
Abstract
Metals like manganese (Mn) and iron (Fe) are essential for metabolism, while cadmium (Cd) is toxic for virtually all living organisms. Understanding the transport of these metals is important for breeding better crops. We have identified that OsNRAMP5 contributes to Mn, Fe and Cd transport in rice. OsNRAMP5 expression was restricted to roots epidermis, exodermis and outer layers of the cortex as well as in tissues around the xylem. OsNRAMP5 localized to the plasma membrane and complemented the growth of yeast strains defective in Mn, Fe and Cd transport. OsNRAMP5 RNAi (OsNRAMP5i) plants accumulated less Mn in the roots and less Mn and Fe in shoots and xylem sap. The suppression of OsNRAMP5 promoted Cd translocation to shoots, highlighting the importance of this gene for Cd phytoremediation. These data reveal that OsNRAMP5 contributes to Mn, Cd and Fe transport in rice and is important for plant growth and development.
Similar content being viewed by others


Introduction
Manganese (Mn) and iron (Fe) are essential micronutrients for almost all living organisms including plants. Mn is required as a cofactor or activator for enzymes belonging to different functional groups which perform diverse functions. For example, oxalate oxidase, Mn superoxide dismutase (MnSOD), RNA polymerase, malic enzyme, isocitrate dehydrogenase and PEP carboxykinase use Mn as a cofactor1,2,3,4. Mn is also important in photosynthetic oxygen evolution in chloroplasts. The oxygen evolving complex, which is a part of photosystem II in the thylakoid membrane of chloroplasts, is responsible for the terminal photooxidation of water during the light reactions of photosynthesis and it has a metalloenzyme core containing four Mn atoms5,6,7. Mn deficiency causes interveinal chlorosis and a reduction in the content of fructans and structural carbohydrates, resulting in lax leaves8. Consequently, Mn-deficient plants are more susceptible to low-temperature stress and pathogen infection, leading to significant decreases in crop yield1,9. Similarly, Fe servers as an integral cofactor for numerous proteins. In plants, Fe is essential for chlorophyll biosynthesis and the synthesis of heme and its deficiency significantly impairs plant growth and development10,11.
The transport of Mn is believed to share an entry route with Fe and cadmium (Cd). Fe is taken up by IRT1, a member of the Fe-regulated transporter-like protein (ZIP) gene family12,13,14,15. IRT1 has a relatively broad substrate spectrum and is reportedly capable of transporting other metal ions, including Mn and Cd16. Natural resistance associated macrophage protein (NRAMP) family members, which have been identified in many plant species, play a major role in metal homeostasis. In Arabidopsis, it was recently reported that AtNRAMP1 is essential for Mn uptake under Mn deficiency17. In addition to Mn, AtNRAMP1 transports Fe and is induced by Fe deficiency18. AtNRAMP6, a homolog of AtNRAMP1, does not transport Mn and Fe and seems to affect the intracellular distribution of Cd19.
In rice plants, little is known about Mn transport at the molecular level and the transporter for Mn uptake from soil has not been reported yet. The only reported rice Mn transporter is OsYSL2, which transports Mn- and Fe-nicotianamine (NA) complexes and contributes to phloem translocation20,21. On the other hand, Fe uptake systems in rice plants are well known as Strategy II and direct ferrous Fe uptake systems. Strategy II Fe uptake systems rely on a ferric Fe chelation system through the secretion of deoxymugineic acid (DMA)10,11,22,23. The secreted DMA chelate Fe and the resulting Fe(III)-DMA complex is transported by OsYSL1523,24. Besides using Strategy II systems, rice plants possess a direct ferrous Fe uptake system that uses the ferrous Fe transporter OsIRT125. Recently we have reported that rice also secretes phenolics to increase solubility of apoplasmic Fe26,27,28. All genes related to Fe uptake are highly induced by Fe deficiency. Thus many of rice genes have the potential to complement the function of other genes for Fe transport.
Unlike Fe and Mn, Cd is a highly toxic metal for most living organisms, including humans29. Even at low concentrations, cumulative uptake of Cd dramatically affects cellular metabolism30. The effects of daily intake of Cd-contaminated rice were seen in Japanese people during the 1910–1970s, resulting in a disease termed “itai-itai”31,32. Cd contamination in crops is spreading all over the world, especially in countries with high industrial activity33. In a globalized market, Cd-contaminated food threatens not only the production areas, but also consumers living far from contaminated fields. Phytoremediation, which utilizes plants to extract soil Cd, is a promising method to decontaminate soils high in Cd. This method is cost-effective, applicable to large areas and environmentally friendly34. Rice is a good candidate for Cd phytoremediation35 due to its adaptation to a range in environmental conditions, such as flooded and drained fields. Moreover, rice has a short life cycle, a large biomass and well-established cultivation and harvesting methods. Phytoremediation using a naturally high Cd-accumulating rice cultivar for two years showed a 38% reduction in soil Cd content and a 51% reduction in grain Cd content of successive rice crops36,37. Anjana Dhan is one of the highest Cd accumulating natural rice cultivars among world rice collections38, raising the challenge to determine if its Cd accumulation capacity could be further enhanced.
Here, we show that OsNRAMP5 is a Mn, Fe and Cd transporter in rice plants. OsNRAMP5 is not regulated by Mn or Fe deficiency. The repression of OsNRAMP5 expression via RNAi lowered root and shoot Mn accumulation. In OsNRAMP5i plants, the Fe concentration was unchanged in roots, but was low in the xylem sap and shoots and the genes related to Fe uptake and translocation were induced. OsNRAMP5 knockdown plants accumulated more Cd in shoots, making it a promising candidate for Cd phytoremediation.
