
Abstract
Climate change strongly impact vegetation phenology, with considerable potential to alter land-atmosphere carbon dioxide exchange and terrestrial carbon cycle. In contrast to well-studied spring leaf-out, the timing and magnitude of autumn senescence remains poorly understood. Here, we use monthly decreases in Normalized Difference Vegetation Index satellite retrievals and their trends to surrogate the speed of autumn senescence during 1982–2018 in the Northern Hemisphere (>30°N). We find that climate warming accelerated senescence in July, but this influence usually reversed in later summer and early autumn. Interestingly, summer greening causes canopy senescence to appear later compared to an advancing trend after eliminating the greening effect. This finding suggests that summer canopy greening may counteract the intrinsic changes in autumnal leaf senescence. Our analysis of autumn vegetation behavior provides reliable guidance for developing and parameterizing land surface models that contain an interactive dynamic vegetation module for placement in coupled Earth System Models.
Similar content being viewed by others


Introduction
Autumnal senescence is a critical component of the seasonal evolution of terrestrial ecosystems, which regulates the length of the growing season and terrestrial carbon uptake1,2. Yet, such event and its response to changing climate is not well represented in most Earth System Models (ESMs)3. Impact of climate change on the trends of autumn senescence as well as its role in terrestrial carbon, water, nutrient cycling, fitness, and distribution of tree species have been reported over the past decades1,2,4,5,6,7,8,9. Many of them focused on the timing of specific phenophase, e.g., leaf coloring and abscission1. However, the process of autumn senescence consists of a series of complex metabolic events such as macromolecule degradation, decreased photosynthesis, and most importantly, nutrient re-absorption10,11,12. These events are co-dominated by plant internal conditions (e.g., metabolic adjustments, genetic expressions), near-surface meteorological factors (e.g., temperature, solar radiation and wind), water availability and their interactions12,13,14. Despite the complex climatic responses of autumn phenology, it remains unclear, however, which stage of the senescence process contributes more to changes in the timing of end of the growing season (EOS). Moreover, previous studies pay too much attention to individual phenophases (e.g., EOS), which may conceal other components of the senescence process, such as the variations in the environmental regulations during the multiple stages of the autumn senescence process15. These gaps in knowledge would leave the reliable modelling of autumn senescence in doubt16, and finally hamper the ability of ESMs to simulate vegetation dynamics under future climatic scenarios17,18.
Here, we provide a timely assessment of autumn senescence speed through the monthly decreases in NDVI (ΔNDVI, see schematic representation in Fig. 1), which is also critical for the extraction of EOS. We first investigate the regulations of climatic factors (temperature, soil moisture, insolation i.e., downward shortwave radiation, and windspeed) on the speed of autumn senescence (i.e., the interannual trends in ΔNDVI). Given that such behavior may be confounded by rising CO2-induced summer canopy greening (i.e., uptrends in annual maximum NDVI, NDVImax), we additionally calculated the relative decrease of NDVI (δNDVI = ΔNDVI/NDVImax) as a surrogate for ΔNDVI (see definitions in Supplementary Table 1). Then, we quantified the effects of increasing NDVImax (i.e., canopy greening) on ΔNDVI, and consequently, the extracted EOS. Together with this analysis, we discussed the implications of our study on benchmarking phenological modules for future generations of Earth System Models (ESMs).

In this study, autumn senescence starts at Peak Of Season (POS) when NDVI reaches its annual maximum (NDVImax), and ends at the End Of Season (EOS). The monthly decrease in NDVI (ΔNDVI) is estimated as the NDVI difference between the current and the following months (in positive values), and the remaining NDVI is the NDVI left in the EOS month (in positive values). The sum of monthly ΔNDVI and the remaining NDVI is therefore equal to NDVImax. Detailed definitions of these items can be found in Supplementary Table 1.
Results and Discussion
Changes in the speed of autumn senescence
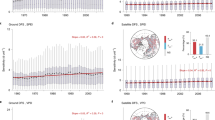
Across the Northern Hemisphere (>30°N, NH, thereafter), divergent trends in ΔNDVI can be discerned among the senescence months, with larger increasing trend found in July (1.8 × 10−4 yr−1) and September (2.1 × 10−4 yr−1), whereas decreasing trends were found for August (−3.7 × 10−5 yr−1) and October (−1.7 × 10−4 yr−1; Fig. 2a). There is also an increasing trend in ΔNDVI for the entire senescence period (July to October, JASO) (Fig. 2b), although noting this may be expected due to rising NDVImax under planetary greening (Fig. 2c). Dramatic regional variations, however, existed in ΔNDVI trends across the NH (Fig. 2d–i). Specifically, we found ΔNDVI increased in northwestern North America in July, which dominated the acceleration of autumnal senescence during the senescence period (Fig. 2d, h). In contrast, negative ΔNDVI trends were mainly distributed in northeastern North America and eastern Eurasia during August, which contributed to a slowing of the speed of autumn senescence (Fig. 2e, h). In September, contrasting changes in ΔNDVI were observed between the high and middle latitudes of Eurasia (Fig. 2f), resulting in opposite trends in the speed of autumn senescence (e.g., acceleration in middle Eurasia but deceleration in higher latitudes) (Fig. 2h). We also found that the speed of autumn senescence slowed down in the west and east of North America in October, when vegetation was already in dormant phase across most high latitudes (Fig. 2g).

