
Abstract
Freshwater wetlands have experienced disproportionate biodiversity loss due to environmental change. The lack of long-term biological data limits our understanding of wetland biodiversity dynamics and principal drivers. Here, we combine aquatic biomonitoring with satellite-derived imagery and spatial-explicit analysis to assess the changes in macroinvertebrate diversity and driving factors from 1988 to 2017 in Dongting Lake, China. The results revealed declining macroinvertebrate alpha diversity (species richness and functional redundancy) but increased beta diversity (taxonomic and functional dissimilarity). An increase in nutrients due to changes in land-use intensity in the surrounding terrestrial ecosystem was the primary mechanism for the losses of species and functional groups. Alpha diversity changes were most influenced by land-use changes in adjacent land areas 1.5 kilometers from the sampling sites, while beta diversity was affected within 50 meters. Our results highlight a need for attenuating land-use effects by establishing protected buffer areas to protect biodiversity in the future.
Similar content being viewed by others


Introduction
Wetlands provide essential ecological functions and services for human, animal, and plant life, including carbon storage, water supplies, fisheries, biodiversity, and recreation, but are under increased threat from human activities and climate change1. Since 1990, at least 64–71% of wetlands worldwide have suffered degradation, which includes changes to their physical states2, and these changes may result in losses of biodiversity and jeopardize the functions and stability of their ecosystems3. In fact, the past 30 years represent a critical time period of global change, especially in developing regions where wetland ecosystems have undergone drastic changes4,5. Numerous studies have verified the species loss of terrestrial and aquatic vertebrates in the Anthropocene6,7,8 (but see Su et al.9). However, there is still a relatively limited understanding of the multidecadal patterns of changes in freshwater macroinvertebrate communities. In response to those substantial changes, the Ramsar Convention on wetlands, an intergovernmental treaty with 172 contracting parties, encourages ongoing ecological monitoring and the formulation of wetland management plans to promote their sustainable use and halt future biodiversity decline10. Therefore, in order to develop effective management schemes consistent with these goals, we must understand the drivers of wetland ecosystem deteriorations.
Climate change, land-use changes, and hydrological alteration, as well as pollution, eutrophication, increasing water temperature, and decreasing lake area, are known to threaten the health and sustainability of wetland ecosystems4,11. These environmental changes often occur concurrently over a long timescale and result in habitat degradation and biodiversity losses12. Evidence has shown that warmer climates can alter the physical state of lakes, such as lake surface temperature, water levels, mixing patterns, etc., thus increasing nutrient cycling and enhancing production primary productivity13,14. Furthermore, intensive agriculture has fundamentally altered the hydrologic cycle at the Earth’s surface by changing the availability of active nitrogen at large scales15. Aquatic and terrestrial ecosystems have frequent nutrient exchanges16, where land-use changes, especially the increase in agricultural land, exacerbate the eutrophication of wetland ecosystems17. The seepage of nitrogen from agricultural land has been proven to reduce water quality at local, regional, and continental scales18, and these changes can modify freshwater community composition, species diversity, and biological integrity at multiple trophic levels (diatoms, invertebrates, and fish)12. Moreover, artificial modifications of the river-lake connections may provoke habitat degradation of lakes. For instance, it is mainly believed that the Three Gorges Dam has caused the loss of lake wetland area in the middle and lower reaches of the Yangtze River19, leading to accelerated vegetation area expansion and the response of algal blooms to hydrology20,21.
The lack of long-term biological indicators (e.g., macroinvertebrates) in China limits the understanding of historical changes in biodiversity in response to environmental factors, complicating the identification of major drivers of wetland declines22. In wetland ecosystems, macroinvertebrates play essential roles in regulating primary productivity, decomposition, and nutrient cycling, and macroinvertebrate communities are undergoing substantial changes due to multiple environmental stressors; thus, these organisms represent critical indicators of the state of the entire ecosystem23. A long-term record of macroinvertebrate diversity is essential to explore the key drivers affecting ecosystem health. The lack of established freshwater biomonitoring systems in China prior to 2010 has made it challenging to capture longer-term freshwater community dynamics24.
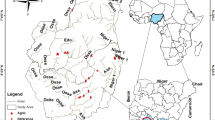
However, among the lakes in China, Dongting Lake, which is the second largest lake (lake area: 2820 km2; max depth: 31 m) connected to the Yangtze River (Supplementary Fig. 1), and is listed as a typically altered wetland in the Ramsar Convention on Wetlands25,26,27, has the longest and most complete continuous biological monitoring data in the country, with a record spanning 30 years (since 1988), since it was the first freshwater ecosystem to be the subject of routine biomonitoring. The region around Dongting Lake is one of the most important agricultural production areas in China and has a history of reclaiming farmland from wetlands associated with the lake28. The declines in biodiversity and aquatic community integrity over the past three decades in Dongting Lake have been accompanied by changes in land-use, climate change, and local hydrology19,29. Nevertheless, the relative importance of these drivers has not yet been quantified.
Here, we combined a continuous 30-year-long biomonitoring dataset from Dongting Lake with satellite imagery to investigate the drivers of local ecosystem change (Supplementary Fig. 1 and Supplementary Table 1). We hypothesize that human-induced changes, including changes in land-use, climate, and hydrology, led to a decline in the integrity of the aquatic community in Dongting Lake (Fig. 1a). We further hypothesize that these changes occurred along three paths:
-
1.
The “land-water connection” path: Increases in the land-use intensity of the terrestrial ecosystem around the lake by human populations lead to an increase in dissolved nutrients in the water, which subsequently alters macroinvertebrate communities.
-
2.
The “climate change” path: Global warming leads to an increase in lake water temperature30, which directly affects the macroinvertebrate community or indirectly leads to the decline of the community by promoting nutrient increases and algal blooms.
-
3.
The “river-lake connection” path: The weakening of the connectivity with the Yangtze River leads to the shrinkage of Dongting Lake, which directly leads to a reduction in macroinvertebrate habitat.

