
Abstract
Greenspaces are important for sustaining healthy urban environments and their human populations. Yet their capacity to support multiple ecosystem services simultaneously (multiservices) compared with nearby natural ecosystems remains virtually unknown. We conducted a global field survey in 56 urban areas to investigate the influence of urban greenspaces on 23 soil and plant attributes and compared them with nearby natural environments. We show that, in general, urban greenspaces and nearby natural areas support similar levels of soil multiservices, with only six of 23 attributes (available phosphorus, water holding capacity, water respiration, plant cover, arbuscular mycorrhizal fungi (AMF), and arachnid richness) significantly greater in greenspaces, and one (available ammonium) greater in natural areas. Further analyses showed that, although natural areas and urban greenspaces delivered a similar number of services at low (>25% threshold) and moderate (>50%) levels of functioning, natural systems supported significantly more functions at high (>75%) levels of functioning. Management practices (mowing) played an important role in explaining urban ecosystem services, but there were no effects of fertilisation or irrigation. Some services declined with increasing site size, for both greenspaces and natural areas. Our work highlights the fact that urban greenspaces are more similar to natural environments than previously reported and underscores the importance of managing urban greenspaces not only for their social and recreational values, but for supporting multiple ecosystem services on which soils and human well-being depends.
Similar content being viewed by others
Introduction
Marked changes have occurred in urban areas over the past century due to the increasing concentration of people in large cities. Currently, more than half of the world’s population lives in cities, placing increasing stress on urban environments, their residents and infrastructure1. This urban population is expected to increase to about 68% by 20502, increasing the need to protect and improve the quality and accessibility of urban greenspaces, which include city parks, residential gardens and roadside verges. The health and social benefits of urban greenspaces are increasingly important for promoting social and physical wellbeing, and for alleviating the negative stresses associated with living in large urban centres3,4,5,6,7,8. Globally, there is increasing recognition by state and city planning bodies of the importance of ecosystem services both in urban and semi-natural environments9. More recent work aims to improve the extent to which land use decisions affect the provision of ecosystem services and therefore human well-being10. Greenspace soils support a wide range of ecosystem services such as plant productivity, habitat for soil organisms, nutrient availability for plants and microbes, plant-soil symbiosis, pathogen control, and water regulation. All of these services are critical for providing habitat, ameliorating urban heat island effects, and sustaining plant growth11,12,13,14,15.
Despite their importance, we have a relatively basic understanding of the capacity of urban greenspaces to support multiple ecosystem services compared with natural areas. Such a comparison is important because there is a general perception that greenspaces are depauperate and degraded, compared with natural areas, despite the fact that they often support distinct plant and animal communities16. Further, in order to urban planning as urban areas expand, it is critical to understand whether greenspaces provide similar environmental quality and psychological well-being as natural areas17. Our poor understanding of the relative effects of greenspaces compared with natural and semi-natural area effects is likely due to three reasons. First, most studies of urban greenspaces have tended to focus on single services such as carbon sequestration18 or microbial diversity19, yet studies of multiple ecosystem services (hereafter ‘multiservices’) have rarely been considered. This is important in highly managed systems such as urban greenspaces where existing levels of specific soil properties may be a direct consequence of artificial fertilisation rather than a residual signature of the original ecosystem. Second, most studies of urban greenspaces have been conducted at the local scale, for example, within specific parks20 or cities21,22,23,24, often without due recognition of the way in which they evolved across continents. Similarly, urban greenspaces vary markedly in their size and age, such that smaller sites may be more heavily used or managed, have larger edge effects, and therefore a lower capacity to support multiple services than larger sites. Older greenspaces may also have lower levels of services because they have been subjected to a longer period of management e.g., fertilisation and irrigation13. Different styles of greenspace management (mowing, fertilising, irrigation) would also likely affect the extent of services by affecting plant growth rates, litter removal and soil nutrient levels13,14. Thus, considering the size and age of greenspaces is important if we are to understand how their soils function. Finally, there are relatively few studies that directly compare urban greenspaces and their nearby natural analogues. Urban ecosystems are often regarded as highly disturbed environments that have been developed on artificial substrates. However, they range from semi-natural (wild) areas that are less intensively managed, to carefully manicured and extensively modified lawns and gardens25,26,27. City parks, for example, have highly compacted and fertilised soils, are often irrigated, frequently mown, and dominated by exotic turf grasses28,29. Yet, in many cases, urban greenspaces are on land that was formerly natural forest, or soils that supported natural ecosystems. Consequently, urban greenspaces may support levels of soil ecosystem services that more closely resemble those from natural ecosystems than previously expected. Overall, we know relatively little about the extent to which greenspaces support similar levels of services provided by soils, including multiple services (multiservices) that exist in natural ecosystem, and whether any effects vary among different global cities with different levels of management, age or size. This knowledge is needed if we are to manage greenspaces so that they continue to provide a range of ecosystem services under the pressure of increasing urban growth.