a Temporal trends of ΔNDVI for senescence months in the Northern Hemisphere (NH) (recalling that a positive value of ΔNDVI corresponds to a decrease in NDVI). b Temporal trends of ΔNDVI over the senescence period (from July to October, JASO) and remaining NDVI at the end of the autumn senescence. c Temporal trends of the annual maximum NDVI (NDVImax). The bars inserted in a indicate trends in ΔNDVI for each month, the whole senescence period, and the remaining NDVI. d–i Spatial patterns of ΔNDVI trends during 1982–2018 for senescence months (d–g), the entire senescence period (h), and remaining NDVI (i). Black dots in d–i indicate significant trends at 0.05 level, while the inserted stacked bars indicate the frequency distributions of ΔNDVI trends.
Overall, ΔNDVI over the entire senescence period increased in 53.1% of the NH (significant in 17.9%, Fig. 2h), implying an overall acceleration of autumn senescence would occur in these areas. Although acceleration of senescence was found in more than half of the NH, the remaining NDVI (NDVI left in the month of EOS, see definition in Fig. 1 and Supplementary Table 1) still exhibited positive trends in 69.4% of the NH (Fig. 2i). These findings are anticipated due mainly to the apparent summer greening (i.e., a prevalent increase in NDVImax, Supplementary Fig. 1). Indeed, when averaged across NH, NDVImax increased significantly at a rate of 8.4 × 10−4 yr−1 (p < 0.001) during the senescence period (Fig. 2c). Meanwhile, ΔNDVI over JASO general increased, but its magnitude (2.1 × 10−4 yr−1, p > 0.05) is weaker than that of NDVImax. As a result, the remaining NDVI had an overall positive trend (6.3 × 10−4 yr−1, p < 0.001). Our analysis, therefore, reveals that changes in the speed of autumn senescence (ΔNDVI) and the peak greenness (NDVImax) can affect vegetation greenness at the later stage of the growing season.
Climatic regulation on the speed of autumn senescence
To investigate the climatic regulations on the speed of autumn senescence, we conducted partial correlation analyses to isolate the influence of insolation, wind speed, soil moisture, and temperature (Supplementary Fig. 2). The factor with the strongest partial correlation with ΔNDVI was defined as the dominant driver of autumn senescence (see Methods). Again, the dominant factors that accelerate or slow down the speed of autumn senescence were heterogeneous and discernible in their patterns over the NH (Supplementary Fig. 3).
Our noted strong spatial variations in ΔNDVI trends may be explained by the divergent biological responses of plants to climate change across different climatic regions. In fact, climatic regulations on ΔNDVI trends (higher values indicate a greater senescence speed, Fig. 1) were mainly associated with local hydrothermal conditions (Fig. 3). At the beginning of the autumn senescence period (that is, July), ΔNDVI correlated positively with temperature, but negatively with soil moisture in colder (mean annual temperature, MAT < 5 °C) and relatively drier (mean annual precipitation, MAP < 500 mm) regions (Fig. 3a, f). This finding indicated a possible acceleration of senescence caused by warming, because climatic warming may in turn lead to severe soil water deficits. Meanwhile, insolation was identified as a major promotor of the speed of autumn senescence in 33.2% of NH (Fig. 3k). Interestingly, the overall temperature effects on ΔNDVI shifted from positive (July) to negative (August, September, and October) as autumn senescence progresses. The negative temperature dominance on ΔNDVI trends over the NH increased from ~31% (August and September, Fig. 3l, m) to 46.9% (October, Fig. 3n). These results imply that a warming climate could accelerate autumn senescence in earlier stages of the senescence period, probably due to the detrimental summer extreme temperatures. On the other hand, it might significantly slow down the speed of senescence in its later stages by prolonging the exposure of vegetation to favorable summer climate.

a–e The distribution of dominant climatic factors was negatively correlated with ΔNDVI for the senescence months (a–d) and the remaining NDVI (e) in the climate space. f–j The distribution of dominant climatic factors was positively correlated with ΔNDVI for the senescence months (f–i) and the remaining NDVI (j) in the climate space. Each climate bin was defined by 0.5 °C intervals of mean annual temperature (MAT) and 20 mm intervals of mean annual precipitation (MAP). The factor that appeared most frequently for each bin was presented in these figures. k–o The percentages of pixels occupied by dominant climatic factors, which positively (left bars, add to 100%) and negatively (right bars, add to 100%) correlated with ΔNDVI for senescence months (k–n) and the remaining NDVI (o). Note that the positive correlation with ΔNDVI or the negative correlation with the remaining NDVI indicates that this factor can accelerate the speed of autumn senescence, and vice versa.
Unlike such reversal of temperature regulation on the speed of autumn senescence, the effects of insolation, soil moisture and windspeed remained almost consistent along the senescence period. To be specific, insolation was identified as the positive dominant factor of ΔNDVI, particularly over the warmer regions (MAT > −5 °C) and the lower latitudes (Fig. 3). Because the increasing insolation might raise the risk of leaf photo-oxidative damage, which could promote the progress of leaf senescence19. As for soil moisture and wind speed, their influence is only apparent in a few sporadic areas.