a Three possible pathways. Solid and dashed lines represent direct and indirect effects, respectively. b Six major environmental parameters: human land-use intensity, mean annual temperature, input charge from the Yangtze River, total nitrogen, total phosphorus, and algal bloom intensity. The arrows to the left of the panel represent the important events that might have influenced the ecological status of Dongting Lake. c Annual variation in the species richness of macroinvertebrates. d Annual variation in the functional redundancy. e Annual variation in the taxonomic dissimilarity. f Annual variation in the functional dissimilarity. The dashed lines in (c) and (d) show the gamma diversity for each year. Taxonomic and functional dissimilarity was represented by Bray‒Curtis distances between different sampling sites. The colored envelope surrounding each line represents 95% of generalized additive models.
To explore these paths, we aimed to (A) analyze the temporal trends of land-use, climate change, river-lake connectivity, and the taxonomic and functional diversity of macroinvertebrates at the scale of Dongting Lake; (B) test and identify how the major mechanisms associated with these three human-related factors impact the diversity of macroinvertebrates; and (C) further evaluate the spatial extent of the impact that land-use changes have on biodiversity shifts, that is, identify the distance at which land-use change has a significant impact on lake biodiversity. By analyzing the longest aquatic biomonitoring data series in China, we revealed the 30-year dynamics of the ecological status of Dongting Lake and identified the main drivers impacting that change from among multiple stressors.
Results
Decadal fluctuation in the ecological status of Dongting Lake
To determine changes in the ecological status of Dongting Lake over the 30-year period relative to anthropogenically driven factors, we integrated continuous global satellite imaging and biomonitoring over the past 30 years.
Environmental parameters
From 1988 to 2017, the region around the Dongting Lake wetland experienced comprehensive environmental changes, such as an increase in land-use intensity, warming of the climate, fluctuations in its connectivity with the Yangtze River, and increasing frequency of algal blooms (Fig. 1b–e). Significant tipping points of ecological change were identified in approximately 2000 and 2010 (Supplementary Fig. 2 and Supplementary Table 2). The land-use intensity did not change significantly before 1996 or after 2009 but increased significantly from 1996 to 2009 (p < 0.001). The overall annual average temperature increased by approximately 0.91 °C over the 30 years studied (p = 0.021), but no significant changes were identified at different time segments (p > 0.05). Input charge from the Yangtze River was chosen as the proxy of the connectivity with the Yangtze River, and this parameter was found to be significantly correlated with the other three hydrological parameters (TISD, FTSD, and NSD, Supplementary Fig. 3). Input charge from the Yangtze River significantly decreased 267.68 × 108 m3 from 1997 to 2009 (p = 0.021) and was jointly affected by the Three Gorges Dam and precipitation conditions (Supplementary Fig. 4). Total nitrogen and total phosphorus increased by 0.87 mg L−1 and 0.05 mg L−1 from 1988 to 2017, respectively (TN p < 0.001, TP p = 0.031), but their temporal dynamics were different (Fig. 1b): total nitrogen increased significantly from 2006 to 2010 (p = 0.042), while total phosphorus decreased substantially from 2001 to 2017 (p = 0.003). The density of planktonic algae increased by 10,000 to 40,000 cells L−1 per year (Supplementary Fig. 5). Overall, during the study period, the algal density increased before 2013, and the algal bloom frequency remained stable after 2013 (Fig. 1b and Supplementary Table 2).
Alpha diversity
The three-decade time series analysis showed how the taxonomic (species richness) and functional (functional redundancy) alpha diversity of macroinvertebrates decreased considerably between 1988 and 2017 (Fig. 1c, d). The species richness from 8 sites within Dongting Lake exhibited significant tipping points in 2001 and 2012 (Supplementary Fig. 2), between which the trend of the species richness decline was more pronounced (Supplementary Table 2). The functional redundancy of macroinvertebrate communities, described by multiple traits related to body shape, feeding type, life cycle, and other life history traits (Supplementary Table 3), changed significantly in 1994 and 2001 (Supplementary Table 2). The species richness and functional redundancy of macroinvertebrates decreased significantly at six and three sites, respectively (Supplementary Fig. 5).
Beta diversity
The taxonomic and functional composition of macroinvertebrate communities were altered in approximately 2000 and 2010, with the spatial beta diversity significantly increasing in the last 30 years (Fig. 1e, f).
The taxonomic composition of macroinvertebrate communities changed significantly in 2000, 2009, and 2013 (Supplementary Fig. 6). Initially, the composition was characterized by a wide diversity of taxonomic groups, including Insecta and Bivalvia, which significantly shifted toward Oligochaeta over the last three decades (Supplementary Fig. 7 and Supplementary Table 4). The taxonomic spatial dissimilarity (hereafter, taxonomic dissimilarity) exhibited significant tipping points in 2001 and 2012, increasing significantly from 1988 to 2001 and decreasing significantly from 2001 to 2012, with a more pronounced decline after 2012 (Supplementary Table 2).
The functional composition was altered considerably in approximately 2010 (Supplementary Fig. 6). More specifically, the behavior of organisms strongly corresponded to the annual alteration of the functional composition of macroinvertebrates (LME p = 0.031), with the abundance and species richness of Burrowers declining annually (Supplementary Fig. 7 and Supplementary Table 4). Additionally, the functional spatial dissimilarity (hereafter, the functional dissimilarity) increased significantly over the study period, with tipping points in 1999 and 2009. The annual taxonomic and functional dissimilarity patterns mainly reflected species turnover rather than the nesting of subsets of species (Supplementary Fig. 8).
Identification of major drivers and paths
We applied path analysis based on piecewise fitting of component hierarchical linear-mixed-effects models to test the three hypotheses in Fig. 1 for the dynamic changes of the macroinvertebrate communities in Dongting Lake.