Here, we examined differences in eight soil ecosystem services, and their average value, and 23 separate attributes at 56 greenspace-natural paired sites, and sought to explore the major factors associated with the differences between greenspaces and adjacent natural areas. Such a study gives us the opportunity to assess the value of greenspaces as functional soil islands within a matrix of urban development and provides valuable insights into the extent to which management actions might be required to maintain their ability to provide services at a level similar to natural systems from which they were derived.
Results
Greenspaces and natural areas support similar levels of ecosystem multiservices
We found no difference in average ecosystem multiservices between greenspaces and nearby natural areas assessed using either unweighted (average) or weighted approaches (Fig. 1b). Values of unweighted and weighted approaches were highly correlated (Supplementary Fig. 1). We also investigated the capacity of natural systems and urban greenspaces to deliver multiple ecosystem services over a functional threshold. The multi-threshold approach indicated that natural areas and greenspaces delivered a similar number of services at low (25% threshold) and moderate (50%) levels of functioning, but natural systems delivered about 53% more services when operating at higher levels of functioning (75%, Fig. 2). Analyses at the individual soil service level further revealed that nutrient availability and plant-soil symbiosis were significantly greater in greenspaces. Pest control was greater in natural areas, but there were no differences in the other services between land uses (Fig. 1b). When we examined individual attributes, we found that values of available phosphorus, water holding capacity, water respiration, plant cover, arbuscular mycorrhizal fungi (AMF), and arachnid (spider) richness were significantly greater in greenspaces, while available ammonium was significantly greater in natural areas. The remaining attributes did not vary between land uses (Fig. 3). We did however find some effects of site size, with declining glucose, lignin, water and xyline respiration, and available phosphorus with increasing size of vegetation patch, irrespective of land use (natural cf. greenspace). Spider (arachnid) richness increased with increasing size, but only in natural areas (Supplementary Fig. 2).

a Locations of the 56 paired natural and urban greenspaces included in this study and b mean (±SE) ecosystem multiservices for mean (unweighted) and weighted ecosystem management services (EMS) and the eight ecosystem services for greenspaces and natural systems. Asterisks indicate significant differences in ecosystem services between greenspace and natural areas at P < 0.05.

Mean ecosystem multiservices (±SE) for greenspaces and natural systems at >25% threshold, >50% threshold, and >75% threshold. The number of functions at >75% threshold is greater (P < 0.05) in natural than greenspaces.

Mean (±SE) values of individual attributes in greenspaces and natural systems. Asterisks indicate a significant difference between greenspace and natural systems at P < 0.05. Units: Potential infiltration (ml water s−1), plant cover (%), water holding capacity (%), available P, available nitrate, available ammonium (mg kg−1 soil), AM fungal biomass (nmol g−1 soil), glucose, water and lignin respiration (μg CO2-C g−1 soil h−1), beta glucosidase, NAG, xylanase, phosphatase nmol activity g−1soil hr−1, NDVI (unitless), pest control (% x-1), and invertebrate and biocrust richness (number of ASVs).
Environmental factors associated with multiple ecosystem services
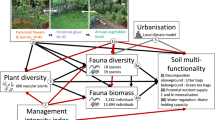
We found similar effects of different factors on weighted multiservices in both greenspaces and natural systems, and most of these effects were direct (Fig. 4a, b). There was also a strong locational effect for both greenspaces and natural systems, though the mechanisms were different. For example, in greenspaces, increasing latitude (distance from the equator) was associated with lower ecosystem multiservices, whereas in natural systems there were no strong effects (Fig. 4a−c). Total nitrogen and MAT were the most important factors associated with multiservices in natural areas, and sites with coarser soils were associated with lower multiservices, but only in natural sites (Fig. 4a−c). Reductions in MAT were associated with greater levels of multiservices in both systems. In greenspaces, greater levels of multiservices were associated with sites that were mown, but there were no effects of fertilisation or irrigation. The major factors associated with ecosystem multiservices and individual services remained relatively consistent when we considered unweighted (mean) multiservices (Supplementary Fig. 3), but there were some differences. For example, total N was positively associated with mean multiservices in both greenspaces and natural ecosystems.

Direct and indirect relationships between spatial (latitude, longitude), climate (MAP, MAT), size, age, management factors (irrigation, fertilisation, mowing), soils organic matter [OM], soil total nitrogen [TN] and total phosphorus [P], soil pH, soil texture [sand content] and the weighted ecosystem multiservices (EMS) for a greenspaces and b natural areas. Red, blue and grey vectors represent negative, positive and mixed relationships, respectively, and values on the vectors represent the path coefficients. c Standardised total effects (STE), i.e., the sum of direct and indirect effects of each of the attributes on ecosystem multiservices. Note that the model for natural areas does not contain information on management factors (mowing, fertilising, irrigating). The variance explained by the model (R2) of each parameter is given. Non-significant relationships are not shown. Model fit: Greenspace: χ2 = 0.28, df = 1, P = 0.594, NFI = 0.99; Natural area: χ2 = 0.423, df = 1, P = 0.516, NFI = 0.99.