While for the remaining NDVI (higher values indicate slower senescence speed, Fig. 1), it was generally positively related to temperature and insolation, but negatively related to wind speed (Fig. 3e, j, o). The positive temperature effect was prevalently observed over the NH (Supplementary Figs. 2x, 3f), which confirmed again that warming climate contribute to slow down the speed of senescence in late autumn. Followed by insolation, its positive and dominant effects on the remaining NDVI were mainly distributed in North Siberia and North America (with MAT < 0 °C) (Supplementary Figs. 2e, 3e), indicating the negative light regulation on leaf senescence was stronger than that of warming for these regions20. We also found that the wind speed exerted negative controls on the remaining NDVI in 34.3% of NH (Fig. 3e, o) through its role in promoting leaf abscission and intensifying drought stress by increasing evapotranspiration7. The influence of soil moisture expressed a contrasting regional difference (Supplementary Fig. 2q). More specifically, in terms of inland Eurasia and North America, the remaining NDVI was positively regulated by soil moisture (Supplementary Fig. 2q), indicating that increases in soil moisture can slow down the speed of autumn senescence in these water-limited ecosystems. However, in high-latitude Siberia (>50°N), soil moisture was negatively correlated with the remaining NDVI (Supplementary Fig. 2q), mainly due to the anoxia conditions under high soil moisture21 and limited nutrient availability in these regions of permafrost6.
The effect of summer greening on the extraction of autumn phenology
Parallel to climate change, the observed summer greening (that is, an increase of 8.4 × 10−4 yr−1 in NDVImax, p < 0.001, Fig. 2c) could modulate the extracted autumn phenology and its temporal trend. We hypothesize that, accompanied with summer greening, the increasing mature leaves in the peak season would contribute to an increasing trend in ΔNDVI, thus creating an illusion of faster senescence (Fig. 4a). Such effect could be greatly reduced by replacing monthly senescence (ΔNDVI) with the relative decrease in NDVI (δNDVI = ΔNDVI/NDVImax). Summer greening, in a similar way, would contribute to delayed EOS calculated from fixed NDVI thresholds (EOSa) than that from fixed percentage of NDVImax (EOSp), implying the influence of changing NDVImax in EOS detection (Fig. 4b, also see definitions in Supplementary Table. 1).

a For each senescence month, monthly senescence is proxied by the decrease in NDVI (ΔNDVI) or percentage of decrease in NDVI relative to NDVImax (δNDVI = ΔNDVI/NDVImax). Peak of season (POS) greening would lead to an apparent increase in ΔNDVI even if δNDVI stays invariant. b In correspondence to ΔNDVI and δNDVI, end of season can be determined by fixed NDVI thresholds (EOSa) or fixed percentage of NDVImax (EOSp). If NDVImax were kept unchanged, these two methods would deliver the same EOS, whereas increases in NDVImax would results to a later EOSa than EOSp.
Notably, trends in δNDVI generally presented similar spatial patterns as that of ΔNDVI in each of the senescence months (Supplementary Fig. 4a–e). After separating the contribution of trends in NDVImax and δNDVI to trends in ΔNDVI (Supplementary Fig. 5a–e, see Methods), we found that the increase in NDVImax accounted for ~40% of ΔNDVI trends in NDVI for parts of Siberia and central Eurasia in August and September (Supplementary Fig. 5b, c). Despite this, the overall contribution of NDVImax to ΔNDVI trends was marginal for other regions and periods (e.g., only 1.7, 5.9, 5.0 and 2.6% for Jul, Aug, Sep and Oct, respectively). Such influence was confirmed by the relatively small portion of significant correlations between trends in NDVImax and ΔNDVI for these months (Supplementary Fig. 6a–d). Besides, similar spatial patterns of dominant climatic factors were also observed in δNDVI, further indicating their tight connection (Supplementary Fig. 3).
We also found that the influence of increasing NDVImax on trends of the remaining NDVI was apparent (36.7% on average, Supplementary Fig. 5f), which was in line with the widespread significant correlations between trends in NDVImax and the remaining NDVI over the NH (Supplementary Fig. 6f). This leads to higher fractions of positive trend areas for the remaining NDVI (69.4% with 30.1% significant, Fig. 2i) than its percentage-based counterpart (55.3% with 14.7% significant, Supplementary Fig. 4f). Obviously, the impact of conspicuous summer greening on the remaining NDVI may have cascaded effects on the extraction of EOS.
We therefore further compared the temporal trends of EOSa and EOSp (definitions in Fig. 4b, also see definitions in Supplementary Table. 1) during the period 1982–2018. Across the NH, we found areas occupied with delayed EOSa (60.2%, significant in 22.2%, Fig. 5a) exceeds that of EOSp (51.1%, significant in 15.1%, Fig. 5b). As a result, an overall delayed trend of EOSa (0.6 days dec−1) but advanced EOSp (−0.4 days dec−1) trend was presented in NH (Fig. 5a–c). This divergence summed up to 1.0 days dec−1 NDVImax-induced delay in EOSa. Given that such an advance in autumn senescence was supported by a meta-analysis based on field observations22, we reckon that EOSp may be a better indicator of autumnal leaf phenology, which captures the physiological changes of plants. EOSa, carrying the additional effect of NDVImax, my instead, serve as a better indicator for canopy greenness browning. Our results thus infer that the increase in the number of mature leaves in the canopy under summer greening may counteract the intrinsic changes in autumnal leaf senescence.