Alpha diversity
In the investigation of alpha diversity, the nonsignificant p value (p = 0.188) obtained for the confirmatory path analysis indicated that the model fully supported the hypothetical causal network (Fig. 2a). The effect of increased nutrients in the lake caused by increasing human land-use intensity on the shifts in macroinvertebrate communities exceeded those of other mechanisms. The GLS regression results showed that total nitrogen was the factor that most strongly resulted in the reductions in species richness and functional redundancy (Supplementary Fig. 9). Therefore, among the three hypotheses tested, the land–water connection was identified as the dominant path by which the invertebrate community changed. In contrast, the impacts of climate change and the river–lake connection were relatively weak (Fig. 2a). The increased intensity of human land-use in the surrounding terrestrial ecosystem significantly increased the total nitrogen concentration in the water body (Fig. 2b, LME p = 0.007). The reduction in species richness and functional redundancy of macroinvertebrates was directly related to total nitrogen concentrations (Fig. 2c, LME p < 0.001) and was indirectly related to exacerbated algal bloom occurrence (Fig. 2d, e, LME p < 0.001).

a Hypotheses about the influences of various factors were checked by a structural equation model. Positive and negative paths are shown as blue and red lines, respectively, and nonsignificant paths are shown in gray. b–e Mixed linear models were fitted for vital paths. The site was used as the random factor to address nonindependence between samples. The colored envelope surrounding each line represents 95% of linear mixed models.
Beta diversity
The beta diversity path analysis further verified the important role of human land-use in the temporal alteration of macroinvertebrates (Fig. 3). The results of the GLS model revealed that taxonomic and functional dissimilarities were jointly controlled by human land-use intensity, total phosphorous, and algal bloom intensity (Supplementary Fig. 9). The direct impact of human land-use differences on taxonomic dissimilarity was the main path that led to the increase in macroinvertebrate beta diversity (Fig. 3, p < 0.001). Meanwhile, the total phosphorus differences between sites, accelerated by land-use alteration, significantly increased community dissimilarity (Fig. 3, p < 0.001).

a Hypotheses about the influences of various factors were checked by a structural equation model. Positive and negative paths are shown as blue and red lines, respectively, and nonsignificant paths are shown in gray. b–e Mixed linear models were fitted for vital paths. Taxonomic and functional dissimilarity were standardized by geographical distance to minimize the effect of distance between sites. The colored envelope surrounding each line represents 95% of linear models.
The spatial extent of the effects of land-use on macroinvertebrate diversity
To explore the best spatial extent of the associations between human land-use intensity at the surrounding terrestrial ecosystem and parameters in the lake, we tested the associations with increasing terrestrial extent from the sample locations.
Human land-use intensity across multiple spatial extents
We extracted human land-use intensity (the total proportion of cropland and urban area) within buffer zones ranging from 50 m to 20 km around each site, over a time span ranging from 1988 to 2017. Linear fittings between human land-use intensity and the lake trophic status, and the macroinvertebrate diversity for each site and spatial extent were performed. The human land-use intensity significantly decreased (p < 0.001) between buffer distances of 50 m to 1 km but increased from 1 km to 20 km (Supplementary Fig. 10, p < 0.001). Differences in human land-use intensity between sites decreased remarkably with increasing spatial extent (Supplementary Fig. 10, p < 0.001).
Alpha diversity
The acceleration of human land-use intensity in the terrestrial ecosystems surrounding Dongting Lake over time significantly affected macroinvertebrate alpha diversity within 20 km of each sampling site, with the largest effect occurring at distances of approximately 1.5 km (Fig. 4b–e). The temporal changes in human land-use intensity were correlated with elevated total nitrogen in water and larger algal blooms within a spatial range of 7 km and 3 km, respectively (Fig. 4b, c). The intensity of the impact on macroinvertebrate alpha diversity had a unimodal relationship with the increase in buffer radius. The response of species diversity to human land-use intensity peaked at 0.9 km, while that of functional redundancy peaked at 1.5 km (Fig. 4d, e).

a The spatial distribution map of human land-use, taking buffers of multiple sizes at site “ED” as an example. b–e The explained variances in human land-use intensity across distance to trophic parameters in the water and alpha diversity of macroinvertebrates. The R2 of the null model is shown as a gray dashed line. f–i The explained variances in Δ human land-use intensity across distance to differences in trophic parameters in water and beta diversity of macroinvertebrates. The colored envelope surrounding each line represents 95% of the models. The dotted vertical lines are imported points where the fitted line reaches a plateau or hill or intersects with the null model.
Beta diversity
The human land-use intensity differences (ΔHLU) increased taxonomic and functional dissimilarities between sites, and the effects peaked within 50 m of the sites (Fig. 4f–i). Differences in lake water total phosphorus were significantly associated with human land-use alteration only in the 0.5 km to 0.8 km buffer zones (Fig. 4f). Human land-use in all the buffer zones significantly accelerated the spatial differences in algal blooms, with a maximum difference occurring at 15 km (Fig. 4g). The explained variances between human land-use and the taxonomic and functional dissimilarities were highest in the 50 m buffer zone (Fig. 4h, i).
Discussion
Understanding the drivers of wetland ecosystem decline is essential for effective management, given the numerous factors jeopardizing their health and sustainability23,31. Our results suggest that the impact of land-use changes has exceeded that of climate change and hydrological alteration over the past 30 years, leading to the loss of macroinvertebrate diversity in Dongting Lake. We found that the acceleration of land-use intensity in the terrestrial ecosystem surrounding Dongting Lake, which caused an increase in nutrient loading, was the main mechanism driving the continuous changes in the macroinvertebrate communities. Therefore, managing the interface between wetlands and adjacent terrestrial ecosystems will be essential to preserving the biodiversity of these ecosystems in the future.