Finally, we found mostly positive correlations among the 23 attributes and the environmental factors for both natural areas and greenspaces (Fig. 5). For example, the activity of most enzymes was positively correlated with soil total nitrogen and total phosphorus, and total phosphorus was positively associated with soil respiration and measures of available nitrogen. Further detailed analyses, however, suggest that there were also some clear differences between natural areas and greenspaces. For example, sandy soils were associated with lower available nitrate and ammonium, but only in natural systems. For greenspaces, we also found some management effects, with mown and fertilised sites associated with more plant cover and available phosphorus, as well as greater nematode richness.

Heatmap of Spearman’s rho correlations among individual ecosystem functions and environmental variables for greenspaces and natural areas. Only significance (P < 0.05) relationships are shown.
Discussion
Our current understanding of urban greenspaces is that their biotic and abiotic signatures differ from that of natural systems, largely because greenspace soils are highly altered and intensively managed26. However, such a claim is not well supported by the prevailing literature, likely due to the large range in environmental conditions experienced globally across different cities and their associated greenspaces26,30,31. The overwhelming result of our global study was that urban greenspaces not only delivered multiple ecosystem services, but the levels were of a similar magnitude to those found in nearby natural ecosystems. Our analyses suggest that the capacity of both systems to deliver multiservices is controlled primarily by climate, and to a lesser extent, soil total nitrogen and organic matter. Our results also show that management of urban greenspaces (mowing) can have positive effects on how they function and provide multiple ecosystem services. In general, urban and natural greenspaces were notably similar in areas with low levels of management. Our global assessment therefore adds to a growing recognition that greenspaces are far from being biologically or ecologically depauperate systems. Rather, they are crucial for supporting the flow of multiple services in urban systems.
Our global survey provides empirical evidence that, in general, urban greenspaces can deliver important services such as soil stability, water regulation, nutrient availability, plant and soil symbiosis, organic matter decomposition, net primary productivity, pest control, and soil biodiversity, which have been largely undervalued in previous urban studies when compared with natural ecosystems32. These services are not only critical for the maintenance and long-term sustainability of greenspaces for recreation and social well-being4,5, but also the maintenance of a diverse community of organisms that inhabit urban soils19. Even so, we also found that urban greenspaces may still have capacity to improve their levels of functioning when compared with natural ecosystems. For example, our analyses revealed that while urban greenspaces have a similar capacity to support multiple services (multiservices) at medium/low levels of functioning than natural ecosystems, natural ecosystems still have a significantly larger capacity to support multiple services that function at higher levels than urban greenspaces. Disturbance associated with urban greenspace management may constrain such high levels of functioning, those these disturbances were not identified in our study. These could include site-specific differences in the intensity of human use, or forms of irrigation or mowing that we could not account for in our study. Similarly, although we found generally similar results for greenspaces and natural areas when analysing both individual services and multiservices using the multi-threshold approach, both approaches detected small, but apparent, differences between greenspaces and natural systems. For example, nutrient availability and plant and soil symbiosis were significantly greater in greenspaces but pest control was greater in natural areas. Greenspaces are typically fertilised and irrigated, often with recycled water or effluent33,34, which would be expected to increase the availability of nutrients, particularly nitrogen. Together, we found that urban greenspaces are more similar to natural ecosystems than previously reported, and identified those services which could be improved when compared with natural ecosystems.
We also found similarities in the factors associated with greenspaces and natural systems. For example, total nitrogen was highly correlated with decomposition in both systems. Nitrogen availability drives decomposition, largely via its effects on soil microbial activity35, and to a lesser extent, arthropod abundance, through predation on bacteria and fungi36. Soil pH is an important statistical predictor of belowground biodiversity during pedogenesis37. However, we found that pH was poorly associated with most functions and services in greenspaces, potentially because pH conditions are controlled by management. In natural systems, however, soil pH was positively associated with lignin and glucose respiration (Fig. 4). The locational effect was also a shared predictor between urban and natural environments. These results suggest that similar environmental constraints control ecosystem services in both environments, explaining their overall similar level of functioning. Further, regions of the globe with finer soil textures supported lower levels of function. We also found declines in some services with increasing site size, irrespective of greenspace or natural area. Interestingly, we identified only one age-related effect, an increase in spider richness with increasing age of natural areas, despite the fact that overall spider richness was greater in greenspaces. Increasing spider richness with site age is likely due to greater habitat structure and complexity at older, natural sites38.
This study advances our understanding of the multiple ecosystem services supported by urban greenspaces. Despite their importance, these areas remain largely unstudied compared with natural ecosystems, particularly at the global scale. Only when multiple ecosystem services are considered at a high level of functioning do natural areas surpass greenspaces in the support of multiple services. Taken together, our results demonstrate that urban greenspaces are more similar to natural ecosystems than previously reported, and have a previously underestimated capacity to support multiple ecosystem services contributing to the stability and functioning of terrestrial ecosystems.
