treatment. TNF-α mRNA expression in the brain, heart and liver of mice was significantly increased at 5 days after EMCV infection as compared with that at 0 and 2 days after EMCV infection (Fig. 1B).
Histopathological finding and viral load of the tissues after EMCV infection
Histological changes were observed in the brain and heart. The mice were inoculated intraperitoneally with 20 pfu of EMCV. Neuronal cell death and the presence of some inflammatory cells in hippocampus or brain surface of the brain and marked infiltration of inflammatory cells in the heart were seen at 5 days after EMCV infection. The infiltrating cells mainly included macrophages and neutrophils (Fig. 2A). In the liver, a little infiltration of inflammatory cells was observed at 5 days after EMCV infection and no remarkable change was seen in the lung during each infection period (Data not shown). All tissues at 8 h after LPS treatment in EMCV-infected mice were not observed the significant increase of inflammatory cells in comparison with the tissues before LPS treatment (data not shown).

Histopathological finding and viral load of the tissues after EMCV infection.
Histopathological examination in the brain and heart at 0, 2 and 5 days after EMCV infection was performed. Tissue sections were deparaffinized, stained with hematoxylin-eosin and examined under light microscopy. Scale bars, 200 μm (low-power field) and 50 μm (high-power field). These experiments were performed with 4 mice in each group and produced the same results (A). Viral RNA in the brain, heart, liver, lung and spleen at 0, 2 and 5 days after EMCV infection was analyzed by real-time RT-PCR and was determined on the basis of 18S rRNA expression. The data are represented as means ± SD of the results of 4 mice in each group. The statistical analysis was performed by comparing with Day 2 (B). *p<0.05
We measured the viral load in the brain, heart, liver, lung and spleen at 2 and 5 days after EMCV infection. Viral loads in all tissues at 5 days after EMCV infection were increased as compared with those at 2 days after EMCV infection. In particular, viral loads in the brain and heart markedly increased at 5 days after the infection (Fig. 2B).
Effect of EMCV infection on mRNA expression of chemokines in the tissues
Chemoattractant protein-1 (MCP-1) is a major chemoattractant responsible for the recruitment of macrophages19,20. macrophage inflammatory protein-2 (MIP-2) and keratinocyte-derived chemokine (KC), which are produced by macrophages, exhibit potent neutrophil chemotactic activity21,22. Because the infiltrated cells were mainly composed of macrophages and neutrophils as revealed by histopathological findings (Fig. 2A), the mRNA expressions of MCP-1, MIP-2 and KC were determined in the brain and heart at 0, 2 and 5 days after EMCV infection. The mRNA expression of these chemokines in the brain and heart on 5 days after EMCV infection was significantly up-regulated as compared with that at 0 and 2 days after EMCV infection (Fig. 3). MCP-1 mRNA expression at 5 days after EMCV infection was 808-fold in the brain and 125-fold in the heart based on 0 days. Similarly, MIP-2 mRNA expression was enhanced 254-fold in the brain and 44-fold in the heart and KC mRNA expression was increased 23-fold in the brain and also in the heart. Although the expression of chemokines in the liver, lung and spleen had a tendency to up-regulated at 5 days after EMCV infection, the up-regulation was not more remarkable than that in the brain or heart.

Effect of EMCV infection on mRNA expression of chemokines in the tissues.
The expression of MCP-1 (A), MIP-2 (B) and KC (C) mRNA in the brain, heart, liver, lung and spleen at 0, 2 and 5 days after EMCV infection was analyzed by real-time RT-PCR and was determined on the basis of 18S rRNA expression. The data were calculated referring to mRNA levels of the respective tissues in control mice (Day 0). The data are represented as means ± SD of the results of 6 mice in each group. *p<0.05
Effect of EMCV infection on TLR4 mRNA expression in tissues and infiltrating cells in the heart
The expression of TLR4 mRNA was examined in the brain, heart, liver, lung and spleen at 0, 2 and 5 days after EMCV infection. TLR4 mRNA expression in the brain, heart and liver at 5 days after EMCV infection was significantly increased. On the other hand, TLR4 mRNA expression in the lung and spleen showed no significant change after viral infection (Fig. 4A).

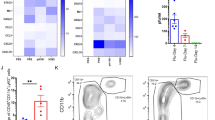
Effect of EMCV infection on TLR4 mRNA expression in tissues and infiltrating cells in the heart.