Results
Identification of OsNRAMP5
Rice plants grown hydroponically in a solution containing 10 µM Cd accumulated less Mn, zinc and copper in their roots and shoots, while Fe accumulation decreased in the shoots and increased in the roots ( Fig. 1 ). Microarray analyses were performed to identify the genes responsible for the changes in metal accumulation. Our results indicate that the ferrous Fe and Cd transporters OsIRT2 and OsNRAMP1, the ferrous Fe- and Mn-NA transporter OsYSL2 and zinc transporter OsZIP3 were induced in the roots of plants exposed to Cd, while OsIRT2, OsZIP2, OsZIP3, OsZIP8 and the ferric Fe-DMA transporter OsYSL15 were induced in the shoots ( Supplementary Table 1 )21,24,25,39,40. Among the seven members of the NRAMP family in rice, OsNRAMP140 expression was increased in roots and shoots in the presence of Cd, while OsNRAMP5 (Os07g0257200) expression was decreased in both of these tissues ( Supplementary Table 1 ). The root and shoot expression of other OsNRAMP genes did not change significantly. We next characterized OsNRAMP5 to determine its role in Mn, Cd and Fe transport. OsNRAMP5 shares 74.5% homology with OsNRAMP1 and the expression of OsNRAMP5 did not change in response to Mn or Fe deficiency ( Fig. 2a and b ), while the expression of OsNRAMP5 decreased in response to the addition of Cd ( Fig. 2c ). In contrast, OsNRAMP1 expression was upregulated by Mn deficiency in roots and shoots ( Supplementary Fig. 1a and b ).

Metal concentrations of rice plants grown in the presence of Cd.
Shoot and root metal concentrations under control conditions (Ctrl.) or in the presence of 10 µM Cd (+Cd). (a, e) Mn, (b, f) Fe, (c, g) zinc and (d, h) copper concentrations. Error bars represent the SD. Columns bars followed by different letters are statistically different according to analysis of variance followed by Student-Newman-Keuls (SNK) test (a, p = 0.0034; b, p = 0.0074; c, p = 0.0000; d, p = 0.0000; e, p = 0.0017; f, p = 0.0069; g, p = 0.0005; h, p = 0.0010); n = 3.

OsNRAMP5 is expressed in rice roots.
(a–c) Expression pattern of OsNRAMP5 in rice grown hydroponically under Mn-deficient (-Mn) (a), Fe-deficient conditions (-Fe) (b) and in the presence of 10 µM Cd (+Cd) (c) compared to normal nutrient conditions (Ctrl.). (d–f) OsNRAMP5 promoter GUS analysis in rice roots. Longitudinal section (d). Transverse section (e). Enlargement of part of the stele (f). MXII, metaxylem II; MXI, metaxylem I; PX, protoxylem. Scale bar = 400 μm for (d) and 50 μm for (f). Error bars represent the SD. Columns bars followed by different letters are statistically different according to analysis of variance followed by SNK test (p = 0.0034); n = 3.
OsNRAMP5 is expressed in the root epidermis, exodermis and outer layers of the cortex
To obtain greater insight into the physiological role of OsNRAMP5, we investigated the tissue-specific localization of OsNRAMP5 in roots through an analysis of promoter-β-glucuronidase (GUS) transgenic rice. GUS staining attributed to OsNRAMP5 promoter activity was observed in the epidermis, exodermis and outer layers of the cortex ( Fig. 2d and e ). In the stele, GUS activity was detected mainly around the xylem ( Fig. 2f ).
OsNRAMP5 is a plasma membrane-localized Mn, Fe and Cd transporter
OsNRAMP5 fused to the N terminus of synthetic green fluorescent protein (sGFP) was transiently expressed in onion epidermal cells under control of the cauliflower mosaic virus (CaMV) 35S promoter. OsNRAMP5-sGFP fluorescence was observed at the plasma membrane ( Fig. 3a ). To further confirm the localization, onion cells transformed with OsNRAMP5-sGFP were stained with plasma membrane dye FM4-64 ( Fig. 3b ) and the merged images clearly showed that OsNRAMP5 localizes to plasma membrane ( Fig. 3c ). On the other hand sGFP alone was localized to the cytoplasm and nucleus ( Fig. 3d ). The OsNRAMP5 complemented the growth of yeast strains Δsmf141 and Δfet3Δfet442, which are defective in Mn and Fe transport, respectively ( Fig. 3e and f ). In addition to Mn and Fe, OsNRAMP5 was shown to affect the growth of yeast strain Δycf1 ( Fig. 3g ), which is defective in Cd sequestration to the vacuole and which exhibits slower growth in the presence of Cd43; however, OsNRAMP5 did not restore the growth of any of the yeast zinc and copper transport mutants ( Supplementary Fig. 2 ). OsNRAMP1 used as a control transported Fe and Cd but not Mn in line with the previous reports18,40. The results clearly showed that OsNRAMP5 is localized to the plasma membrane and involved in Mn, Fe and Cd transport. Moreover the expression around xylem indicated that OsNRAMP5 may also contribute to Mn, Fe and Cd uptake from xylem.

OsNRAMP5 contributes to Mn, Fe and Cd transport.
(a–d) Green fluorescent protein (GFP) fluorescence in onion epidermal cells observed by confocal laser scanning microscopy (a), OsNRAMP5-sGFP fluorescence (b), fluorescence of FM4-64 and (c) overlay of FM4-64 and OsNRAMP5-sGFP. (d) Fluorescence of sGFP only. Scale bar = 50 μm. (e–g) Serial dilutions of yeast cells for Δsmf (Mn uptake mutant) (e), Δfet3fet4 (Fe uptake mutant) (f), Δycf1 (Cd-sensitive mutant) (g), transformed with empty vector (V.C.), OsNRAMP1 or OsNRAMP5.
OsNRAMP1 does not contribute to Mn uptake
AtNRAMP1 transports Fe and Mn and is upregulated by Fe and Mn deficiency17,18. OsNRAMP1 is induced by Fe deficiency and transports Fe, but not Mn18,40. To analyze the relationship between OsNRAMP1 and Mn uptake in rice plants, we developed and analyzed OsNRAMP1 RNAi (OsNRAMP1i) plants. In OsNRAMP1i plants, OsNRAMP1 expression was significantly downregulated ( Supplementary Fig. 1c and d ); however, there was no significant difference in Mn or Fe concentration between OsNRAMP1i and wild type plants ( Supplementary Fig. 3 ).