Methods
Study sites
We conducted a global standardised field survey of 56 paired urban greenspaces and natural ecosystems across 17 countries and six continents (Fig. 1a). Our survey targeted well-established urban parks and large residential gardens (greenspaces), and systems that represented the most common ecosystem type in each location in the absence of urbanisation (natural or semi-natural). Natural ecosystems were, on average, ~25 km distant from urban greenspaces. In each location, we established a 30 m × 30 m plot comprising three parallel transects of 30 m, spaced 15 m apart. Natural ecosystems were subject to variable but lower levels of management than urban environments (Supplementary Table 1). For example, while all greenspaces were sprayed for weeds, spraying was only carried out in three of the natural areas. Similarly, small amounts of debris were removed from eight of the 56 natural areas to assist with managing wildfires. Urban environments were exposed to a range of management practices such as mowing (n = 34 sites), fertilising (18 sites) and watering (30 sites; Supplementary Table 1).
Soil sampling
Across the sites, we collected three composite soil samples (0–5 cm) from five soil cores to account for locational heterogeneity. All samples were collected from the dominant site-specific microsite, i.e., beneath vascular plants, open areas (between plant canopies, either bare soil or supporting non-vascular plants). Each composite soil sample was divided into two subsamples. One was immediately frozen (−20 °C) for molecular analyses and the second was air-dried for physicochemical analyses. We analysed a total of 336 composite soil samples, based on three composite samples from each of the 56 pairs of plots in urban greenspaces and natural ecosystems. In all analyses, we report data based on the three plot-level composite samples.
Ecosystem and management attributes
Information on the age of sites was obtained from local management authorities such as reginal councils and government officials. Some of this information, particularly for large popular greenspaces and natural areas, was obtained from publicly available reports. Other information was sourced by contacting the relevant park management authorities. We defined park age as the time since major refurbishment (e.g., re-turfing, planting of new gardens) rather than establishment. For example, the Real Jardin Botanico in Madrid, Spain was constructed in 1755 but was assigned an age of 42 years because of extensive and significant refurbishment and soil removal in the late 1970s, which would have influenced our measures of soil ecosystem services. It was difficult to assign age to many of the natural areas as these could potentially have been unchanged for millennia. In the absence of other information on age, we assigned a nominal and conservative age of 3000 years to natural sites in order not to overinflate the explanatory power of some of our models. The size of sites was determined by digitising the area of each park using Google Maps. This was relatively straight forward for discrete greenspaces but more nuanced for natural areas, particularly extensive natural areas that were part of extensive natural systems (e.g., in Minas Gerais State, Brazil). In these cases, we used the boundary of the specific state or national park, but recognise that this is part of a larger area of natural area. Although we did not examine the potential effects of primary use (e.g., sports field cf. road verge) due to low number of observations, we acknowledge that these differences could affect different soil ecosystem services across our global dataset.
The extent of different actions such as mowing, irrigation, and fertilisation was gathered from the relevant local management authorities described above. Although these management practices were coded as binary data (0,1), we acknowledge that this is somewhat simplistic. Sites mown or irrigated more than six times per year were regarded as mown/irrigated. Data on fertiliser usage were obtained from park workers, and sites coded as ‘fertilised’ had to have received two or more applications per year. Not all urban sites would have received fertiliser at consistent rates or frequencies over time, but there were insufficient data to characterise temporal use of fertilisers or irrigation. We realise that the assignment of binary values is somewhat arbitrary, and because it does not account for historic use, it is a potential weakness of our study. However, our binary assignment in relation to moving, irrigation and fertilisation reflect an average practice across different greenspaces.
Ecosystem services
We measured 23 ecosystem attributes, combinations of which can be combined to provide surrogates or proxies of eight soil ecosystem services (soil stability, water regulation, nutrient availability, plant and soil mutualist relationships, organic matter decomposition, net primary productivity, pest control, and soil biodiversity).
Plant cover was used as a surrogate of soil stability because it is strongly and negatively related to soil erosion39. Plant cover was measured using the line-intercept method using data from the three 30 m transects. The water regulation service was assessed using the laboratory-based infiltrability technique of ref. 40, as well as water holding capacity (WHC), which measures the capacity of the soil to retain moisture and support plant growth. The water regulation service is critical for regulating important processes such as flooding, which causes substantial damage in cities worldwide15,41,42. Nutrient availability was assessed using measures of available phosphorus, nitrate, and ammonium, which were extracted from soils with ion exchange membranes (IEMs) (i.e., root simulators) in 1:15 soil:water solutions over 24 h. Resin-adsorbed N and P was extracted with 0.7 M NaCl for 1 h and determined using colorimetric analyses43. Nutrient availability assesses nutrients that are directly available to plants and microbes, and differs from measurements of total nutrients, which are more likely to be controlled by fertilisation practices. The plant-soil mutualism service was assessed using the biomass of arbuscular mycorrhizal fungi (AMF). The biomass of AMF was measured using microbial phospholipid fatty acids (PLFAs) (16:1w5c) according to ref. 44 using an Agilent 6890 gas chromatograph (Agilent Technologies, Palo Alto, CA, USA). This service is a measure of the capacity of terrestrial ecosystems to support key symbiotic process associated with the provision of nutrients from soil organisms to support plant growth. Our measure of the organic matter decomposition service was assessed by measuring soil (water-induced) respiration, glucose-induced respiration, lignin-induced respiration, and the activity of four enzymes associated with sugar (β-glucosidase), chiton (N-acetyl glucosaminidase; NAG), hemicellulose (xylanase) and phosphorus (phosphatase) degradation. These extracellular soil enzyme activities were measured with fluorometry on 1 g of soil45. The decomposition service is critical for decomposing organic matter in soils and making it available for plants through mineralisation processes. For Plant productivity we used NDVI values as a proxy for plant productivity because it is a measure of photosynthetic activity and large-scale vegetation distribution46. The NDVI data were average data from three remote sensing satellites: Sentinel-2, Landsat 8 and MODIS Terra MOD13Q1. Plant productivity is related to factors such as habitat for invertebrates, but also suppression of raindrops which can lead to water erosion.