The expression TLR4 mRNA in the brain, heart, liver, lung and spleen at 0, 2 and 5 days after EMCV infection was analyzed by real-time RT-PCR and was determined on the basis of 18S rRNA expression. The data were calculated referring to mRNA levels of the respective tissues in control mice (Day 0). The data are represented means ± SD from 4 mice of each group (A). Following reaction with anti-CD16/CD32 antibody to suppress nonspecific binding, MNCs from the heart were stained with anti-CD3ε, anti-CD11b, anti-CD11c, anti-CD19, anti-CD49b and anti-TLR4 antibody. The positive rate of the cells was made on the basis of isotype control. The data are representative of three separate experiments (B). *p<0.05
Next, we determined the phenotype of mononuclear cells (MNCs) from the heart at 5 days after EMCV infection. Although a lot of CD11b+TLR4+ cells were contained in MNCs from the heart at 5 days after EMCV infection, few CD11c+, CD3+, CD19+ and CD49b+ cells were contained (Fig. 4B).
LPS-induced TNF-α production in CD11b+ cells from the heart in vivo and in vitro
Because MNCs from the heart at 5 days after EMCV infection were mainly occupied by CD11b+ cells, we evaluated the ability of LPS-induced TNF-α production in CD11b+ cells in vivo. The mice were intravenously inoculated with brefeldin A (250 μg/mouse) and LPS (10 μg/mouse) at 5 days after EMCV infection and MNCs from the heart were harvested at 1 h after LPS treatment. TNF-α producing cell in gated CD11b+ cells on the basis of isotype control were increased by LPS treatment (Fig. 5A).

LPS-induced TNF-α production in CD11b+ cells from the heart in vivo and in vitro.
The mice were inoculated intravenously with brefeldin A (250 μg/mouse) and LPS (10 μg/mouse) at 5 days after EMCV infection and MNCs from the heart were harvested at 1 h after LPS treatment. MNCs were stained with anti-CD11b and anti-TNF-α antibody. The data are representative of three separate experiments (A). Isolated CD11b+ and CD11b− MNCs and total cells before isolation were cultured at 1×105 cells per 200 μL with LPS (1 μg/mL) for 24 h. The concentration of TNF-α was determined in culture supernatant. The data are represented means ± SD in triplicate cultures (B). *p<0.05
CD11b+ and CD11b- cells from the heart at 5 days after EMCV infection were isolated by magnetic cell sorting. Isolated cells and total cells before isolation were cultured at 1×105 cells/200 μL with LPS (1 μg/mL) for 24 h. LPS-induced TNF-α production in CD11b+ cells was significantly increased as compared with that in total cells and CD11b− cells. CD11b− cells could not produce TNF-α after LPS stimulation (Fig. 5B).
Discussion
In this study, we demonstrated that LPS treatment at 5 days after EMCV infection induces an excess of TNF-α production in the brain and heart and lethal shock (Table 1, Fig. 1). Because this lethal effect of LPS was cancelled out in TNF-α KO mice infected with EMCV (Table 3), it is likely that TNF-α from the brain and heart plays a critical role in the lethal effect of LPS on EMCV infection. Septic shock is characterized by hypotension, decreased systemic vascular resistance and impaired vascular reactivity and TNF-α has been implicated as a principal mediator in the pathogenesis of septic shock23. TNF-α is mainly produced by macrophages in the peripheral tissue after pathogen infection and is involved in the elimination of pathogens from the host24,25. A protective role of TNF-α in EMCV-infected mouse model is previously reported11. In contrast, an excessive production of TNF-α induces hypothermia, hypotension, multiple organ failure, septic shock and death5,6. In particular, previous reports demonstrated that the macrophages treated with some pre-stimulants produce a large amount of TNF-α on subsequent LPS stimulation. Propionibacterium acnes, an anaerobic gram-positive bacterium, exerts strong immunomodulatory activities and participates in the formation of intrahepatic granulomas and induction of hypersensitivity for LPS in mice. Additionally, these activities depended on the recognition of bacteria via TLR9 and subsequent IL-12-mediated IFN-γ production26,27. EMCV infection also markedly increased the LPS-induced TNF-α mRNA level and viral road in the brain and heart (Fig. 1B, 2B). Namely, the susceptibility to LPS was enhanced in the EMCV-infected site.