Biodiversity losses of macroinvertebrates
The alpha diversity of macroinvertebrates in Dongting Lake, although fluctuating, declined over the study period, and the spatial beta diversity increased, indicating a dramatic loss and shift of taxa and functional groups in macroinvertebrates. These results were in line with shifts observed in freshwater macroinvertebrate diversity over time in other countries. A study conducted in diverse rivers in England detected increases in the abundance of carnivores and herbivores, while decomposers declined from 2002 to 201932. Between 1994 and 2007, a significant decline was observed in many macroinvertebrate families that favor colder waters and faster-flowing habitats in Australia33. Meanwhile, the diversities of freshwater macroinvertebrates in the US and Europe have been gradually recovering34,35. Our results suggest that macroinvertebrate communities in Dongting Lake deteriorated at both the taxonomic and functional levels over the study period. The species richness of four macroinvertebrate classes (the other two showed no significant trend) and all functional groups decreased significantly (Supplementary Table 4). The increased beta diversity was attributed to the loss of local species, which resulted from the overall decrease in gamma diversity (Supplementary Fig. 7). In the initial stage of environmental change, the loss of local species can buffer gamma diversity and lead to unchanged or increased beta diversity36. Moreover, changes in spatial beta diversity may also depend on the relative trends of generalist and specialist species37. In summary, over the past 30 years, the macroinvertebrate community of Dongting Lake has undergone both biodiversity loss and community shift, and this declining trend has yet to be halted.
Dominate role of human land-use
We found that the increased human land-use intensity, including expanding agricultural and urban areas, was the dominant factor affecting the integrity of Dongting Lake. Multiple expectations of possible mechanisms, which could be categorized as direct effects, such as the toxicity of excessive nutrients, and indirect effects, such as anoxia caused by cyanobacteria blooms, have been proposed and thoroughly discussed in the field and mesocosm studies. Previous studies showed that excess nutrients lead to changes in the element stoichiometry of macroinvertebrates38, even causing the death of some species39 or indirectly reducing the diversity of macroinvertebrates through anoxia caused by cyanobacteria blooms40. As the global population continues to expand, the expansion and intensification of agriculture will further exacerbate the ecological degradation of wetland habitats41. In wetland research, most alleviation and restoration measures focus on the water, such as removing nutrients42,43, while land-based restoration has been studied only sporadically44. Our analyses indicate that land-use change has caused substantial degradation of a Ramsar-protected wetland ecosystem. Our analyses indicate that land-use change has caused the substantial biodiversity loss of macroinvertebrates, highlighting the importance of considering land-water relationships in wetland conservation and restoration.
Impacts of climate change
The impact of climate change on macroinvertebrates was not the dominant path in Dongting Lake, although climate change is considered one of the most serious threats to lake ecosystems globally3. Nevertheless, the effects of climate change may not be as pronounced as the effects of other concurrent environmental changes, such as those from shifting land-use practices12. For example, in eastern China, the lake area is less affected by climate than by human activities4. To date, anthropogenic pressures via land-use change have surpassed the influence that climatic factors have on the temporal variability in algal blooms45 and on biodiversity in lakes and reservoirs in China. In this study, we found that climate warming was correlated with increased nitrogen but increased phosphorus loading in Dongting Lake, although these effects were weaker than the effects induced by human land-use changes (Fig. 2). However, some studies have shown that lake warming intensifies the seasonal pattern of internal nutrient cycling and creates conditions beneficial for longer cyanobacteria blooms46. This study did not focus on this issue due to the lack of seasonal data. Further research should focus on finer time scales (e.g., quarterly, monthly) to better understand the effects of climate change from a multi-stressor perspective.
Impacts of river–lake connection
We found that the weakening of the river-lake connections had a limited impact compared to human land-use on the macroinvertebrate diversity of Dongting Lake. As the first large lake downstream of the Three Gorges Dam, Dongting Lake experienced serious changes in hydrology and sediment conditions after the dam was built, and the inundation time of Dongting Lake decreased by 3.6% from 2000 to 201019. The relative humidity and surface runoff have declined sharply since the Three Gorges impoundment in 200319 (but see Huang et al.47). However, human activities such as water intake and land reclamation around the lake also intensified dramatically during this period29, and these factors may be additional drivers causing the shrinking lake area and changing hydrological conditions. A recent study on the dynamics of algal blooms in Dongting Lake found that climatic-related parameters changed significantly before and after the completion of the Three Gorges Dam21. Here, with a random forest model, we found that the connectivity between the Yangtze River and Dongting Lake (represented as the YRQ) was jointly influenced by the Three Gorges Dam and changes in precipitation (Supplementary Fig. 4). Therefore, the co-analysis of multiple stressors helped us better understand the role of the Three Gorges Dam in the ecological fluctuation of Dongting Lake. In this study, both the alpha and beta diversities of macroinvertebrates were much less affected by the establishment of the Three Gorges Dam than by land-use change (Figs. 2 and 3). Our results indicate that under the multi-stressor scenario, the impact of dam construction on the lake ecosystem should be analyzed together with other possible factors to obtain more objective results.
The spatial extent of land-use effects
While land-use change has been the main stressor affecting Dongting Lake in the last 30 years, its effect was geographically localized, which suggests that the establishment of land buffer areas may provide substantial benefits for preserving ecosystem integrity. Current restoration and convention efforts considering the “land–water connection” face challenges in implementation due to a lack of theoretical frameworks, particularly in addressing the spatial extent of terrestrial disturbances on aquatic biodiversity48,49. A previous study on the Thur River in Switzerland found that the association between aquatic biodiversity and plant functional diversity in terrestrial ecosystems peaked in a smaller buffer area (about 400 m)49. Yet for large rivers such as the Amazon, the diversity of fishes could be impacted by deforestation over a range of 30 km50. Here, we showed that changes in the human land-use intensity of the surrounding terrestrial ecosystems significantly impact macroinvertebrate diversity, with the greatest impact found within 1.5 km of the lake (Fig. 4). Therefore, in formulating protection and restoration measures for wetlands experiencing eutrophication, including the Dongting Lake wetland, attention should be given to the management and improvement of the surrounding terrestrial ecosystems, especially the lakeside buffer zones.