The difference between the inverse of the relative abundance of soil-borne pathogenic nematodes (abundance x – 1) was used as a proxy of pathogen control or pathogenic nematode control. Many nematodes are known to be important plant pathogens that cause extensive damage to plant roots through herbivory. Nematodes were determined using the sequencing approach described below. The proportion of nematode sequences associated with pathogenic lifestyles was determined using the NEMAGUILD pipeline (https://rdrr.io/github/brendanf/FUNGuildR/man/get_funguild_db.html). Finally, the richness of tardigrades and biocrusts (estimated in the field quadrats) was used as a measure of soil faunal/biocrust diversity. A diverse community of soil invertebrates is important for decomposing litter and making it available to plants. Biocrust taxa also provide surface protection against erosion. This service was determined by calculating the richness of soil invertebrates (annelids, nematodes, arachnids, collembola, rotifers and tardigrades) using the Illumina MiSeq platform (Illumina Inc., CA, USA), and soil biocrusts (generally bryophytes, but sometimes lichens). DNA from 336 composite soil samples was extracted using the DNeasy PowerSoil Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. Invertebrate richness was also measured via amplicon sequencing. Bioinformatic processing was performed using the DADA2 pipeline47. Phylotypes [i.e., amplicon sequence variants (ASVs)] were identified at the 100% identity level. The ASV abundance tables were rarefied at 250 (18S rRNA gene for invertebrates) sequences per sample. Biocrust richness (mosses and lichens) determined in the field using data from the three 30 m transects. The cover of different biocrust types was assessed along the 30 m transects using the line-intercept method. Specimens (mosses and liverworts) were photographed and collected for identification by bryophyte experts48.
Statistical modelling
Multiservices
To address our research questions, we used a range of multiple ecosystem service (multiservice) indices: (i) average, (ii) weighted and (iii) multi-threshold multiservices. These indices provide complementary insights into how simultaneous ecosystem services might differ between greenspaces and natural systems. We determined the average multiservice as the average value of variables after standardisation (z-transformed, 0–1). To determine the weighted multiservice, we first calculated the standardised (0–1) average of the eight individual services then averaged these values. These two indices provide basic information on the average levels of multiple ecosystem services. The multiple-threshold method calculates the number of functions (services) that simultaneously exceed multiple critical thresholds49. To do this, the maximum observed value of each measured function is calculated and the number of functions that exceed a pre-established threshold are enumerated. In our study, we used three thresholds (25, 50 and 75%) that cover the whole spectrum. These thresholds provide information on the functional level in which more functions are maximised under natural and urban conditions.
Statistical approaches
We used linear models, with the ‘lmer’ function in the R statistical package50, to test for potential differences in individual attributes between greenspaces and natural environments. In these analyses a blocked design was included to account for the paired nature of greenspaces and natural environments at each of the 56 locations. Our models included information on the size and age of each greenspace and natural environment at each of the 56 locations and tested the effects of landuse (natural cf. greenspace), age, size and interactions between age and size. In the modelling we used sites as a random effect and allowed slopes to vary.