TLR4 recognizes LPS from gram-negative bacteria and its recognition is essential for the activation of the innate immune system. In humans with myocarditis, TLR4 mRNA is also increased in the heart28. In the present study, the mRNA expression of TLR4 in the brain, heart and liver was increased after EMCV infection (Fig. 4A) and a significant increase in TNF-α mRNA expression was also confirmed after subsequent LPS stimulation (Fig. 1B). Histological findings revealed neuronal cell death and the presence of some inflammatory cells in hippocampus or brain surface of the brain, remarkable infiltration of inflammatory cells in the heart and a little infiltration of inflammatory cells in the liver at 5 days after EMCV infection; further, the infiltrating cells were mainly composed of macrophages and neutrophils. These results indicate that the accumulation of inflammatory cells in the heart after EMCV infection is associated with this lethal septic shock model. In fact, there is a correlation between TLR4 mRNA expression and the number of infiltrated TLR4 positive cells in the heart. A previous report also demonstrated the augmentation of TLR4 mRNA expression by infiltration of TLR4-positive leukocytes into the liver in an IL-17-induced multiple tissue inflammation model29. Furthermore, our previous studies proved the infiltration of TLR4-positive leukocytes into the liver on α-galactosylceramide administration and the excessive response to subsequent LPS stimulation8,15. In this study, CD11b+/TLR4+ cells in the heart increased at 5 days after EMCV infection, but few CD11c+, CD3+, CD19+ and CD49b+ cells were contained (Fig. 4B). LPS-induced TNF-α was mainly produced by CD11b+ cells in the heart at 5 days after EMCV infection in vivo and in vitro (Fig. 5). Therefore, it is suggested that the infiltration of CD11b+ cells into the heart involves in the up-reguration of TLR4 mRNA and subsequent LPS induced TNF-α production.
mRNA expression of MCP-1, MIP-2 and KC in the brain and heart at 5 days after EMCV infection was markedly increased than that in uninfected mice (Fig. 3). The enhancement of the mRNA expression of these chemokines, especially MCP-1, may be involved in the infiltration of inflammatory cells into the heart. MCP-1 acts as a potent chemoattractant and activator of monocytes/macrophages19,20. Shiratsuchi et al30 showed that macrophages phagocytose influenza virus-infected HeLa cells in a manner mediated by phosphatidylserine that appears on the surfaces of infected cells during the process of apoptosis. In addition, the inhibition of macrophage recruitment by MCP-1 augmented alveolar epithelial damage and apoptosis during influenza pneumonitis31. Thus, MCP-1 has an effective role in viral clearance, but it also may aggravate lethal shock by subsequent LPS stimulation under the EMCV-infected state.
Although the mice treated with 500 μg LPS died within 48 to 72 h in the general LPS-induced shock model32,33, all mice treated with 10 μg LPS died within 24 hours in our model; this suggests that LPS susceptibility in mice markedly rises under the EMCV-infected condition. It is known that infection by the influenza virus, coxsackie virus and adenovirus are involved in the onset of viral encephalitis or myocarditis34,35, but there are few reports showing the effect of the secondary bacterial superimposed infection on viral encephalomyocarditis and its mechanism. In the present study, even a low-dose of LPS could cause the lethal septic shock in EMCV-infected mice. Thus, it is necessary to monitor serum endotoxin levels and prevent bacterial infection in patients with encephalitis and myocarditis.
In conclusion, we established a septic shock model using mice with viral encephalomyocarditis and demonstrated that the infiltration of CD11b+ cells into tissues is involved in this lethal shock. From an immunological point of view, our animal model is useful for investigating the pathological state of bacterial superimposed infection on viral encephalomyocarditis.
Methods
Reagents
LPS from Escherichia coli O111:B5 and brefeldin A were purchased from Sigma-Aldrich (St Louis, MO, USA). Mouse TNF-α antibody (monoclonal rat IgG1) for neutralize was purchased from R&D Systems (Minneapolis, MN, USA).
Mice
Mouse experiments were performed according to the guidelines of the Animal Ethics Committee of Gifu University Graduate School of Medicine. C57BL/6J mice aged approximately 8–10 weeks were obtained from Japan SLC (Hamamatsu, Japan) and used as wild-type (WT) mice. TNF-α gene knockout (TNF-α KO) mice of a C57BL/6J background were produced by gene targeting as described previously11,36,37.