a, b Spatial patterns of trends in EOSa and EOSp in 1982–2018. Black dots indicate a significant trend at the 0.05 level. The stacked inserted bars indicate the frequency distributions of trends in EOSa and EOSp. c Difference between trends in EOSa and EOSp (trends in EOSa minus trends in EOSp). d Contribution of trends in EOSp (green bars) and NDVImax (i.e., EOSa-EOSp, orange bars) to trends in EOSa (purple bars) for different vegetation types (see Methods). MF mixed forests, DBF deciduous broadleaf forests, DNF deciduous needleleaf forests.
Among the studied plant functional types (PFTs), we also observed clear mismatch between trends in EOSa and EOSp (Fig. 5d). In line with the NH average (a combination of all PFTs), we found the opposite contributions of NDVImax (delay) and EOSp (advance) to EOSa trends in grasslands, savannas and shrublands in boreal and arctic regions. However, the delaying effect of summer greening could hardly compensate for the large advancement in EOSp, as these ecosystems generally locate in resource-limited areas (Supplementary Fig. 7). Growth enhancement (greening) during the earlier season may prematurely consume water and nitrogen, which would in turn speed up the progress of vegetation senescence and cause earlier end of growing season23,24,25. Indeed, the largest advancement in EOSp was observed in grasslands of central Eurasia, which were characterized with severe water limitations and significant vegetation greening (Supplementary Fig. 1).
The observed delay in EOSp for the woody species could be ascribed to the higher resource availability in their relatively wetter and fertile habitants, and their ability to accumulate large nutrient and carbohydrate reserves in autumn26,27. The increase in NDVImax contribute to later EOSa than EOSp, albeit varied across these PFTs (Fig. 5d). We quantified the contribution of increasing NDVImax to the delayed EOSa through the residue proportion of trends after subtracting EOSp from EOSa (i.e., the proportion of orange bars within the transparent purple bars, last section of methods). As a result, the increase in NDVImax contributed to more than half of the delay in EOSa for woody savannas (69.2%) and temperate shrublands (88.5%). However, such influence is minor in forests, contributing to 20.3% and 9.5% in mixed forests and deciduous broadleaf forests, respectively. On the contrary, the decrease in NDVImax in deciduous needleleaf forests advanced EOSa by 29.3%, comparing to EOSp.
Remarkably, NDVImax-induced divergence between EOSa and EOSp trends exerted strong controls on their responses to climate change (Supplementary Figs. 2, 8). Despite the observed clear positive temperature dominance for EOSa and EOSp (Supplementary Fig. 3f, r), the difference in the sensitivities of EOSa and EOSp to temperature was examined (e.g., EOSa (γa) and EOSp (γp), see Methods). The partial least squares regressions analysis suggested that both EOSa and EOSp had positive temperature sensitivities in most areas, while negative sensitivities were concentrated in xeric grasslands of central Eurasia and USA (Supplementary Fig. 9a, b). Note that EOSa presented larger temperature sensitivities than EOSp in 69.5% of northern areas, e.g., γa (2.1 days °C−1) was more than 50% higher than γp (1.3 days °C−1) when averaged over the NH (Supplementary Fig. 9c), due mainly to the additional NDVImax greening trend carried by EOSa. Indeed, the prominent differences between γa and γp were discovered in grasslands and temperate shrublands (Supplementary Fig. 9d), where EOSa trends were predominantly contributed by changes in NDVImax (Fig. 5d).
Implications for model benchmarking
The mismatch between phenology metrics based on absolute (ΔNDVI, EOSa) and relative (δNDVI, EOSp) changes in NDVI indicates the considerable effect of summer greening on the extraction of autumn phenology. In other word, the increase in the number of mature leaves in canopy under summer greening might confound the actual changes in the timing of leaf senescence. Such an effect is most clearly shown by the mismatch between EOSa and EOSp. EOSa, derived from fixed thresholds of NDVI, is a direct reflection of drop in canopy greenness, with a delayed trend in EOSa indicating the prolonged canopy browning process. EOSp, derived from fixed percentages of NDVImax, instead, better captures senescence of leaves under the overall changes in canopy greenness. As shown in our results, the widely observed global greening (Supplementary Fig. 1) in this study and previous studies28,29, has, indeed, counteracted the intrinsic advance of leaf senescence by an apparent delay in canopy browning. Despite that these two metrics may represent phenology at different ecological levels (e.g., leaf vs. canopy), leaf phenology is more closely related to the time when plants enter winter dormancy and is a key regulator of vegetation dynamics and terrestrial carbon sequestration2. The absence of eliminating the summer greening effect in the extraction of autumn phenology (i.e., threshold-based methods: EOSa) may introduce bias to our understanding of the leaf senescence process and consequently, the vegetation-climate interactions and the terrestrial carbon cycle. Since the timing of leaf senescence is crucial for reconstructing historical plant carbon uptake and predicting its future changes in Earth system models, it is essential to update phenology modules embedded in state-of-the-art land surface schemes using accurate representations of these processes. This will enable a better simulation of the terrestrial carbon cycle30,31.