Structural equation modelling51 was used to explore potential direct and indirect effects of aridity, and soil attributes (soil organic matter, sand content, pH, total nitrogen, total phosphorus) on the weighted multiservice values for both natural areas and greenspaces separately, while accounting for the effects of geographical location (latitude, sine longitude, cosine longitude). Aridity was identified as precipitation/potential evapotranspiration and was derived from Consortium for Spatial Information (CGIAR-CSI) for the 1950–2000 period52 (https://cgiarcsi.community/2019/01/24/global-aridity-index-and- potential-evapotranspiration-climate-database-v2/). Mean annual temperature (MAT) and mean annual precipitation (MAP) data were obtained from the Worldclim database v253. We assessed soil pH in a 1:2.5 mass: volume soil and water suspension and total nitrogen (N) with an Elemental Analyser (C/N Flash EA 112 Series-Leco Truspec). Total soil P was determined using an ICP-OES spectrometer (ICAP 6500 DUO; Thermo-Scientific, Waltham, MA, USA) following digestion with nitric-perchloric acid and an ICP-OES spectrometer (ICAP 6500 DUO; Thermo-Scientific, Waltham, MA, USA). Additionally, we included management effects in the analyses for greenspaces, i.e., whether they were irrigated, fertilised and/or mown (coded as 0 or 1). Structural equation modelling allowed us to test hypothesised relationships among our various predictors and the weighted multiservices index based on an a priori model of expected effects given prior knowledge. Our models for natural environments demonstrated an adequate goodness of fit, as measured using χ2 (χ2 = 0.423; df = 1, P = 0.516, Root Mean Error of Approximation (RMSEA = 0.00; P = 0.91) and Bollen-Stine Bootstrap (P = 0.94). For greenspaces, these were χ2 = 0.28; df = 1, P = 0.594, RMSEA = 0.00; P = 0.87, Bollen-Stine Bootstrap P = 0.93). We also calculated the standardised total effects of each explanatory variable to demonstrate its total effect. Analyses were performed using AMOS 22 (IBM, Chicago, IL, USA) software. Fertilisation, mowing and irrigation were considered only in urban environments as natural ecosystems were not subjected to these management actions (Supplementary Table 1). Finally, we used Spearman’s correlations to investigate the relative influence of different environmental (aridity, organic matter, total N, soil pH, texture, sand content), management practice (mowing, fertilising, irrigating), size and age attributes on unweighted values of the 23 individual attributes.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
All the materials, raw data, and protocols used in the article are available upon request and without restriction, and all data will be made publicly available in a public repository (Figshare https://doi.org/10.6084/m9.figshare.24873135.v1) upon publication.
References
Cafaro, P., Hansson, P. & Götmark, F. Overpopulation is a major cause of biodiversity loss and smaller human populations are necessary to preserve what is left. Biol. Conserv. 272, 109646 (2022).
United Nations, Department of Economic and Social Affairs, Population Division (2019). World Urbanization Prospects: The 2018 Revision (ST/ESA/SER.A/420). New York: United Nations.
Enssle, F. & Kabisch, N. Urban green spaces for the social interaction, health and well-being of older people—An integrated view of urban ecosystem services and socio-environmental justice. Environ. Sci. Policy. 109, 36–44 (2020).
Kardan, O., Gozdyra, P. & Misic, B. et al. Neighborhood greenspace and health in a large urban center. Sci. Rep. 5, 11610 (2015).
Lee, A. C. K. & Maheswaran, R. The health benefits of urban green spaces: a review of the evidence. J. Public. Health. 33, 212–222 (2011).
Huang, C., Yang, J., Lu, H., Huang, H. & Yu, L. Green spaces as an indicator of urban health: evaluating its changes in 28 mega-cities. Remote. Sens. 9, 1266 (2017).
Jabbar, M., Yusoff, M. M. & Shafie, A. Assessing the role of urban green spaces for human well-being: a systematic review. GeoJ. 87, 4405–4423 (2022).
Kabisch, N. & Haase, D. Green justice or just green? Provision of urban green spaces in Berlin, Germany. Lands. Urban Plan. 122, 129–139 (2014).
Woodruff, S. C. & BenDor, T. K. Ecosystem services in urban planning: comparative paradigms and guidelines for high quality plans. Lands. Urban Plan. 152, 90–100 (2016).
Dorning, M. A., Koch, J., Shoemaker, D. A. & Meentemeyer, R. K. Simulating urbanization scenarios reveals tradeoffs between conservation planning strategies. Landsc. Urban Plan. 136, 28–39 (2015).
Liu, W., Chen, W. & Peng, C. Assessing the effectiveness of green infrastructures on urban flooding reduction: a community scale study. Ecol. Model. 291, 6–14 (2014).
Paul, M. J. & Meyer, J. L. Streams in the urban landscape. Annu. Rev. Ecol. Evol. Syst. 32, 333–365 (2001).
Setälä, H., Francini, G., Allen, J. A., Jumpponen, A., Hui, N. & Kotze, D. J. Urban parks provide ecosystem services by retaining metals and nutrients in soils. Environ. Pollution. 23, 451–461 (2017).
Haase, D., Frantzeskaki, N. & Elmqvist, T. Ecosystem services in urban landscapes: practical applications and governance implications. Ambio. 43, 407–412 (2014).
Chapman, E. J., Small, G. E. & Shrestha, P. Investigating potential hydrological ecosystem services in urban gardens through soil amendment experiments and hydrologic models. Urban Ecosyst. 25, 867–878 (2022).
Sandström, U. G., Angelstam, P. & Mikusiński, G. Ecological diversity of birds in relation to the structure of urban green space. Landsc. Urban Plan. 77, 39–53 (2006).
Sandifer, P. A., Sutton-Grier, A. E. & Ward, B. P. Exploring connections among nature, biodiversity, ecosystem services, and human health and well-being: opportunities to enhance health and biodiversity conservation. Ecosyst. Serv. 12, 1–15 (2015).
Strohbach, M. W., Arnold, E. & Haase, D. The carbon footprint of urban green space—A life cycle approach. Landsc. Urban Plan. 104, 220–229 (2012).