Viral inoculation and LPS treatment
A myocarditic variant of EMCV was generously provided by Dr. Seto (Keio University, Tokyo, Japan). The virus stock was stored at −80°C in Hanks' balanced salt solution (HBSS) with 0.1% BSA required for use. The mice were inoculated intraperitoneally with 20 pfu of EMCV in 0.2 mL saline and were treated intravenously with 10 μg LPS in 0.2 mL saline at 0 (non-infected), 2 and 5 days after the viral inoculation unless otherwise noted. The lethality was determined at 24 h after LPS treatment. The experiments were performed according to the institutional guidelines of Gifu University for microbiologic study.
Measurement of serum TNF-α concentration
The concentration of TNF-α in the serum and culture supernatant was determined using a mouse TNF-α Quantikine ELISA kit (R&D Systems, Minneapolis, MN, USA), according to the manufacturer's recommendations.
Real-time RT-PCR analysis
Real-time RT-PCR was used to quantify the levels of TNF-α, TLR4, MCP-1, MIP-2, KC mRNA and EMCV RNA. Total RNA in the brain, heart, liver, lung and spleen was isolated using Isogen (Nippon Gene, Tokyo, Japan) and transcribed to cDNA by using the High capacity cDNA transcription kit (Applied Biosystems, Foster City, CA, USA). Purified cDNA was used as the template for real-time PCR conducted using pre-designed primer/probe sets for TNF-α, TLR4, MCP-1, MIP-2, KC and 18S rRNA (Applied Biosystems), according to the manufacturer's recommendations. The determination of EMCV RNA was performed by the use of the LightCycler DNA Master SYBR Green I (Roche Diagnostic Systems, Indianapolis, IN, USA) and the following oligonucleotide primer pairs: EMCV sense, 5′-GTCGTGAAGGAAGCAGTTCC-3′, antisense, 5′-CACGTGGCTTTTGGCCGCAGAGGC-3′11. 18S rRNA was used as an internal control. Real-time PCR was carried out using a Light-Cycler rapid thermal cycler system (Roche Diagnostic Systems).
Histological examination
Histopathological examination of the brain and heart was performed at 0, 2 and 5 days after EMCV infection. The tissues were fixed in 10% formalin in PBS for 48 h and embedded in paraffin. Tissue sections were deparaffinized, stained with H&E and examined under light microscopy.
Cell isolation in heart
Single cells from the heart, particularly non-parenchymal cells, were isolated in reference to a previous report38. The heart was minced with scissors and was shaken with 5 mL HBSS containing collagenase type 2 (600 U/mL) and deoxyribonuclease I (60 U/mL) (Worthington, Lakewood, NJ, USA) for 30 min at 37°C. The specimen was filtered through a stainless steel mesh and red blood cells were lysed. After washing and filtration through a 70-μm cell strainer, the single cells were suspended in RPMI 1640 medium (Wako Pure Chemical Industries; Osaka, Japan) containing 10% heat-inactivated fetal bovine serum (Thermo Fisher Scientific Inc, Waltham, MA, USA) and cultured at 37°C in a 5% CO2 atmosphere.
Flow cytometry analysis
MNCs from the heart at 5 days after EMCV infection were obtained by centrifugation of the single cells with Ficoll-Conray (IBL, Gunma, Japan). Flow cytometry was used to evaluate the expression level of CD3, CD11b, CD11c, CD19, CD49b and TLR4 in MNCs from the heart at 5 days after EMCV infection. Following reaction with anti-CD16/CD32 antibody to suppress nonspecific binding, MNCs were stained with FITC-conjugated hamster anti-mouse CD3ε antibody (clone 145-2C11; BD Biosciences, Franklin Lakes, NJ, USA), FITC-conjugated rat anti-mouse CD11b antibody (clone M1/70; eBioscience, San Diego, CA, USA), FITC-conjugated hamster anti-mouse CD11c antibody (clone HL-3; BD Biosciences), FITC-conjugated mouse anti-mouse CD19 antibody (clone MB19-1; eBioscience), FITC-conjugated rat anti-mouse CD49b antibody (clone DX5; eBioscience) and PE-conjugated rat anti-mouse TLR4/MD2 complex antibody (clone MTS510; eBioscience). The positive rate of the cells was made on the basis of isotype control. The phenotypic characterization of the cells was carried out using FACSCanto II (BD Biosciences).
Intracellular cytokine staining in vivo
The mice were intravenously inoculated with brefeldin A (250 μg/mouse) and LPS (10 μg/mouse) at 5 days after EMCV infection. MNCs from the heart at 1 h after LPS treatment were fixed and permeabilized with the Cytofix/Cytoperm buffer (BD Biosciences) and were stained with PE-conjugated rat anti-TNF-α antibody (clone MP6-XT22; eBioscience). The cells were analyzed using FACSCanto II (BD Biosciences).