OsNRAMP5 RNAi and overexpression (OX) plants
To understand the role of OsNRAMP5 in rice plants (cultivar Tsukinohikari), RNAi knockdown plants (T5i) and OX plants of OsNRAMP5 were developed. The OsNRAMP5 transcript levels in T5i roots were five times lower than those in wild type, while the expression of OsNRAMP5 in the OX lines was comparable to that in wild type plants ( Fig. 4a ). In shoots, OsNRAMP5 expression was observed only in the OX lines ( Fig. 4b ). The growth of the T5i lines was comparable to that of wild type plants in terms of root and shoot length and SPAD reading under control conditions ( Fig. 4c and e–g ). Shoot length in the OX lines was significantly reduced compared to that in wild type plants under control conditions ( Fig. 4f ). The root and shoot Mn concentrations in the T5i plants were dramatically reduced compared to those in wild type plants ( Fig. 4h and i ). There was no significant difference in the concentration of Fe in roots ( Fig. 4j ) while the Fe concentration was significantly lower in shoots of T5i plants ( Fig. 4k ). Microarray analysis revealed that many Fe deficiency-inducible genes were upregulated in the T5i plants ( Supplementary Table 2 ).

Characterization of OsNRAMP5i and OsNRAMP5 OX lines.
(a, b) OsNRAMP5 transcription in roots (a) and shoots (b) of wild type, OsNRAMP5i (T5i-1, T5i-2) and OsNRAMP5OX (OX1, OX2) plants (cultivar Tsukinohikari) grown in control hydroponic culture. (c, d) Phenotypes of wild type and T5i plants grown under control (c) and Mn-deficient conditions (d). Scale bars = 10 cm. Root length (e), shoot length (f), SPAD value (g), root (h) and shoot (i) Mn concentrations, root (j) and shoot (k) Fe concentrations of plant grown under control conditions or Mn-deficient (-Mn) conditions. Error bars represent the SD. Columns bars followed by different letters are statistically different according to analysis of variance followed by SNK test (a, p = 0.0013; b, p = 0.0003; e, p = 0.0017; f, p = 0.0009 (Ctrl.) & p = 0.0006 (-Mn); g, p = 0.0001; h, p = 0.0000 (Ctrl.) & p = 0.0067 (-Mn); i, p = 0.0000 (Ctrl.) & p = 0.0004 (-Mn); j, p = 0.0095; k, p = 0.0000; n = 3.
In plants grown under Mn-deficient conditions, the root and shoot length and SPAD value of the T5i lines were significantly decreased ( Fig. 4d–g ). The growth of the OX lines was also affected in terms of shoot length and SPAD value ( Fig. 4f and g ). The T5i lines accumulated less Mn than wild type plants both in roots and shoots ( Fig. 4h and i ), while the Mn concentration in the OX plants was similar to that in wild type roots ( Fig. 4h ). The Mn and Fe concentrations in xylem sap from the T5i lines were also significantly reduced compared to those in wild type plants ( Fig. 5a and b ), whereas no difference was observed in the Ca concentration ( Fig. 5c ). Under Fe-deficient conditions, there were no significant differences in plant growth, SPAD value and metal concentrations ( Supplementary Fig. 4 ).

Xylem sap metal concentrations in OsNRAMP5i plants.
Xylem Mn (a), Fe (b) and Ca (c) concentrations in T5i plants grown under control or Mn-deficient (-Mn) conditions. Error bars represent the SD. Columns bars followed by different letters are statistically different according to analysis of variance followed by SNK test (a, p = 0.0001 (Ctrl.) & p = 0.0010 (-Mn); b, p = 0.0000 (Ctrl.) & p = 0.0001 (-Mn); n = 3.
The Mn concentration in T5i plants grown in the presence of 10 µM Cd was significantly lower in shoot and roots compared with wild type ( Fig. 6a and e ). There was no difference in Fe concentration between the T5i and wild type plants ( Fig. 6b and f ). The Cd concentration and total Cd content in the shoots were higher in the T5i plants than in wild type ( Fig. 6c and d ). On the other hand, the root Cd concentration and total Cd content of roots in the T5i plants were less than those in wild type ( Fig. 6g and h ). The total Cd content (root plus shoot) in the T5i plants was also significantly less than that in wild type plants.

Suppression of OsNRAMP5 leads to high Cd in rice shoots.
Shoot (a) and root (e) Mn; shoot (b) and root (f) Fe; shoot (c) and root (g) Cd concentrations and shoot (d) and root (h) Cd content in T5i and OX plants grown under 10 µM Cd conditions. Error bars represent the SD. Columns bars followed by different letters are statistically different according to analysis of variance followed by SNK test (a, p = 0.0004; c, p = 0.0005; d, p = 0.0001; e, p = 0.0067; g, p = 0.0017; h, p = 0.0001); n = 3.
Development of high Cd accumulating Anjana Dhan lines
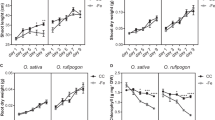
As the OsNRAMP5i (T5i) shoots accumulated more Cd than did wild type shoots, we developed OsNRAMP5i plants (A5i) using a high Cd accumulating cultivar, Anjana Dhan38. There was no significant difference in shoot length between the Anjana Dhan OsNRAMP5i (A5i) and wild type plants when plants were grown in the presence of 10 µM Cd ( Fig. 7a ). OsNRAMP5 expression in the A5i roots was 33–66% less than that in wild type when grown under normal nutrient conditions ( Fig. 7b ). At 10 µM Cd conditions, slightly less Mn accumulated in the A5i shoots and there was no difference in Fe concentration ( Fig. 7c and d ). Moreover, there was no significant difference in shoot weight; however, the shoot Cd concentration in A5i was up to 4.9 times that in wild type ( Fig. 7e and f ) and the shoot Cd content in A5i was up to 4.3 times greater than that in wild type ( Fig. 7g ).