Our analysis filled a gap in our understanding of the historical dynamics of wetland ecosystems in China while also providing a valuable case study for comparisons worldwide. Furthermore, our work suggested that the impact that the construction of the Three Gorges Dam had on the biodiversity loss of lakes in the middle and lower reaches of the Yangtze River might be limited. Finally, we revealed that the increase in local human land-use intensity was the main factor altering macroinvertebrate taxonomic and functional diversity. The results suggest that the land adjacent to the lake is critical for maintaining wetlands and lake biodiversity. Consequently, establishing protected land buffer areas around these wetlands and lakes may be the most suitable strategy for wetland/lake conservation, particularly in the developing world.
Methods
Macroinvertebrate community and nutrients
Data on macroinvertebrates, nutrients, and algal blooms were collected during routine monitoring by the Dongting Lake Eco-Environment Monitoring Centre of Hunan Province from 1988 to 2017. Routine monitoring of macroinvertebrates, phytoplankton, and nutrients was carried out at eight transects in the main lake (Supplementary Fig. 1). The eight transects in this study were distributed in the key habitats of Dongting Lake (southern, western, and eastern parts of Dongting Lake). In situ monitoring was conducted each year from 1988 to 2018. However, no data were collected in 2002, 2003, 2015, and 2016 due to objective reasons such as water level changes. All transects were sampled every January, May, and September to capture variability among those seasons. Samples were collected from three locations along each transect; two samples were collected at 100 m from either bank, and two samples were collected from the middle of the transect. These samples were then mixed for subsequent chemical analysis and species identification (Supplementary Table 1). Macroinvertebrate samples were sampled with a 1/16 m2 modified Peterson dredger. The samples with animal specimens were sieved, washed through a 40-mesh copper sieve, and placed in a sealed pocket for storage. On the same day, after screening and washing, the samples were put into the white porcelain dissecting plate and sorted under cold light. Then, put the selected animal sample into a 50 mL sample bottle and fix it with a 10% formalin solution. The animals were identified and fixed in the laboratory. Specimen identification mainly referred to relevant literature and was identified at the genus level. Phytoplankton samples were collected by filtering 20 L of water through a plankton net (20 mesh). The precipitates were fixed with Lugol’s solution for 48 h and counted under the microscope. The algal bloom intensity was defined as the annual average abundance of planktonic algae. For Nutrients, the duplicated 1 L surface water samples were used to measure nutrient variables, including total phosphorus (TP) and total nitrogen (TN), following the Chinese national standard “Environmental quality standards for surface water (GB 3838-2002).” See Supplementary Note 1 for details.
Land-use, climate, and hydrological data
Geospatial information for the Lake Dongting basin was obtained from the Resource and Environment Science and Data Center of the Chinese Academy of Sciences (https://www.resdc.cn/). One-kilometer-wide buffer zones encompassing the sampling sites were generated with ArcGIS 10.7. Land-use data from 1988 to 2017 were extracted from the first Landsat-derived annual China land cover dataset (CLCD)51, which classified land cover into seven types (cropland, forest, grass, pasture, urban, water, and unused). We defined human land-use intensity (HLU) as the total proportion of cropland and urban areas. Mean annual air temperature (MAT, °C) and mean monthly precipitation (MAP, mm month−1) data for the study area were obtained from the CRU TS monthly high-resolution gridded multivariate climate dataset (version 4)52.
The mainstream of the Yangtze River inputs water and sediment into Dongting Lake through three inlets (Supplementary Fig. 1 and Supplementary Table 5). Total annual water discharge data from five gauging sites, provided by the Dongting Lake Eco-Environment Monitoring Centre of Hunan Province, were used to represent the input from the Yangtze River (YRQ, 108 m3) into the lake. Moreover, to validate the ability of these data to represent important hydrological changes, sediment-related hydrological variables (i.e., sediment discharge at three inlets (TISD) and from four tributaries (FTSD) and the net sediment deposition, NSD) were collected from published literature21, and Spearman correlation analysis showed that the YRQ was significantly correlated with other hydrological parameters (Supplementary Fig. 3). To study the climate-derived and dam-derived effects on the YRQ, we extracted the number of rainstorm days (R50mm), simple precipitation intensity index (SDII), number of summer days (SU), warm spell duration index (WSDI) and yearly anomalies of ENSO events (ENSOi) from published literature21 via GetData Graph Digitizer. The Three Gorges Dam (TGD) has been shown to weaken the connectivity between the Yangtze River and Dongting Lake19. We used a categorical variable, with 0 (before 2003) and 1 (after 2003) indicating the operation status of the TGD. A random forest model with 1000 permutations and 500 trees for each iteration was fitted to identify the main drivers of YRQ dynamics using the “rfPermute” R package in R4.0.5.
Biodiversity indices
Taxonomic alpha diversity was calculated as the annual average number of macroinvertebrate species. The functional redundancy analysis process followed Toussaint et al.6. We identified 29 categories belonging to six traits (Supplementary Table 3 and Supplementary Note 2). These traits are closely related to the resilience and resistance of benthic communities, represent the biological and ecological characteristics of macroinvertebrates, and are widely used in research on the functional diversity of inland water bodies53. Principal component analysis (PCA) was used to determine the two main axes of functional trait variation. The “TPD” R software package was used to estimate the multivariate kernel density, and the probability distribution of each species in the functional space was estimated by using all species with functional information54. Finally, the redundancy () function in the “TPD” R packages was used to calculate each sample’s macroinvertebrate functional redundancy. Spatial dissimilarity (beta diversity) was defined as the distance of community composition between sites in the same year. The total spatial dissimilarity was partitioned into turnover and nestedness components with beta.pair.abund() in the “betapart” package. Taxonomic dissimilarities were based on Bray‒Curtis matrices. Based on the Soerensen matrix, functional dissimilarities were calculated and partitioned using the hypervolume algorithm55.