Isolation of CD11b+ cells
MNCs from the heart obtained at 5 days after EMCV infection were separated into CD11b+ and CD11b− cells using anti-CD11b-conjugated magnetic beads (Miltenyi Biotec GmbH, Bergisch Gladbach, Germany). The magnetically labeled cells were purified using the QuadroMACS separation unit attached to a MACS multistand and LS columns (Miltenyi Biotec GmbH).
Statistical analysis
In each experiment, the results were expressed as the mean ± SD. The statistical significance of the difference in mean values was determined by Student's t test or one-way analysis of variance followed by Scheffe's test. P values of less than 0.05 were considered significant.
References
Bakaletz, L. O. Developing animal models for polymicrobial diseases. Nat. Rev. Microbiol. 2, 552–568 (2004).
Brundage, J. F. Interactions between influenza and bacterial respiratory pathogens: implications for pandemic preparedness. Lancet Infect. Dis. 6, 303–312 (2006).
Cohen, J. The immunopathogenesis of sepsis. Nature 420, 885–891 (2002).
Bannerman, D. D. & Goldblum, S. E. Direct effects of endotoxin on the endothelium: barrier function and injury. Lab Invest 79, 1181–1199 (1999).
Tracey, K. J. & Cerami, A. Tumor necrosis factor: a pleiotropic cytokine and therapeutic target. Annu Rev Med 45, 491–503 (1994).
Brouckaert, P. & Fiers, W. Tumor necrosis factor and the systemic inflammatory response syndrome. Curr Top Microbiol Immunol 216, 167–187 (1996).
Gumenscheimer, M., Mitov, I., Galanos, C. & Freudenberg, M. A. Beneficial or deleterious effects of a preexisting hypersensitivity to bacterial components on the course and outcome of infection. Infect Immun 70, 5596–5603 (2002).
Ito, H. et al. Lethal endotoxic shock using alpha-galactosylceramide sensitization as a new experimental model of septic shock. Lab Invest 86, 254–261 (2006).
Kishimoto, C., Kuribayashi, K., Masuda, T., Tomioka, N. & Kawai, C. Immunologic behavior of lymphocytes in experimental viral myocarditis: significance of T lymphocytes in the severity of myocarditis and silent myocarditis in BALB/c-nu/nu mice. Circulation 71, 1247–1254 (1985).
Topham, D. J., Adesina, A., Shenoy, M., Craighead, J. E. & Sriram, S. Indirect role of T cells in development of polioencephalitis and encephalomyelitis induced by encephalomyocarditis virus. J Virol 65, 3238–3245 (1991).
Wada, H. et al. Tumor necrosis factor-alpha (TNF-alpha) plays a protective role in acute viralmyocarditis in mice: A study using mice lacking TNF-alpha. Circulation 103, 743–749 (2001).
Yamamoto, K. et al. Attenuation of virus-induced myocardial injury by inhibition of the angiotensin II type 1 receptor signal and decreased nuclear factor-kappa B activation in knockout mice. J Am Coll Cardiol 42, 2000–2006 (2003).
Nasu-Nishimura, Y. et al. Cellular prion protein prevents brain damage after encephalomyocarditis virus infection in mice. Arch Virol 153, 1007–1012 (2008).
Ito, H. et al. Augmentation of lipopolysaccharide-induced nitric oxide production by alpha-galactosylceramide in mouse peritoneal cells. J Endotoxin Res 11,213–219 (2005).
Ohtaki, H. et al. Valpha14 NKT cells activated by alpha-galactosylceramide augment lipopolysaccharide-induced nitric oxide production in mouse intra-hepatic lymphocytes. Biochem Biophys Res Commun 378, 579–583 (2009).
Ohtaki, H. et al. Interaction between LPS-induced NO production and IDO activity in mouse peritoneal cells in the presence of activated Valpha14 NKT cells. Biochem Biophys Res Commun 389, 229–234 (2009).
Fejer, G. et al. Adenovirus infection dramatically augments lipopolysaccharide-induced TNF production and sensitizes to lethal shock. J Immunol 175, 1498–1506 (2005).
Nansen, A. & Randrup Thomsen, A. Viral infection causes rapid sensitization to lipopolysaccharide: central role of IFN-alpha beta. J Immunol 166, 982–988 (2001).