However, leaf senescence so far is one of the most difficult processes to be parameterized in terrestrial ecosystem models, which is restricted by our incomplete understandings of its complex mechanisms. To our knowledge, DGVMs coupled with land surface models generally determine the onset of plant dormancy by tracking the thresholds of modelled or prescribed leaf area index30. For example, in the ORCHIDEE model, it is defined as the date when the modelled leaf area index (LAI) drops below 0.232. Evidence from our study suggests that fixed thresholds, however, may not accurately reproduce the timings of autumn leaf phenology when the summer greening effect is not thoroughly considered. Other models, for example, the Ecosystem Demography model version 2 (ED2), used the prescribed satellite-derived EOS estimates to parameterize the date of leaf drop during the year33. In this case, an accurate estimation of EOS, especially taking the effect of summer greening into consideration, is of great importance for accurate model parameterizations. Moreover, a large body of models simulates the timing of leaf shed by a function of air temperature and (or) day length34,35,36. Although these models directly simulate leaf phenology, caution is still needed for benchmarking models against satellite-derived results, since phenology based on absolute changes in vegetation greenness may be not sufficient to characterize the trajectories of leaf senescence.
We also found that EOSa, carrying the additional greening trend at POS, generally exhibits higher temperature sensitivities than EOSp. This discrepancy was in support of findings in Zhang et al. (2020), who reported that the response of greenness-based phenology to climate change was greater than that of photosynthesis-based phenology. Therefore, if the apparent sensitivity of satellite-based leaf phenology (with the absence of a summer greening effect in the extraction of phenology) is used to parameterize leaf behavior in DGVMs, the future carbon sink potential of terrestrial ecosystems might be overestimated.
Our study provides a novel angle on the speed of autumn senescence and its response to climate change over the NH. We find that warming climate accelerates senescence in July, but this influence usually reversed in later summer and early autumn. Insolation generally accelerates senescence across the entire senescence period. Our analysis also verifies a summer greening (increase in NDVImax)-induced delay in EOS, as canopy greening may counteract the intrinsic changes in leaf phenology, and lead to higher temperature sensitivities for EOS carrying the additionally greening trend. Global warming and increasing atmospheric CO2 have been reported to continually contribute to earth greening, especially in NH28,29. Our analysis thus allows us to reconsider the linkage between changes in apparent canopy greenness and changes in intrinsic leaf physiology, which could provide an important benchmark for those developing DGVMs for use in ESMs.
Methods
Datasets
In this study, NDVI during the 1982–2018 period was extracted from the third-generation product of NASA’s Global Inventory Modeling and Mapping Studies (GIMMS) group37. Various issues, such as orbital drift, calibration, viewing geometry and volcanic aerosols, have been corrected in this product38. This dataset consists of fortnightly NDVI observations at a spatial resolution of 0.083° (~8 km). The monthly NDVI data was estimated through the maximum composition of each month’s two records. In this analysis, we screened out pixels dominated by cropland and evergreen forests (inferred from the MODIS Landcover Product, MCD12C1, IGBP classification)39, because their seasonal cycles are unclear. Consistent with the NDVI time span, the monthly mean 2-m surface temperature was derived from CRU.TS.4.05 with 0.5° spatial resolution40. Monthly insolation was extracted and summed from CRU-JRA v2.2 with 0.5° spatial and 6-h temporal resolution41. The monthly surface soil moisture was acquired from the C3S dataset provided by the European Center for Medium-Range Weather Forecasts (ECMWF) with a 0.25° spatial resolution, which was then interpolated to 0.5° × 0.5°. The monthly wind speed was obtained from the ERA5 reanalysis data at a spatial resolution of 0.25° 42 and was interpolated to 0.5° × 0.5°.
Phenology extraction methods
Our study was conducted across north of 30°N (excluding the subtropical regions), where plants have clear seasonal variations and are capable to extract phenological metrics. The Autumn senescence period is defined as the time window between POS (Supplementary Fig. 10) and EOS (Supplementary Fig. 11). The decrease in NDVI (ΔNDVI) during the entire senescence period was defined as the difference in NDVI between POS month and EOS month. Similarly, the monthly ΔNDVI was the NDVI difference between the current and the following month. The remaining NDVI i.e., NDVI at EOS month, refers to the remaining greenness in the final stage of autumn senescence. For the minor regions with later POS (e.g., later than July, only 6.2% of the study area), the calculation of ΔNDVI also starts from POS month, NDVI data prior to the POS month was excluded from the analysis.
Besides monthly senescence, we further obtained threshold-based EOS (EOSa). To avoid the influence of potential snow cover on EOS extraction, we first used the spline function to interpolate the monthly temperature to a daily basis, and then obtained the boundary of the thermal growing season by a sequence of five days below 0 °C. Before EOS extraction, NDVI pixels outside this thermal growing season were replaced by that of the temporally nearest snow-free date. We than determined EOSa according to the following steps (Polyfit-Maximum method43; Supplementary Fig. 12): (1) calculate the averaged NDVI curves during the past 37-year (1982–2018), (2) estimate the rates of changes in the average NDVI curves as: NDVIratio(t)=[NDVI(t + 1)-NDVI(t)]/NDVI(t), (3) determine the time T when the minimum NDVIratio occurred, (4) use the NDVI at time T (i.e., NDVI(T)) as the threshold for EOSa extraction, (5) apply the threshold to determine EOSa for each year. The Polyfit-maximum method have been successfully used in many of previous phenological studies44,45,46, and its outcomes are comparable with other commonly used methods6. The priorly determined NDVI threshold also gives us the chance to correspondently obtain a percentage-based threshold, which is then used for further analysis (see detection of EOSp in next paragraph).