Temporal trend analysis
Decadal trends of major environmental parameters, namely bloom intensity, species richness, and functional redundancy, were first analyzed using generalized additive models (gam() function in the “mgcv” package) to capture their overall temporal trends. To reveal significant changes in the trends of these ecological parameters over time, we used Bayesian change point (bcp) analysis to estimate the posterior probabilities for the presence of tipping points at which the trends change using the “bcp” package in R56. To study the temporal trends before and after the identified tipping points, we defined three temporal segments for each parameter divided by years with the two most probable tipping points. Each time segment had to include at least four years of data. Temporal trend analysis was performed for the different time segments based on tipping points in Supplementary Fig. 2 using linear model fitting. To identify the significant tipping points in the taxonomic and functional community structure, we performed a multivariate regression tree (MRT) analysis using the “mvpart” package57.
Identification of major drivers
To identify the major drivers and mechanisms causing the decline in diversity and the compositional shifts of the macroinvertebrate community, we selected six explanatory variables, including human land-use intensity (HLU), mean annual temperature (MAT), input charge from the Yangtze River (YRQ), total nitrogen (TN), total phosphorus (TP) and algal bloom intensity (Bloom), based on the stated hypotheses. We assessed the potential multicollinearity among the variables by conducting pairwise Pearson correlation and variance inflation factor (VIF) analyses prior to any calculations. The results showed that all absolute correlation values (|r|) < 0.70, and VIF < 10, indicating no multicollinearity among the variables.
For alpha diversity, all explanatory and response parameters were z-score transformed prior to analysis to standardize the data. First, we applied the generalized least squares (GLS) model to investigate the relationships between biodiversity parameters and explanatory variables. The adjusted R2 of the model was computed with the function r2() from the package “performance.” The standardized effect sizes of each variable on species richness and functional redundancy were extracted with the function effectsize() from the “effectsize” package. Second, we conducted confirmatory path analysis based on piecewise fitting of component hierarchical linear mixed-effects models to explore the major mechanisms affecting biodiversity58. Path analysis is a widely accepted method for exploring complex causal relationships and assessing direct and indirect effects in a structured and systematic way59. The “NLME” package was used to fit the observed variables into the general linear mixed effect model, with sites as the random factor, to address the issue of nonindependence between samples60. The overall path model was evaluated using Shipley’s directional separation test61, which concluded that Fisher’s C could be compared with χ2 statistics for distribution comparison. If the obtained p value is >0.05, we assumed that the model fully reproduced the hypothetical causal network. Third, major paths were visualized by general linear mixed-effect models with the site as the random factor.
For beta diversity, the spatial differences in human land-use (ΔHLU), total nitrogen (ΔTN), total phosphorus (ΔTP), and algal bloom (ΔBloom) were characterized as the absolute value of the difference between sites. To eliminate differences in beta diversity between sites due to dispersal limitations62, taxonomic and functional spatial dissimilarity were standardized as the quotient of community composition distances and geographic distances. The identification of major factors affecting beta diversities followed the same method as that of alpha diversity, except fitted linear models replaced linear mixed models in path analysis.
Linear regression and null models across multiple spatial extents
Human land-use intensity was measured over multiple spatial extents. Specifically, 18 buffers around each sampling site were created at intervals of 0.05 km between 0.05 and 0.1 km, 0.1 km between 0.1 and 0.9 km, 1 km between 1 and 5 km, and 5 km between 10 and 20 km with ArcGIS 10.7. Human land-use intensities within the buffer zones were extracted, and sites with >80% water in the buffer area were removed. The absolute human land-use area depends on the surface area measured within each spatial extent, making the absolute value of human land-use dependent on the spatial extent under consideration. Similarly, within each spatial extent, a buffer from a higher spatial extent covers all points from a site from a smaller spatial extent, once again making the human land-use area dependent on the area under consideration. Therefore, the human land-use intensity was calculated as a percentage the absolute human land-use area divided by the surface area.
For each sampling site and spatial extent, linear regression models were fitted between human land-use and ecological responses (total nitrogen, bloom intensity, species richness, and functional redundancy) over time. The significance and the variance explained by each model were calculated using p values and the R2 coefficient of determination, respectively. Null models were developed to ensure that the spatial association between aquatic parameters and terrestrial land-use was not a measurement artifact. Specifically, the spatial location of pixels within the Dongting Lake basin was randomly shuffled 1000 times, and the human land-use proportion for each sampling site was extracted according to the distance buffers presented above. Then, the process applied to the “true” dataset was applied to the randomized data. To explore the spatial extent to which human land-use most affected the biodiversity of macroinvertebrates, we identified the spatial extent where R2 peaked, reached a plateau, or intersected with the null models.
This procedure was also carried out to assess beta diversity. In short, the difference in human land-use between sites (ΔHLU) was fitted to the spatial differences in total phosphorus (ΔTP), algal bloom (ΔBloom), and taxonomic and functional dissimilarities of macroinvertebrates. No null model was generated, as there was only one point for each buffer size. The best spatial extent was identified as that with the largest R2 value.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The data of China’s annual land cover product (CLDD) can be accessed at https://zenodo.org/record/5210928#.YcZ_nWBByUk. CRU TS climate data can be downloaded from the Centre for Environmental Data Analysis (CEDA: http://data.ceda.ac.uk//badc/cru/data/cru_ts/) and the CRU website: https://crudata.uea.ac.uk/cru/data/hrg/. All data that support the findings of this study are available in figshare at https://doi.org/10.6084/m9.figshare.2449969963.
Code availability
Scripts for generating the figures are available on GitHub at https://github.com/Laura61616/DongtingLake.
References
Huang, S., Zhang, K., Lin, Q., Liu, J. & Shen, J. Abrupt ecological shifts of lakes during the Anthropocene. Earth Sci. Rev. https://doi.org/10.1016/j.earscirev.2022.103981 (2022).
Davidson, N. C. How much wetland has the world lost? Long-term and recent trends in global wetland area. Mar. Freshw. Res. 65, 934–941 (2014).