OsNRAMP5i Anjana Dhan for Cd phytoremediation.
(a) Phenotypes of four-week-old wild type and OsNRAMP5i (A5i-2) Anjana Dhan plants transferred to a nutrient solution containing 10 µM CdCl2 and cultivated for two weeks. Scale bars = 10 cm. (b) OsNRAMP5 transcripts in the roots of wild type and OsNRAMP5i (A5i-1, A5i-2, A5i-3, A5i-4) Anjana Dhan plants grown in normal nutrient hydroponic culture. Shoot Mn (c), Fe (d) concentration, shoot dry weight (e), shoot Cd concentration (f) and Shoot Cd content (g) of wild type and A5i plants grown under 10 μM Cd condition. Error bars represent the SD. Columns bars followed by different letters are statistically different according to analysis of variance followed by SNK test (b, p0.0000; e, p = 0.0113; f, p = 0.0000; g, p = 0.0000); n = 3.
Discussion
In rice plants, the mechanisms of Mn uptake have not been elucidated. Until now only OsYSL2, a Mn- and Fe-NA transporter, has been reported to be involved in phloem Mn transport20,21 while the gene involved in Mn uptake from soil has not been characterized yet. Like OsYSL2, numerous transporters can transport more than one metal; for example, AtNRAMP1 transports Mn and Fe17,18,44, while OsNRAMP1 transports Fe and Cd18,40. Thus, many metals compete with each other and an abundance of one metal can disturb the uptake of others. In this scenario, it was not surprising that Cd showed a negative correlation with Mn absorption and accumulation in shoots. We have previously reported 22K microarray analysis using different concentrations of Cd and observed more significant changes in gene expression for 10 µM Cd treatment compared with 1 µM Cd treatment45. In the present study, when plants were grown in the presence of 10 µM Cd, in addition to decreased Mn accumulation in rice plants, changes in the expression of numerous metal transporters were observed. We focused on the role of OsNRAMP5 in Mn, Fe and Cd transport. NRAMP family members are involved in Mn transport in other organisms17,46,47. Although a role for rice NRAMPs in Mn transport was previously suggested48, the involvement of OsNRAMP family proteins in Mn transport has not been investigated. Among the seven members of the NRAMP family in rice, the expression of OsNRAMP1 increased in roots and shoots in the presence of Cd, while the expression of OsNRAMP5 decreased in both tissues ( Supplementary Table 1 ). The expression of other OsNRAMP family genes did not change significantly in roots and shoots. The decreased Mn content associated with reduced OsNRAMP5 expression suggests that gene's involved in Mn transport and our data confirm that OsNRAMP5 plays a role in Mn transport in rice plants.
The Arabidopsis, AtNRAMP1 transports Mn, in addition to Fe17,18,44. AtNRAMP1 is upregulated by Mn and Fe deficiency, indicating that AtNRAMP1 is a multi-functional transporter involved in Mn and Fe uptake. On the other hand, AtNRAMP6, the closest homolog of AtNRAMP1, does not transport Mn19. In rice plants, although the expression of OsNRAMP1 was upregulated by Fe and Mn deficiency, it does not transport Mn ( Fig. 3e ). Moreover, no difference in Mn concentration was observed between OsNRAMP1i and wild type plants ( Supplementary Fig. 3 ). On the other hand, OsNRAMP5 contributes to Mn and Fe transport, although its expression is not upregulated by Mn or Fe deficiency ( Fig. 2 ). The growth of OsNRAMP5i was comparable to that of wild type plants under normal conditions; however, when these lines were grown under Mn-deficient conditions, plant growth was significantly reduced ( Fig. 4 ). The phenotype of AtNRAMP1 also becomes prominent under Mn-deficient conditions17. The reduced Mn concentration in T5i plants strongly suggested that OsNRAMP5 contributes to Mn transport in rice.
Under normal growth conditions in rice, OsNRAMP1, OsIRT1 and OsYSL15 are expressed at low levels and are highly induced by Fe deficiency24,25,40. On the other hand, OsNRAMP5 is a constitutively expressed transporter ( Fig. 2 )49. OsNRAMP5 contributes to Fe transport in addition to Mn and Cd, as revealed by yeast complement assay ( Fig. 3e–g ). In T5i plants, the Fe concentration did not change in roots however significant changes were observed for Fe concentration in the xylem sap as well as in shoot ( Fig. 4j, k and Fig. 5b ). The expression of genes involved in Fe uptake and translocation were highly induced in T5i plants ( Supplementary Table 2 ). The expression of NA and DMA synthesis related genes and Fe(II)/Mn(II)-NA and Fe(III)-DMA transporter genes was also upregulated. The up-regulations of these genes suggest that the decrease in Fe uptake by the suppression of OsNRAMP5 would be complemented by an increase in Fe uptake from soil through these genes, however the root to shoot Fe transport via root xylem may not be complemented by increased expression of these genes. These data also suggest that OsNRAMP5 is involved in constitutive uptake of Fe and that OsNRAMP1, OsIRT1 and OsYSL15 play a significant role under Fe deficiency. In rice plants, OsNRAMP5 would function jointly in Fe uptake with these transporters.