Ramirez, K. S., Leff, J. W., Barberán, A. & Bates, S. T. et al. Biogeographic patterns in below-ground diversity in New York City’s Central Park are similar to those observed globally. Proc. R. Soc. B. 28, 12014198820141988 (2014).
Li, H. & Liu, Y. Neighborhood socioeconomic disadvantage and urban public green spaces availability: a localized modeling approach to inform land use policy. Land. Use Policy. 57, 470–478 (2016).
Joassart-Marcelli, P. Leveling the playing field? urban disparities in funding for local parks and recreation in the Los Angeles Region. Environ. Plan A. 42, 1174–1192 (2010).
de la Barrera, F., Reyes-Paecke, S. & Banzhaf, E. Indicators for green spaces in contrasting urban settings. Ecol. Indic. 62, 212–219 (2016).
Jo, H. K., Kim, J. K. & Park, H. M. Carbon reduction and planning strategies for urban parks in Seoul. Urban For. Urban Gree. 41, 48–54 (2019).
Xu, L., You, H., Li, D. & Yu, K. Urban green spaces, their spatial pattern, and ecosystem service value: the case of Beijing. Habitat. Int. 56, 84–95 (2016).
Dobbs, C., Kendal, D. & Nitschke, C. R. Multiple ecosystem services and disservices of the urban forest establishing their connections with landscape structure and sociodemographics. Ecol. Indic. 43, 44–55 (2014).
Pavao‐Zuckerman, M. A. The nature of urban soils and their role in ecological restoration in cities. Restor. Ecol. 16, 642–649 (2008).
Bertoncini, A. P., Machon, N., Pavoine, S. & Muratet, A. Local gardening practices shape urban lawn floristic communities. Landsc. Urban Plann. 105, 53–61 (2012).
Hitchmough, J. Exotic plants and plantings in the sustainable, designed urban landscape. Landsc. Urban Plan. 100, 380–382 (2011).
Cheung, P. K., Nice, K. A. & Livesley, S. J. Irrigating urban green space for cooling benefits: the mechanisms and management considerations. Environ. Res. 1, 015001 (2022).
Klaus, V. H. & Kiehl, K. A conceptual framework for urban ecological restoration and rehabilitation. Basic. Appl Ecol. 52, 82–94 (2021).
Polovina, S., Radić, B., Ristić, R., Kovačević, J., Milčanović, V. & Živanović, N. Soil erosion assessment and prediction in urban landscapes: a new G2 model approach. Science 11, 4154 (2021).
Lahoti, S., Lahoti, A., Joshi, R. K. & Saito, O. Vegetation structure, species composition, and carbon sink potential of urban green spaces in Nagpur City, India. Land 9, 107 (2020).
Zhang, Z., Paschalis, A. & Mijic, A. Planning London’s green spaces in an integrated water management approach to enhance future resilience in urban stormwater control. J. Hydrol. 597, 126126 (2021).
Ferrini, F. & Baietto, M. Response to fertilization of different tree species in the urban environment. Arboric. Urban For 32, 93–99 (2006).
Treseder, K. K. Nitrogen additions and microbial biomass: a meta-analysis of ecosystem studies. Ecol. Lett. 11, 1111–1120 (2008).
Delgado-Baquerizo, M., Eldridge, D. J., Ochoa, V., Gozalo, B., Singh, B. K. & Maestre, F. T. Soil microbial communities drive the resistance of ecosystem multifunctionality to global change in drylands across the globe. Ecol. Lett. 20, 1295–1305 (2017).
Delgado-Baquerizo, M., Bardgett, R. D. & Vitousek, P. M. et al. Changes in belowground biodiversity during ecosystem development. Proc. Nat. Acad. Sci. USA. 116, 6891–6896 (2019).
Platen, R., Berger, G. & Malt, S. The impact of structural and landscape features of set-asides on the spiders (Araneae) of the herb layer. J. Arachnol. 41, 143–150 (2013).
Durán, Z. V. H. & Rodríguez, P. C. R. Soil-erosion and runoff prevention by plant covers. A review. Agron. Sustain. Dev. 28, 65–86 (2008).
Mills, A. J. & Fey, M. V. A simple laboratory infiltration method for measuring the tendency of soils to crust. Soil. Use. Manage. 20, 8–12 (2004).
Paprotny, D., Sebastian, A. & Morales-Nápoles, O. et al. Trends in flood losses in Europe over the past 150 years. Nat. Commun. 9, 1985 (2018).
Chen, A. S., Hammond, M. J., Djordjević, S., Butler, D., Khan, D. M. & Veerbeek, W. From hazard to impact: flood damage assessment tools for mega cities. Nature 82, 857–890 (2016).
Maestre, F. T., Quero, J. L., Gotelli, N. J., Escudero, A. & Ochoa, V. et al. Plant species richness and ecosystem multifunctionality in global drylands. Science 335, 214–218 (2012).
Frostegard, A. et al. Use and misuse of PLFA measurements in soils. Soil. Biol. Biochem. 43, 1621–1625 (2011).