Xi, Y., Peng, S., Ciais, P. & Chen, Y. Future impacts of climate change on inland Ramsar wetlands. Nat. Clim. Chang. 11, 45–51 (2020).
Tao, S. et al. Changes in China’s lakes: climate and human impacts. Natl Sci. Rev. 7, 132–140 (2020).
Wang, X. et al. Gainers and losers of surface and terrestrial water resources in China during 1989-2016. Nat. Commun. 11, 3471 (2020).
Toussaint, A. et al. Extinction of threatened vertebrates will lead to idiosyncratic changes in functional diversity across the world. Nat. Commun. 12, 5162 (2021).
Brodie, J. F., Williams, S. & Garner, B. The decline of mammal functional and evolutionary diversity worldwide. Proc. Natl Acad. Sci. USA https://doi.org/10.1073/pnas.1921849118 (2021).
Zhang, H. et al. Extinction of one of the world’s largest freshwater fishes: lessons for conserving the endangered Yangtze fauna. Sci. Total Environ. 710, 136242 (2020).
Su, G. et al. Human impacts on global freshwater fish biodiversity. Science 371, 835–838 (2021).
Long, K. E. et al. Human impacts and Anthropocene environmental change at Lake Kutubu, a Ramsar wetland in Papua New Guinea. Proc. Natl Acad. Sci. USA https://doi.org/10.1073/pnas.2022216118 (2021).
Birk, S. et al. Impacts of multiple stressors on freshwater biota across spatial scales and ecosystems. Nat. Ecol. Evol. https://doi.org/10.1038/s41559-020-1216-4 (2020).
Li, F. et al. Human activities’ fingerprint on multitrophic biodiversity and ecosystem functions across a major river catchment in China. Glob. Chang. Biol. 26, 6867–6879 (2020).
Moss, B. Cogs in the endless machine: lakes, climate change and nutrient cycles: a review. Sci Total Environ 434, 130–142 (2012).
Woolway, R. I. et al. Lake heatwaves under climate change. Nature 589, 402–407 (2021).
Peñuelas, J. et al. Human-induced nitrogen–phosphorus imbalances alter natural and managed ecosystems across the globe. Nat. Commun. 4, 2934 (2013).
Harvey, E., Gounand, I., Fronhofer, E. A. & Altermatt, F. Metaecosystem dynamics drive community composition in experimental, multi-layered spatial networks. Oikos 129, 402–412 (2020).
Michalak Anna, M. et al. Record-setting algal bloom in Lake Erie caused by agricultural and meteorological trends consistent with expected future conditions. Proc. Natl Acad. Sci. USA 110, 6448–6452 (2013).
Yu, C. et al. Managing nitrogen to restore water quality in China. Nature 567, 516–520 (2019).
Feng, L., Hu, C., Chen, X. & Zhao, X. Dramatic inundation changes of China’s two largest freshwater lakes linked to the Three Gorges Dam. Environ. Sci. Technol. 47, 9628–9634 (2013).
Xie, Y.-H., Yue, T., Xin-sheng, C., Feng, L. & Zheng-miao, D. The impact of Three Gorges Dam on the downstream eco-hydrological environment and vegetation distribution of East Dongting Lake. Ecohydrology 8, 738–746 (2015).
Fu, H. et al. Linking human activities and global climatic oscillation to phytoplankton dynamics in a subtropical lake. Water Res. 208, 117866 (2022).
Lin, J. et al. Making China’s water data accessible, usable and shareable. Nat. Water 1, 328–335 (2023).
Dalu, T. et al. Drivers of aquatic macroinvertebrate communities in a Ramsar declared wetland system. Sci. Total Environ. 818, 151683 (2022).
Feio, M. J. et al. The biological assessment and rehabilitation of the world’s rivers: an overview. Water 13, 371 (2021).
Ramsar. Dong dongting hu. https://rsis.ramsar.org/ris/551 (2013).
Ramsar. Nan Dongting wetland and waterfowl nature reserve. https://rsis.ramsar.org/ris/1151 (2007).
Ramsar. Xi Dongting Lake (Mupinghu) nature reserve. https://rsis.ramsar.org/ris/1154 (2007).
Xie, C., Huang, X., Mu, H. & Yin, W. Impacts of land-use changes on the lakes across the Yangtze Floodplain in China. Environ. Sci. Technol. 51, 3669–3677 (2017).
Yang, Z.-M. et al. Spatial and temporal changes in wetland in Dongting Lake Basin of China under long time series from 1990 to 2020. Sustainability https://doi.org/10.3390/su14063620 (2022).
Woolway, R. I. et al. Global lake responses to climate change. Nat. Rev. Earth Environ. 1, 388–403 (2020).
Fluet-Chouinard, E. et al. Extensive global wetland loss over the past three centuries. Nature 614, 281–286 (2023).
Powell, K. E. et al. Abundance trends for river macroinvertebrates vary across taxa, trophic group and river typology. Glob. Chang. Biol. https://doi.org/10.1111/gcb.16549 (2022).
Chessman, B. C. Climatic changes and 13-year trends in stream macroinvertebrate assemblages in New South Wales, Australia. Glob. Chang. Biol. 15, 2791–2802 (2009).
Haase, P. et al. The recovery of European freshwater biodiversity has come to a halt. Nature https://doi.org/10.1038/s41586-023-06400-1 (2023).
Artz, C., Pyron, M. & Bowley, L. Long-term macroinvertebrate assemblages of the West Fork White River, Indiana improve following the Clean Water Act. Am. Midland Naturalist https://doi.org/10.1674/0003-0031-184.2.233 (2020).
Socolar, J. B., Gilroy, J. J., Kunin, W. E. & Edwards, D. P. How should beta-diversity inform biodiversity conservation? Trends Ecol. Evol. 31, 67–80 (2016).
Ellingsen, K. E. et al. The rise of a marine generalist predator and the fall of beta diversity. Glob. Chang. Biol. 26, 2897–2907 (2020).