Initially, we expected that the root and shoot Cd concentration in the T5i plants would be low, as OsNRAMP5 contributed to Cd transport in yeast complementation assay ( Fig. 3g ). Unexpectedly, T5i plants accumulated small amounts of Cd in the roots but large amounts in the shoots ( Fig. 6c and g ). The total Cd content (root plus shoot) in the OsNRAMP5i plants was less than that in wild type, suggesting that OsNRAMP5 did not contribute to Cd translocation but to the uptake of Cd from the soil ( Fig. 6d and h ). The expression observed near xylem indicated the OsNRAMP5 may also be involved in Cd uptake from xylem. The role of OsHMA3 in sequestering Cd to vacuole has already been discussed50. It may be possible that in rice the Cd taken from xylem may be further transported into the vacuole through OsHMA3 and OsHMA3 and OsNRAMP5 may function in coordination. Cd uptake and translocation is partly mediated by the Fe uptake system in rice plants40,51. Induction of the genes related to Fe transport enhanced Cd translocation to the shoots in T5i plants ( Supplementary Table 2 ). Expression of these genes in roots is higher in the high Cd accumulating cultivar Habataki than in the low Cd accumulating cultivar Sasanishiki40. Such a difference in Fe transporter expression seems to be responsible for the differences in shoot Cd accumulation between the two rice cultivars. We previously developed rice lines tolerant to a low Fe supply and with greater seed Fe content by regulating the spatial and temporal expression of different genes21,26,52. This led us to expect that shoot Cd translocation in rice plants could be increased by the suppression of OsNRAMP5. Indeed, this was achieved in Anjana Dhan, a natural rice cultivar that accumulates a large amount of Cd without any adverse effects on plant growth. The A5i lines accumulated up to 4.9 times more Cd than wild type. Moreover, the shoot Cd content in A5i was up to 4.3 times higher than in wild type ( Fig. 7 ). Cd phytoremediation using this efficient transgenic rice will rapidly and efficiently enable the removal of Cd from soil, leading to at least a 75% reduction in the Cd phytoremediation period53. This could contribute greatly to food safety and human health worldwide.
In rice plants, net photosynthesis and photosynthetic capacity correlate with the applied dose of Mn and the optimal Mn concentration in hydroponic solutions is around 0.5–2 mg/L54. In this range, the Mn concentration in rice shoots is 350–896 µg/g dry weight. The Mn concentration is normally greater than the optimal concentration for rice in normal and acidic soils54. Therefore, controlling the expression of OsNRAMP5 may enhance photosynthesis in rice plants. Moreover, OsNRAMP5 is closely related to Fe and Cd uptake and translocation; thus, it may control the Cd concentration in rice plants. The identification of a transporter involved in constitutive Mn and Fe transport will increase both our understanding of Mn and Fe transport and of the mechanisms involved in Mn and Fe homeostasis in rice plants.
Methods
Plant material
Seeds of the Oryza sativa cultivars Tsukinohikari and Anjana Dhan were germinated for two weeks on Murashige and Skoog (MS) medium at 28°C under a 16-h light/8-h dark photoperiod. After germination, the seedlings were transferred to a 20-L plastic container containing a standard nutrient solution as described previously55. For the Cd stress treatments, four-week-old plants were transferred to a nutrient solution containing 10 µM CdCl2 as described previously45 and cultivated for two weeks. For the Mn-deficient conditions, four-week-old plants were transferred to a nutrient solution without Mn and grown for two additional weeks. For the Fe-deficient conditions, four-week-old plants were transferred to a nutrient solution without Fe and grown for an additional week. For the soil growth conditions, 8 ppm CdCl2 was added to 500 g of bonsol (Sumitomo Chem. Co., Osaka, Japan). Two-week-old seedlings were transplanted to soil and grown in a greenhouse under natural light conditions.
Microarray and RT-PCR
Microarray analyses were done using a rice 44K custom oligo DNA microarray kit (Agilent Technology, Tokyo, Japan) as described previously55,56. The reproducibility of the analysis was assessed by a dye swap with two biological replicates ( Supplementary Table 1 ) or three biological replicates for the T5i experiment ( Supplementary Table 2 ). qRT-PCR was performed as described previously55 using the primers OsNRAMP5 RT forward 5′-CCGCCGTCGTCGACGCCGAGAAGGC-3′ and OsNRAMP5 RT reverse 5′-ACACCCTTGTCGATCGATCGATCTG-3′. The α-tubulin primers used for qRT-PCR were α-tubulin forward 5′-TCTTCCACCCTGAGCAGCTC-3′ and α-tubulin reverse 5′-AACCTTGGAGACCAGTGCAG-3′. All experiments were performed in triplicate.
Determination of metal concentrations
Roots samples were washed with de-ionized water thrice before sampling. Leaf and root samples were dried for one week at 70°C and were digested with 3 ml of 13 M HNO3 at 220°C for 20 min using a MARS XPRESS microwave reaction system (CEM, Matthews, NC, USA) in triplicate. For xylem sap collection, rice plants were de-topped at a height of 3 cm from the root. The surface of the excised leaf sheath was gently wiped and a tube filled with cotton was placed on the cut end. The entire length of the tube was covered with aluminum foil and the xylem sap was collected for 30 min. Forty-five rice plants were used for xylem sap collection. The xylem sap was collected in cotton and filtered with a 0.45 μm filtering column (Millipore). The extracted xylem sap was stored at −20°C until further analysis. Because the amount of xylem sap differed, 36 samples with median values of these xylem sap amounts were selected and twelve individual samples each were pooled at random into three tubes for further analysis. Xylem samples were digested with 3 ml of 13 M HNO3 at 80°C for 60 min. After digestion, samples were collected, diluted to 5 ml and analyzed by ICP-AES (SPS1200VR; Seiko, Tokyo, Japan) as described previously26,52.
Subcellular localization of OsNRAMP5
The full-length ORF of OsNRAMP5 was amplified by PCR with the forward primer 5′-CACCATGGAGATTGAGAGAGAGAGCAGTG-3′ and reverse 5′-ACACCCTTGTCGATCGATCGATCTG-3′ primers and subcloned into pH7WGF257. Onion epidermal cells were transformed, stained with plasma membrane dye FM4-64 (Invitrogen, Tokyo, Japan) and observed with a LSM5 Pascal laser-scanning confocal microscope (Carl Zeiss, Tokyo, Japan) as described previously25,55.