Bell, C. W., Fricks, B. E., Rocca, J. D., Steinweg, J. M., McMahon, S. K., & Wallenstein, M. D. High-throughput fluorometric measurement of potential soil extracellular enzyme activities. J. Vis. Exp. Nov 15, e50961 (2013).
Pettorelli, N., Vik, J. O., Mysterud, A., Gaillard, J. M., Tucker, C. J. & Stenseth, N. C. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends. Ecol. Evol. 20, 503–510 (2005).
Callahan, B. J. et al. DADA2: high-resolution sample inference from Illumina amplicon data. Nature 7, 581–583 (2016).
Eldridge, D. J., Guirado, E. & Reich, P. B. et al. The global contribution of soil mosses to ecosystem services. Nat. Geosci. 16, 430–438 (2023).
Byrnes, J. E. K. et al. Investigating the relationship between biodiversity and ecosystem multifunctionality: challenges and solutions. Methods Ecol. Evol. 5, 111–124 (2014).
Bates, D., Mächler, M., Bolker, B. & Walker, S. Fitting linear mixed-effects models using lme4. Stat. Comput. 1406, 133–199 (2014).
Grace, J. B. Structural Equation Modelling and Natural Systems. (Cambridge University Press, UK, 2006).
Zomer, R. J., Trabucco, A., Bossio, D. A. & Verchot, L. V. Climate change mitigation: a spatial analysis of global land suitability for clean development mechanism afforestation and reforestation. Agr. Ecosyst. Environ. 126, 67–80 (2008).
Fick, S. E. & Hijmans, R. J. WorldClim 2: new 1‐km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315 (2017).
Acknowledgements
We thank the researchers involved in the MUSGONET project for collection of field data and soil samples. This study was supported by a 2019 Leonardo Grant for Researchers and Cultural Creators, BBVA Foundation (URBANFUN), and by the BES grant agreement No LRB17\1019 (MUSGONET). M.D-B. acknowledges support from the Spanish Ministry of Science and Innovation for the I + D + i project PID2020-115813RA-I00 funded by MCIN/AEI/10.13039/501100011033. M.D-B. is also supported by a project of the Fondo Europeo de Desarrollo Regional (FEDER) and the Consejería de Transformación Económica, Industria, Conocimiento y Universidades of the Junta de Andalucía (FEDER Andalucía 2014−2020 Objetivo temático “01 - Refuerzo de la investigación, el desarrollo tecnológico y la innovación”) associated with the research project P20_00879 (ANDABIOMA). D.J.E. is supported by the Hermon Slade Foundation (HSF21040). H.C. is supported by the National Natural Science Foundation of China (32101335), and The Young Science and Technology Talent Support Project of Jilin Province (QT202226), J.D. is supported by Young Elite Scientists Sponsorship Program by BAST (No. BYESS2023456) and the Fundamental Research Funds for the Central Universities, and M.B. by a Ramón y Cajal grant (RYC2021-031797-I) from Spanish Ministry of Science and Innovation. F.A. acknowledges support from FONDECYT 1220358 and C.P. support from the EU’s H2020 research and innovation programme under grant agreement No 101000224. A.R. acknowledges support from the FCT (SFRH/BDP/108913/2015), the MCTES, FSE, UE, and the CFE (UIDB/04004/2021) research unit financed by FCT/MCTES through national funds (PIDDAC). S.A. thanks the ANID/FONDECYT 1170995 and the ANID ACT 192027. T.P.M. acknowledges funding from the National Research Foundation of South Africa (UID 118981). T.G. and T.U.N. were supported by the research projects J4-3098 and J4-4547, and by the Research Program in Forest Biology, Ecology, and Technology (P4-0107) of the Slovenian Research Agency. J.P.V. thanks the SERB (EEQ/2021/001083, SIR/2022/000626), DST (DST/INT/SL/P-31/2021) and Banaras Hindu University, IoE (6031) incentives grant for providing support for Phytomicrobiome and soil microbiome research. L.W. and J.W. were supported by the Program for Introducing Talents to Universities (B16011), and the Ministry of Education Innovation Team Development Plan (2013-373).
Author information
Authors and Affiliations
Contributions
M.D.-B. and D.J.E. developed the original ideas presented in the manuscript. D.J.E. and H.C. wrote the first draft, which was edited by J. Ding with contributions from all authors. H.C., J.D. and D.J.E. undertook the statistical analyses. Field data were collected by M.D-B., D.J.E., M.B., J.L.B-P., C.P., S.A., F.A., A.R.B., J.D., T.G., J.G.I., Y-R.L., T.P.M., T.U.N., G.F.P-B., A. Rey, A.R., C.S., A.L.T., P.T., L.W., J.W., E.Z., X.Z. and X-Q.Z. Laboratory analyses were performed by M.D-B., J-Z.H., T.S-S., W.S., H.C. and P.T and bioinformatic analyses by M.D-B.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Eldridge, D.J., Cui, H., Ding, J. et al. Urban greenspaces and nearby natural areas support similar levels of soil ecosystem services. npj Urban Sustain 4, 15 (2024). https://doi.org/10.1038/s42949-024-00154-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s42949-024-00154-z