Karimi, R. & Folt, C. L. Beyond macronutrients: element variability and multielement stoichiometry in freshwater invertebrates. Ecol. Lett. 9, 1273–1283 (2006).
Struijs, J., De Zwart, D., Posthuma, L., Leuven, R. S. & Huijbregts, M. A. Field sensitivity distribution of macroinvertebrates for phosphorus in inland waters. Integr. Environ. Assess. Manag. 7, 280–286 (2011).
Huisman, J. et al. Cyanobacterial blooms. Nat. Rev. Microbiol. 16, 471–483 (2018).
Hansen, A. T., Dolph, C. L., Foufoula-Georgiou, E. & Finlay, J. C. Contribution of wetlands to nitrate removal at the watershed scale. Nat. Geosci. 11, 127–132 (2018).
Tickner, D. et al. Bending the curve of global freshwater biodiversity loss: an emergency recovery plan. Bioscience 70, 330–342 (2020).
Horppila, J. Sediment nutrients, ecological status and restoration of lakes. Water Res. 160, 206–208 (2019).
Tian, B., Wu, W., Yang, Z. & Zhou, Y. Drivers, trends, and potential impacts of long-term coastal reclamation in China from 1985 to 2010. Estuarine, Coast. Shelf Sci. 170, 83–90 (2016).
Song, K. et al. Climatic versus anthropogenic controls of decadal trends (1983-2017) in algal blooms in lakes and reservoirs across China. Environ. Sci. Technol. 55, 2929–2938 (2021).
Yindong, T. et al. Lake warming intensifies the seasonal pattern of internal nutrient cycling in the eutrophic lake and potential impacts on algal blooms. Water Res. 188, 116570 (2021).
Huang, Q. et al. Hydrological drought at Dongting Lake: its detection, characterization, and challenges associated with three gorges dam in Central Yangtze, China. Water Resourc. Manag. 28, 5377–5388 (2014).
Linke, S., Pressey, R. L., Bailey, R. C. & Norris, R. H. Management options for river conservation planning: condition and conservation re‐visited. Freshw. Biol. 52, 918–938 (2007).
Zhang, H. et al. A spatial fingerprint of land-water linkage of biodiversity uncovered by remote sensing and environmental DNA. Sci. Total Environ. https://doi.org/10.1016/j.scitotenv.2022.161365 (2023).
Cantera, I. et al. Low level of anthropization linked to harsh vertebrate biodiversity declines in Amazonia. Nat. Commun. 13, 3290 (2022).
Yang, J. & Huang, X. The 30 m annual land cover dataset and its dynamics in China from 1990 to 2019. Earth Syst. Sci. Data 13, 3907–3925 (2021).
Harris, I., Osborn, T. J., Jones, P. & Lister, D. Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Sci. Data 7, 109 (2020).
Heino, J. & Tolonen, K. T. Untangling the assembly of littoral macroinvertebrate communities through measures of functional and phylogenetic alpha diversity. Freshwater Biology 62, 1168–1179 (2017).
Mammen, E., Nielsen, J. P. & Fitzenberger, B. Generalized linear time series regression. Biometrika 98, 1007–1014 (2011).
Carvalho, J. C. & Cardoso, P. Decomposing the causes for niche differentiation between species using hypervolumes. Front. Ecol. Evol. https://doi.org/10.3389/fevo.2020.00243 (2020).
Erdman, C. & Emerson, J. W. bcp: An R package for performing a Bayesian analysis of change point problems. J. Stat. Softw. 23, 1–13 (2007).
De’ath, G. Multivariate regression trees: a new technique for modeling species–environment relationships. Ecology 83, 1105–1117 (2002).
Lefcheck, J. S. piecewiseSEM: piecewise structural equation modelling in r for ecology, evolution, and systematics. Methods Ecol. Evol. 7, 573–579 (2016).
Wootton, J. T. Predicting direct and indirect effects: an integrated approach using experiments and path analysis. Ecology 75, 151–165 (1994).
Pinheiro, J., Bates, D., DebRoy, S. S. & Sarkar, D. Nlme: linear and nonlinear mixed effects models. R package version 31-110. https://cran.r-project.org/web/packages/nlme/nlme.pdf (2013).
Shipley, B. Confirmatory path analysis in a generalized multilevel context. Ecology 90, 363–368 (2009).
Hubbell, S. The Unified Neutral Theory of Biodiversity and Biogeography (The University of Chicago Press, 2001).
Zhang, Y. et al. Data for: Long-term wetland biomonitoring highlights the different impacts of anthropogenically induced factors on regional macroinvertebrate diversity in Dongting Lake, China. figshare https://doi.org/10.6084/m9.figshare.24499699.v1 (2023).
Acknowledgements
This work was supported by the National Key Research and Development Program of China (2021YFC3201003; 2021YFC3200105; 2022YFC3202101) and the National Natural Science Foundation of China (Grant Nos. 41977206; 41977364; 42322710).
Author information
Authors and Affiliations
Contributions
Y.Z. conceived and designed the study, analyzed data, and wrote the manuscript; D.H. conceived and designed the study, performed experiments, and critically reviewed the manuscript; X.J. and X.Z. supervised the overall project, provided funding, and contributed to manuscript editing and finalization; L.L. and C.W. conducted fieldwork, collected and interpreted data, and contributed to manuscript preparation; Y.W., L.P., A.C.J., and F.W. critically revised the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Earth & Environment thanks the anonymous reviewers for their contribution to the peer review of this work. Primary handling editors: Huai Chen, Clare Davis, Martina Grecequet, Heike Langenberg. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhang, Y., Huang, D., Jin, X. et al. Long-term wetland biomonitoring highlights the differential impact of land use on macroinvertebrate diversity in Dongting Lake in China. Commun Earth Environ 5, 32 (2024). https://doi.org/10.1038/s43247-024-01203-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s43247-024-01203-2
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