Yeast strains and growth media
The OsNRAMP5 ORF was cloned into the expression vector pDR19558 and then introduced into yeast cells using the lithium acetate method. The following strains of Saccharomyces cerevisiae were used: wild type parental strain (BY4741; MATalpha, his3Δ1; leu2Δ0; met15Δ0; ura3Δ0), ferrous Fe uptake-defective double mutant Δfet3fet4 (MATalpha, his3Δ1; leu2Δ0; met15Δ0; ura3Δ0; YMR058w::kanMX4; YMR319c::kanMX4), Mn uptake-defective mutant Δsmf1 (MATalpha, his3Δ1; leu2Δ0; met15Δ0; ura3Δ0; YOL122c::kanMX4) and Cd-sensitive mutant Δycf1 (MATalpha, Δtrp1, Δhis3, Δleu2, Δura3; Δycf::TRY1). ycf1 lacks the function for compartmentalization of Cd into vacuoles43. For media without Fe or Mn, 10 μM bathophenanthroline disulfonic acid (BPDS) or 20 mM ethylene glycol-bis-β-aminoethylether-N,N,N',N'-tetraacetic acid (EGTA) with 50 mM 2-morpholinoethanesulfonic acid, monohydrate (MES) were added, respectively (Wako Pure Chemical Industries). For Cd treatment, 10 μM CdCl2 was added. After spotting at three dilutions (optical densities at 600 nm of 0.1, 0.01 and 0.001), the plates were incubated at 30°C for five days.
Rice transformation
The 0.8-kb 5′-flanking region of OsNRAMP5 was amplified by PCR using genomic DNA as template with the forward primer 5′-TTCCCCAAGCTTACACAAGAGGTTTTGCACACAGG -3′ and reverse primer 5′-CTCAATTCTAGATGTTGCTTCCTCTCTTAGCTTCT-3′, which contained HindIII and XbaI restriction sites, respectively. The amplified fragment was fused into pBluescript II SK+ and its sequence confirmed. The OsNRAMP5 promoter was digested with HindIII and XbaI and the digested 0.8-kb fragment was subcloned upstream of the uidA ORF, which encodes β-glucuronidase, in pIG121Hm. Oryza sativa L. cv. Tsukinohikari was transformed and the histochemical localization of OsNRAMP5 was observed in three independent T2 plants as described previously25.
The plasmid pH7FWG2-OsNRAMP5 was used to overexpress OsNRAMP5. To suppress OsNRAMP1 expression, a 300-bp fragment from the 3′-UTR region of OsNRAMP1 was amplified by PCR with the forward primer 5′-CACCTACCCTTCCAGACCGGTTACC-3′ and reverse primer 5′-GATTGTCCTTGTCCTCGCATC-3′. To suppress OsNRAMP5 expression, a 300-bp fragment from the 3′-UTR region of OsNRAMP5 was amplified by PCR with the forward primer 5′-GCAGTAAGAGCAAGATGGGGCCCCAC-3′ and reverse primer 5′-TTGGGAGCGGGATGTCGGCCAGGT-3′. The amplified fragment was cloned into pIG121-RNAi-DEST21 and used for transformation.
SPAD reading
The degree of chlorosis in the youngest fully expanded leaf was determined using a SPAD-502 chlorophyll meter (Minolta Co., Tokyo, Japan).
Statistical analysis
To determine whether the observed differences between the WT and OsNRAMP5i and OX lines were statistically significant, one-way ANOVA was performed using Costat 6.400 (Cohort, USA) with a completely randomized design followed by a Student-Newman-Keuls (SNK) test (P<0.05).
References
Marschner, H. Mineral nutrition of higher plants. 2nd edn, (Academic Press, 1995).
Bowler, C. et al. Manganese superoxide dismutase can reduce cellular damage mediated by oxygen radicals in transgenic plants. EMBO J. 10, 1723–1732 (1991).
Requena, L. & Bornemann, S. Barley (Hordeum vulgare) oxalate oxidase is a manganese-containing enzyme. Biochem. J. 343, 185–190 (1999).
Alscher, R. G., Erturk, N. & Heath, L. S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 53, 1331–1341 (2002).
Britt, R. D. Oxygen evolution. in Oxygenic photosynthesis: the light reactions. Vol. Advances in Photosynthesis and Respiration. 4, 137–159 (Kluwer Academic Publishers 1996).
Clemens, K. L., Force, D. A. & Britt, R. D. Acetate binding at the photosystem II cxygen evolving complex: an S2-state multiline signal ESEEM study. J. American Chem. Society 124, 10921–10933 (2002).
Rutherford, A. W. & Boussac, A. Water photolysis in biology. Science 303, 1782–1784 (2004).
Pearson, J. N. & Rengel, Z. Genotypic differences in the production and partitioning of carbohydrates between roots and shoots of wheat grown under zinc or manganese deficiency. Annals of Botany 80, 803–808 (1997).
Hebbern, C. A., Pedas, P., Schjoerring, J. K., Knudsen, L. & Husted, S. Genotypic differences in manganese efficiency: field experiments with winter barley (Hordeum vulgare L.). Plant and Soil 272, 233–244, 10.1007/s11104-004-5048-9 (2005).
Bashir, K. et al. Cloning and characterization of deoxymugineic acid synthase genes from graminaceous plants. J. Biol. Chem. 281, 32395–32402 (2006).
Bashir, K., Ishimaru, Y. & Nishizawa, N. K. Iron uptake and loading into rice grains. Rice 3, 122–130 (2010).
Eide, D., Broderius, M., Fett, J. & Guerinot, M. L. A novel iron-regulated metal transporter from plants identified by functional expression in yeast. Proc. Natl. Acad. Sci. USA 93, 5624–5628 (1996).
Henriques, R. et al. Knock-out of Arabidopsis metal transporter gene IRT1 results in iron deficiency accompanied by cell differentiation defects. Plant Mol. Biol. 50, 587–597 (2002).
Varotto, C. et al. The metal ion transporter IRT1 is necessary for iron homeostasis and efficient photosynthesis in Arabidopsis thaliana. Plant J. 31, 589–599 (2002).