The extraction of ΔNDVI and EOSa is probably confounded by global greening, especially in the context of the prominent increase in NDVImax (i.e., the effect of summer canopy greening, Fig. 4 and Supplementary Fig. 1). To reduce such an impact, we employed another method based on the percentage of NDVImax. During each month and the entire senescence period, we calculated the percentage of decrease in NDVI compared to the peak season NDVI as δNDVI = ΔNDVI/NDVImax. Similarly, the percentage of remaining NDVI relative to NDVImax was calculated as the division between the remaining NDVI and NDVImax. The percentage-based EOS (EOSp) was extracted from almost the same steps as EOSa, except that the percentage of NDVI(T) relative to NDVImax (NDVI(T)/NDVImax) were used as thresholds to detect yearly EOSP (step 4, Supplementary Fig. 12). The differences in ΔNDVI and δNDVI, and EOSa and EOSp gives us chance to investigate the effect of NDVImax on autumn phenology detection (see last section of methods).
Climatic regulations on the process of autumn senescence
Benefiting from previous studies, we examined the effects of four climatic factors i.e., insolation, wind speed, soil moisture, and temperature on the leaf senescence process6,7,14,47. Note that we did not take precipitation into consideration, due mainly to the following aspects: (1) Soil moisture is extensively correlated with precipitation in the study area (Supplementary Fig. 13) and (2) soil moisture is more closely and directly related to plant water uptake than precipitation5,48. The lagged effect of climate on vegetation dynamics was also considered for each of the phenological matrices (ΔNDVI, δNDVI, EOSa and EOSp) following previous studies6,8,15. Firstly, simple correlations were used to determine the preseason length of each climatic factors, which was defined as the period when the mean value of factors having the largest absolute simple correlation with phenological metrics (Supplementary Figs. 14, 15). The preseason length ranged from the month of phenology metrices to its preceding 3 months, with a monthly time step. The preseason was defined for each of the four phenological metrics and climatic variables. We then performed partial correlation analysis between each of the phenological metrices and climatic factors during the preseason (Supplementary Figs. 2, 8). It enables us to explore the relationship between plant phenology and single climatic factor while eliminating the effects of the remaining factors. The climatic factor with the largest positive (lowest negative) partial correlation coefficient was defined as the positive (negative) dominant factor (Supplementary Fig. 3).
Temperature sensitivity of EOS
Our partial correlation analysis suggested distinct temperature dominances on EOSa and EOSp, while similar patterns were not clear for ΔNDVI and δNDVI. We then performed a partial least squares regression between EOSa (also EOSp) and the four climatic factors, and assigned the coefficient of temperature as the temperature sensitivity of EOSa (γa) and for EOSp (γb). The coefficient was commonly used as a proxy of changes in phenological dates per unit increase in temperature (i.e., temperature sensitivity)49,50,51.
Separation of the effects of NDVImax in ΔNDVI and EOSa
In this study, ΔNDVI for each month of senescence is also expressed as:
This formula inherently indicates that ΔNDVI is jointly determined by δNDVI and NDVImax. Trends of ΔNDVI during 1982–2018, therefore, can be separated into the trends of δNDVI and NDVImax based on the multiplication derivation rule ((uv)’=u’v + v’u, u and v are variables). The following approximation was used:
where tr[ΔNDVI], tr[δNDVI] and tr[NDVImax] are trends of ΔNDVI, δNDVI and NDVImax during the period 1982–2018, respectively. mean[NDVImax] and mean[δNDVI] are the multi-year averages of NDVImax and δNDVI. tr[δNDVI] x mean[NDVImax] and tr[NDVImax] x mean[δNDVI] thus denote the contribution of δNDVI trends and NDVImax trends to ΔNDVI trends, respectively. To test the reliability of such approximation, we additionally checked whether the sum of the terms on the right-hand side of Eq. 2 (i.e., the partitioning approach) is equivalent to the left-hand side. The feasibility of such partitioning is ensured by a perfect 1:1 distribution of values on both side of Eq. 2 (Supplementary Fig. 16).
As for the EOS, once the thresholds of NDVI(T) and NDVI(T)/NDVImax were defined from the multi-year averaged NDVI curve (i.e., step 1 to step 4; Supplementary Fig. 12) the determination of EOSa and EOSp for each year (step 5, based on the prescribed thresholds) is less likely to be influenced by the peak times or decrease speed of NDVI. For example, if NDVImax keeps constant (e.g., 0.8), NDVI(T) for EOSa is 0.2 and the NDVI(T)/NDVImax for EOSp is 25%. No matter how the peak time and decrease speed changes, the time when NDVI value decrease to 0.2 always coincides with the time when a relative decrease to 25% occurred (Supplementary Fig. 17). Therefore, we can conclude that if NDVImax stays the same, EOSa and EOSp should be equal. Difference in EOSa and EOSp can thus be used to indicate asynchronization of EOS resulted from the increasing NDVImax.
Data availability
All observational data sets that we used are publicly available. The GIMMS3g NDVI data sets covering the period 1982–2015 are publicly available at https://data.tpdc.ac.cn/en/data/9775f2b4-7370-4e5e-a537-3482c9a83d88/, datafiles covering the period 2016–2018 were directly requested from J. Tucker, the creator of GIMMS NDVI3g dataset. The CRU.TS.4.05 mean 2-m surface temperature data sets are available at https://data.ceda.ac.uk/badc/cru/data/cru_ts/cru_ts_4.05. The CRU.JRA v2.2 6-h insolation data sets are available at https://data.ceda.ac.uk/badc/cru/data/cru_jra/cru_jra_2.2. The C3S soil moisture data sets are available at https://cds.climate.copernicus.eu/cdsapp#!/dataset/satellite-soil-moisture?tab=overview. The ERA5 reanalysis wind speed data sets are available at https://cds.climate.copernicus.eu/cdsapp#!/dataset/reanalysis-era5-single-levels-monthly-means?tab=overview.