Matsushima, K., Larsen, C. G., DuBois, G. C. & Oppenheim, J. J. Purification and characterization of a novel monocyte chemotactic and activating factor produced by a human myelomonocytic cell line. J Exp Med 169, 1485–1490 (1989).
Jiang, Y., Beller, D. I., Frendl, G. & Graves, D. T. Monocyte chemoattractant protein-1 regulates adhesion molecule expression and cytokine production in human monocytes. J Immunol 148, 2423–2428 (1992).
Driscoll, K. E. Macrophage inflammatory proteins: biology and role in pulmonary inflammation. Exp Lung Res 20, 473–490 (1994).
Lira, S. A. et al. Expression of the chemokine N51/KC in the thymus and epidermis of transgenic mice results in marked infiltration of a single class of inflammatory cells. J Exp Med 180, 2039–2048 (1994).
Tracey, K. J. et al. Anti-cachectin/TNF monoclonal antibodies prevent septic shock during lethal bacteraemia. Nature 330, 662–664 (1987).
Sergerie, Y., Rivest, S. & Boivin, G. Tumor necrosis factor-alpha and interleukin-1 beta play a critical role in the resistance against lethal herpes simplex virus encephalitis. J Infect Dis 196, 853–860 (2007).
Bekker, L. G. et al. Immunopathologic effects of tumor necrosis factor alpha in murine mycobacterial infection are dose dependent. Infect Immun 68, 6954–6961 (2000).
Tchaptchet, S. et al. Innate, antigen-independent role for T cells in the activation of the immune system by Propionibacterium acnes. Eur J Immunol 40, 2506–2516 (2010).
Inatsu, A. et al. Novel mechanism of C-reactive protein for enhancing mouse liver innate immunity. Hepatology 49, 2044–2054 (2009).
Satoh, M. et al. Expression of Toll-like receptor 4 is associated with enteroviral replication in human myocarditis. Clin Sci (Lond) 104, 577–584 (2003).
Tang, H. et al. TLR4 activation is required for IL-17-induced multiple tissue inflammation and wasting in mice. J Immunol 185, 2563–2569 (2010).
Shiratsuchi, A., Kaido, M., Takizawa, T. & Nakanishi, Y. Phosphatidylserine-mediated phagocytosis of influenza A virus-infected cells by mouse peritoneal macrophages. J Virol 74, 9240–9244 (2000).
Narasaraju, T., Ng, H. H., Phoon, M. C. & Chow, V. T. MCP-1 antibody treatment enhances damage and impedes repair of the alveolar epithelium in influenza pneumonitis. Am J Respir Cell Mol Biol 42, 732–743 (2010).
Jung, I. D. et al. Blockade of indoleamine 2,3-dioxygenase protects mice against lipopolysaccharide-induced endotoxin shock. J Immunol 182, 3146–3154 (2009).
Liu, J. et al. The circadian clock Period 2 gene regulates gamma interferon production of NK cells in host response to lipopolysaccharide-induced endotoxic shock. Infect Immun 74, 4750–4756 (2006).
Morishima, T. et al. Encephalitis and encephalopathy associated with an influenza epidemic in Japan. Clin Infect Dis 35, 512–517 (2002).
Yajima, T. & Knowlton, K. U. Viral myocarditis: from the perspective of the virus. Circulation 119, 2615–2624 (2009).
Taniguchi, T., Takata, M., Ikeda, A., Momotani, E. & Sekikawa, K. Failure of germinal center formation and impairment of response to endotoxin in tumor necrosis factor alpha-deficient mice. Lab Invest 77, 647–658 (1997).
Ito, H. et al. Role of TNF-alpha produced by nonantigen-specific cells in a fulminant hepatitis mouse model. J Immunol 182, 391–397 (2009).
Pfister, O. et al. CD31- but Not CD31+ cardiac side population cells exhibit functional cardiomyogenic differentiation. Circ Res 97, 52–61 (2005).
Author information
Authors and Affiliations
Contributions
HO, HI and MS designed the project. HO, HI, YO, AH, TI, HM, KS and MS designed experiments. HO, MH and MT performed the experimental work. HO, HI and MS prepared the manuscript and figures. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareALike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article
Cite this article
Ohtaki, H., Ito, H., Hoshi, M. et al. High susceptibility to lipopolysaccharide-induced lethal shock in encephalomyocarditis virus-infected mice. Sci Rep 2, 367 (2012). https://doi.org/10.1038/srep00367
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep00367
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