Vert, G. et al. IRT1, an Arabidopsis transporter essential for iron uptake from the soil and for plant growth. The Plant Cell 14, 1223–1233 (2002).
Vert, G., Briat, J.-F. & Curie, C. Arabidopsis IRT2 gene encodes a root-periphery iron transporter. Plant J. 26, 181–189 (2001).
Cailliatte, R., Schikora, A., Briat, J.-F., Mari, S. & Curie, C. High-affinity manganese uptake by the metal transporter NRAMP1 is essential for arabidopsis growth in low manganese conditions. The Plant Cell 22, 904–917 (2010).
Curie, C., Alonso, J. M., Le Jean, M., Ecker, J. R. & Briat, J. F. Involvement of NRAMP1 from Arabidopsis thaliana in iron transport. Biochem. J. 347, 749–755 (2000).
Cailliatte, R., Lapeyre, B., Briat, J. F., Mari, S. & Curie, C. The NRAMP6 metal transporter contributes to cadmium toxicity. Biochem. J. 422, 217–228 (2009).
Koike, S. et al. OsYSL2 is a rice metal-nicotianamine transporter that is regulated by iron and expressed in the phloem. Plant J. 39, 415–424 (2004).
Ishimaru, Y. et al. Rice metal-nicotianamine transporter, OsYSL2, is required for the long-distance transport of iron and manganese. Plant J. 62, 379–390 (2010).
Bashir, K. & Nishizawa, N. K. Deoxymugineic acid synthase: A gene important for Fe-acquisition and homeostasis. Plant signaling & behavior 1, 292 (2006).
Nozoye, T. et al. Phytosiderophore efflux transporters are crucial for iron acquisition in graminaceous plants. J. Biol. Chem. 286, 5446–5454 (2011).
Inoue, H. et al. Rice OsYSL15 Is an iron-regulated iron(III)-deoxymugineic acid transporter expressed in the roots and is essential for iron uptake in early growth of the seedlings. J. Biol. Chem. 284, 3470–3479 (2009).
Ishimaru, Y. et al. Rice plants take up iron as an Fe3+-phytosiderophore and as Fe2+. Plant J. 45, 335–346 (2006).
Ishimaru, Y. et al. A rice phenolic efflux transporter is essential for solubilizing precipitated apoplasmic iron in the plant stele. J. Biol. Chem. 286, 24649–24655 (2011).
Ishimaru, Y., Bashir, K., Nakanishi, H. & Nishizawa, N. K. The role of rice phenolics efflux transporter in solubilizing apoplasmic iron. Plant signaling & behavior 6, 1624–1626 (2011).
Bashir, K. et al. Rice phenolics efflux transporter 2 (PEZ2) plays an important role in solubilizing apoplasmic iron. Soil Sci. Plant Nutr. 57, 803–812 (2011).
World Health Organization. Summary and conclusion of the sixty-first meeting of the Joint FAO/WHO Expert Committee on Food Additives. .16–18 (World Health Organization, 2003).
Singh, B. R. </author> & McLaughlin, M. J. Cadmium in Soils and Plants (eds M. J. McLaughlin, & B. R. Singh) 257–267 (Kluwer Academic Publishing, 1999).
Kobayashi, J. in Toxicity of Heavy Metals in the Environment (ed F. W. Oeheme) 199–260 (Marcel Dekker, 1978).
Horiguchi, H. et al. Latest status of cadmium accumulation and its effects on kidneys, bone and erythropoiesis in inhabitants of the formerly cadmium-polluted jinzu river basin in toyama, japan, after restoration of rice paddies. Intern. Archives Occup & Environ. Health 83, 953–970 (2010).
Fishbein, F. Sources, transport and alterations of metal compounds: an overview. I. Arsenic, beryllium, cadmium, chromium and nickel. Environ. Health Perspect. 40, 43–64 (1981).
Ebbs, S. D., Lasat, M. M., Brady, D. J., Cornish, J. & Gordon, R. Phytoextraction of cadmium and zinc from a contaminated soil. J. Environ. Quality 26, 1424–1430 (1997).
Ishikawa, S., Ae, N., Murakami, M. & Wagatsuma, T. Is Brassica juncea a suitable plant for phytoremediation of cadmium in soils with moderately low cadmium contamination? – Possibility of using other plant species for Cd-phytoextraction. Soil Sci. Plant Nutr. 52, 32–42 (2006).
Ibaraki, T., Kuroyanagi, N. & Murakami, M. Practical phytoextraction in cadmium-polluted paddy fields using a high cadmium accumulating rice plant cultured by early drainage of irrigation water. Soil Sci. Plant Nutr. 55, 421–427 (2009).
Murakami, M., Nakagawa, F., Ae, N., Ito, M. & Arao, T. Phytoextraction by rice capable of accumulating Cd at high levels: reduction of Cd content of rice grain. Environ. Sci. & Tech. 43, 5878–5883 (2009).
Uraguchi, S. et al. Root-to-shoot Cd translocation via the xylem is the major process determining shoot and grain cadmium accumulation in rice. J. Exp. Bot. 60, 2677–2688 (2009).
Ishimaru, Y., Bashir, K. & Nishizawa, N. Zn uptake and translocation in rice plants. Rice 4, 21–27 (2011).
Takahashi, R. et al. The OsNRAMP1 iron transporter is involved in Cd accumulation in rice. J. Exp. Bot. 62, 4843–4850 (2011).
Cohen, A., Nelson, H. & Nelson, N. The Family of SMF Metal Ion Transporters in Yeast Cells. J. Biol. Chem. 275, 33388–33394 (2000).
Van Ho, A., Ward, D. M. & Kaplan, J. Transition metal transport in yeast. Annual Review Microb. 56, 237–261 (2002).
Li, Z.-S. et al. A new pathway for vacuolar cadmium sequestration in Saccharomyces cerevisiae: YCF1-catalyzed transport of bis(glutathionato)cadmium. Proc. Natl. Acad. Sci. USA 94, 42–47 (1997).