Code availability
All the analyses and figures are made using MATLAB R2020a and Power Point 2019, and the codes are available from the corresponding author.
References
Piao, S. et al. Plant phenology and global climate change: Current progresses and challenges. Glob Change Biol 25, 1922–1940 (2019).
Piao, S. et al. Net carbon dioxide losses of northern ecosystems in response to autumn warming. Nature 451, 49–52 (2008).
Gallinat, A. S., Primack, R. B. & Wagner, D. L. Autumn, the neglected season in climate change research. Trends Ecol Evol 30, 169–176 (2015).
Chen, L. et al. Leaf senescence exhibits stronger climatic responses during warm than during cold autumns. Nat Clim Chang 10, 777–780 (2020).
Wang, X., Wu, C., Liu, Y., Peñuelas, J. & Peng, J. Earlier leaf senescence dates are constrained by soil moisture. Glob Change Biol 29, 1557–1573 (2022).
Liu, Q. et al. Delayed autumn phenology in the Northern Hemisphere is related to change in both climate and spring phenology. Glob Change Biol 22, 3702–3711 (2016).
Wu, C. et al. Widespread decline in winds delayed autumn foliar senescence over high latitudes. Proceedings of the National Academy of Sciences 118, https://doi.org/10.1073/pnas.2015821118 (2021).
Wu, C. et al. Increased drought effects on the phenology of autumn leaf senescence. Nat Clim Chang 12, 943–949 (2022).
Yu, H., Zhou, G., Lv, X., He, Q. & Zhou, M. Environmental factors rather than productivity drive autumn leaf senescence: evidence from a grassland in situ simulation experiment. Agric For Meteorol 327, https://doi.org/10.1016/j.agrformet.2022.109221 (2022).
Estiarte, M. & Peñuelas, J. Alteration of the phenology of leaf senescence and fall in winter deciduous species by climate change: effects on nutrient proficiency. Glob Change Biol 21, 1005–1017 (2015).
Keskitalo, J., Bergquist, G., Gardeström, P. & Jansson, S. A Cellular Timetable of Autumn Senescence. Plant Physiol 139, 1635–1648 (2005).
Lim, P. O., Kim, H. J. & Gil Nam, H. Leaf Senescence. Annu Rev Plant Biol 58, 115–136 (2007).
Deslauriers, A., Rossi, S. & Sevanto, S. Metabolic memory in the phenological events of plants: looking beyond climatic factors. Tree Physiol 39, 1272–1276 (2019).
Fracheboud, Y. et al. The Control of Autumn Senescence in European Aspen. Plant Physiol 149, 1982–1991 (2009).
Piao, S., Wang, J., Li, X., Xu, H. & Zhang, Y. Spatio‐temporal changes in the speed of canopy development and senescence in temperate China. Glob Change Biol 28, 7366–7375 (2022).
Vitasse, Y., Bresson, C. C., Kremer, A., Michalet, R. & Delzon, S. Quantifying phenological plasticity to temperature in two temperate tree species. Funct Ecol 24, 1211–1218 (2010).
Anav, A. et al. Evaluation of Land Surface Models in Reproducing Satellite Derived Leaf Area Index over the High-Latitude Northern Hemisphere. Part II: Earth System Models. Remote Sens 5, 3637–3661 (2013).
Murray-Tortarolo, G. et al. Evaluation of Land Surface Models in Reproducing Satellite-Derived LAI over the High-Latitude Northern Hemisphere. Part I: Uncoupled DGVMs. Remote Sens 5, 4819–4838 (2013).
Wu, Z. et al. Atmospheric brightening counteracts warming‐induced delays in autumn phenology of temperate trees in Europe. Glob Ecol Biogeogr 30, 2477–2487 (2021).
Zhang, Y., Commane, R., Zhou, S., Williams, A. P. & Gentine, P. Light limitation regulates the response of autumn terrestrial carbon uptake to warming. Nat Clim Chang 10, 739–743 (2020).
Mainiero, R. & Kazda, M. Effects of Carex rostrata on soil oxygen in relation to soil moisture. Plant Soil 270, 311–320 (2005).
Liu, H. et al. Phenological mismatches between above- and belowground plant responses to climate warming. Nat Clim Chang 12, 97–102 (2021).
Leuzinger, S., Zotz, G., Asshoff, R. & Korner, C. Responses of deciduous forest trees to severe drought in Central Europe. Tree Physiol 25, 641–650 (2005).
Lian, X. et al. Summer soil drying exacerbated by earlier spring greening of northern vegetation. Sci Adv 6, https://doi.org/10.1126/sciadv.aax0255 (2020).
Zhang, Y., Keenan, T. F. & Zhou, S. Exacerbated drought impacts on global ecosystems due to structural overshoot. Nat Ecol Evol 5, 1490–1498 (2021).
Dietze, M. C. et al. Nonstructural Carbon in Woody Plants. Annu Rev Plant Biol 65, 667–687 (2014).