Thomine, S., Wang, R., Ward, J. M., Crawford, N. M. & Schroeder, J. I. Cadmium and iron transport by members of a plant metal transporter family in Arabidopsis with homology to Nramp genes. Proc. Natl. Acad. Sci. USA 97, 4991–4996 (2000).
Ogawa, I., Nakanishi, H., Mori, S. & Nishizawa, N. Time course analysis of gene regulation under cadmium stress in rice. Plant & Soil 325, 97–108 (2009).
Supek, F., Supekova, L., Nelson, H. & Nelson, N. A yeast manganese transporter related to the macrophage protein involved in conferring resistance to mycobacteria. Proc. Natl. Acad. Sci. USA 93, 5105–5110 (1996).
Lanquar, V. et al. Mobilization of vacuolar iron by AtNRAMP3 and AtNRAMP4 is essential for seed germination on low iron. EMBO J. 24, 4041–4051 (2005).
Fox, T. C. & Guerinot, M. L. Molecular biology of cation transport in plants. Annual Review Plant Physiol. & Plant Mol. Biol. 49, 669–696 (1998).
Ishimaru, Y. et al. Rice-specific mitochondrial iron-regulated gene (MIR) plays an important role in iron homeostasis. Mol. Plant 2, 1059–1066 (2009).
Ueno, D. et al. Gene limiting cadmium accumulation in rice. Proc. Natl. Acad. Sci. USA 107, 16500–16505 (2010).
Nakanishi, H., Ogawa, I., Ishimaru, Y., Mori, S. & Nishizawa, N. K. Iron deficiency enhances cadmium uptake and translocation mediated by the Fe2+ transporters OsIRT1 and OsIRT2 in rice. Soil Sci. Plant Nutr. 52, 464–469 (2006).
Ishimaru, Y. et al. Mutational reconstructed ferric chelate reductase confers enhanced tolerance in rice to iron deficiency in calcareous soil. Proc. Natl. Acad. Sci. USA 104, 7373–7378 (2007).
Ishikawa, S., Noriharu, A., Murakami, M. & Wagatsuma, T. Is Brassica juncea a suitable plant for phytoremediation of cadmium in soils with moderately low cadmium contamination? – Possibility of using other plant species for Cd-hytoextraction. Soil Sci. Plant Nutr. 52, 32–42 (2006).
Lidon, F. C., Barreiro, M. G. & Ramalho, J. C. Manganese accumulation in rice: implications for photosynthetic functioning. J. Plant Physiol. 161, 1235–1244 (2004).
Bashir, K. et al. The rice mitochondrial iron transporter is essential for plant growth. Nat Commun 2, 322, 10.1038/ncomms1326 (2011).
Shimo, H. et al. Low cadmium (LCD), a novel gene related to cadmium tolerance and accumulation in rice. J. Exp. Bot. 62, 5727–5734 (2011).
Karimi, M., Inzé, D. & Depicker, A. GATEWAY(TM) vectors for Agrobacterium-mediated plant transformation. Trends in plant Sci. 7, 193–195 (2002).
Rentsch, D. et al. NTR1 encodes a high affinity oligopeptide transporter in Arabidopsis. FEBS Letters 370, 264–268 (1995).
Acknowledgements
We thank Dr. Yoshiaki Nagamura of the Rice Genome Project and the NIAS DNA Bank (Tsukuba, Japan) for support in the microarray analysis; Dr. Takafumi Mizuno of Mie University (Mie, Japan) for providing the yeast mutant; and Dr. Daisei Ueno of Kochi University (Kochi, Japan) and Dr. Satoshi Mori from The University of Tokyo (Tokyo, Japan) for helpful discussions. We are obliged to Dr. Pax Blamey from The University of Queensland (Queensland, Australia) for carefully reading the manuscript. This work was supported by the Program for the Promotion of Basic Research Activities for Innovative Biosciences (PROBRAIN) and a grant from the Ministry of Agriculture, Forestry and Fisheries of Japan (Genomics for Agricultural Innovation, GMB0001).
Author information
Authors and Affiliations
Contributions
Y.I., H.N. and N.N. designed the research, Y.I., K.B., R.T., S.I., T.A., H.N., N.N discussed the data and Y.I., K.B., R.T., H.N., N.N wrote the paper. Yeast complementation, GFP and GUS analysis was performed by Y.I. Rice transformations (Tsukinohikari) were performed by H.S. Rice transformations (Anjana Dhan) were performed by K.S., K.O. and M.Y. Metal concentration analysis was performed by Y.I., H.S., R.T. and T.S. Microarray and expression analyses were performed by Y.I. Statistical analyses were performed by K.B.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Additional information
Accession codes: The microarray data have been deposited in Gene Expression Omnibus database under the accession codes GSE35502.
Electronic supplementary material
Supplementary Information
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareALike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article
Cite this article
Ishimaru, Y., Takahashi, R., Bashir, K. et al. Characterizing the role of rice NRAMP5 in Manganese, Iron and Cadmium Transport. Sci Rep 2, 286 (2012). https://doi.org/10.1038/srep00286
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep00286
This article is cited by
-
Nano-enabled agrochemicals: mitigating heavy metal toxicity and enhancing crop adaptability for sustainable crop production
Journal of Nanobiotechnology (2024)
-
Antibacterial Activity, Phytochemical Analysis of C. coggygria Leaf Extracts Against S. typhi, and Computational Studies to Screen Potential Drug Candidates
Indian Journal of Microbiology (2024)
-
Cadmium toxicity: its’ uptake and retaliation by plant defence system and ja signaling
BioMetals (2024)
-
Process Optimization of Antimicrobial Activity of Adhatoda vasica Against MDR Pathogens Using Response Surface Methodology
Proceedings of the National Academy of Sciences, India Section B: Biological Sciences (2024)
-
Potassium transporter OsHAK17 may contribute to saline-alkaline tolerant mechanisms in rice (Oryza sativa)
Journal of Plant Research (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