Fu, Y. S. H. et al. Variation in leaf flushing date influences autumnal senescence and next year’s flushing date in two temperate tree species. Proceedings of the National Academy of Sciences 111, 7355–7360 (2014).
Piao, S. et al. Characteristics, drivers and feedbacks of global greening. Nat Rev Earth Environ 1, 14–27 (2019).
Zhu, Z. et al. Greening of the Earth and its drivers. Nat Clim Chang 6, 791–795 (2016).
Richardson, A. D. et al. Terrestrial biosphere models need better representation of vegetation phenology: results from the North American Carbon Program Site Synthesis. Glob Change Biol 18, 566–584 (2012).
Keenan, T. F. et al. Net carbon uptake has increased through warming-induced changes in temperate forest phenology. Nat Clim Chang 4, 598–604 (2014).
Krinner, G. et al. A dynamic global vegetation model for studies of the coupled atmosphere-biosphere system. Glob Biogeochem Cycle 19, https://doi.org/10.1029/2003GB002199 (2005).
Medvigy, D., Wofsy, S. C., Munger, J. W., Hollinger, D. Y. & Moorcroft, P. R. Mechanistic scaling of ecosystem function and dynamics in space and time: Ecosystem Demography model version 2. J Geophys Res 114, https://doi.org/10.1029/2008JG000812 (2009).
Caffarra, A., Donnelly, A. & Chuine, I. Modelling the timing of Betula pubescens budburst. II. Integrating complex effects of photoperiod into process-based models. Clim Res 46, 159–170 (2011).
Dufrêne, E. et al. Modelling carbon and water cycles in a beech forest. Ecol Model 185, 407–436 (2005).
Jeong, S. J. & Medvigy, D. Macroscale prediction of autumn leaf coloration throughout the continental United States. Glob Ecol Biogeogr 23, 1245–1254 (2014).
Tucker, C. J. et al. An extended AVHRR 8‐km NDVI dataset compatible with MODIS and SPOT vegetation NDVI data. Int J Remote Sens 26, 4485–4498 (2010).
Kaufmann, R. K. et al. Effect of orbital drift and sensor changes on the time series of AVHRR vegetation index data. IEEE Trans Geosci Remote Sensing 38, 2584–2597 (2000).
Friedl, M. A. et al. MODIS Collection 5 global land cover: Algorithm refinements and characterization of new datasets. Remote Sens Environ 114, 168–182 (2010).
Harris, I., Osborn, T. J., Jones, P. & Lister, D. Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Sci Data 7, https://doi.org/10.1038/s41597-020-0453-3 (2020).
Kobayashi, S. et al. The JRA-55 Reanalysis: General Specifications and Basic Characteristics. Journal of the Meteorological Society of Japan Ser II 93, 5–48 (2015).
Bell, B. et al. The ERA5 global reanalysis: Preliminary extension to 1950. Q J R Meteorol Soc 147, 4186–4227 (2021).
Piao, S., Fang, J., Zhou, L., Ciais, P. & Zhu, B. Variations in satellite-derived phenology in China’s temperate vegetation. Glob Change Biol 12, 672–685 (2006).
Jeong, S. J., Ho, C. H., Gim, H. J. & Brown, M. E. Phenology shifts at start vs. end of growing season in temperate vegetation over the Northern Hemisphere for the period 1982-2008. Glob Change Biol 17, 2385–2399 (2011).
White, M. A. et al. Intercomparison, interpretation, and assessment of spring phenology in North America estimated from remote sensing for 1982-2006. Glob Change Biol 15, 2335–2359 (2009).
Cong, N. et al. Changes in satellite-derived spring vegetation green-up date and its linkage to climate in China from 1982 to 2010: a multimethod analysis. Glob Change Biol 19, 881–891 (2013).
Gill, A. L. et al. Changes in autumn senescence in northern hemisphere deciduous trees: a meta-analysis of autumn phenology studies. Ann Bot 116, 875–888 (2015).
Liu, L. et al. Soil moisture dominates dryness stress on ecosystem production globally. Nat Commun 11, https://doi.org/10.1038/s41467-020-18631-1 (2020).
Fu, Y. H. et al. Declining global warming effects on the phenology of spring leaf unfolding. Nature 526, 104–107 (2015).
Menzel, A. et al. European phenological response to climate change matches the warming pattern. Glob Change Biol 12, 1969–1976 (2006).
Yang, Y., Guan, H., Shen, M., Liang, W. & Jiang, L. Changes in autumn vegetation dormancy onset date and the climate controls across temperate ecosystems in China from 1982 to 2010. Glob Change Biol 21, 652–665 (2015).
Acknowledgements
This study was supported by the National Natural Science Foundation of China (41988101).
Author information
Authors and Affiliations
Contributions
S.L.P., Y.C.Z., S.B.H. and Q.L. designed the research; Y.C.Z. performed analysis; Y.C.Z. wrote the first draft of the manuscript. C.H., J.P., S.R. and R.B.M. contributed to the interpretation of the results and to the text.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Earth & Environment thanks the anonymous reviewers for their contribution to the peer review of this work. Primary Handling Editor: Aliénor Lavergne.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhang, Y., Hong, S., Liu, Q. et al. Autumn canopy senescence has slowed down with global warming since the 1980s in the Northern Hemisphere. Commun Earth Environ 4, 173 (2023). https://doi.org/10.1038/s43247-023-00835-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s43247-023-00835-0
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
