
Abstract
Although mammalian target of rapamycin 1 (mTORC1) negatively regulates autophagy in cultured cells, how autophagy impacts mTORC1 signalling, in particular in an in vivo setting, is less clear. Here we show that autophagy supports mTORC1 hyperactivation in neural stem cells (NSCs) lacking tuberous sclerosis complex subunit 1 (Tsc1), thereby promoting defects in NSC maintenance, differentiation and tumourigenesis, and the formation of the neurodevelopmental lesion of tuberous sclerosis complex (TSC). Analysing mice that lack Tsc1 and the essential autophagy gene Rb1-inducible coiled-coil 1 (Rb1cc1, also called Fip200) in NSCs, we find that TSC-deficient cells require autophagy to maintain mTORC1 hyperactivation under energy-stress conditions, likely to provide free fatty acids via lipophagy to serve as an alternative energy source for OXPHOS. In vivo, inhibition of lipophagy or its downstream catabolic pathway reverses defective phenotypes caused by Tsc1-null NSCs and reduces tumourigenesis in mouse models. These results reveal a cooperative function of selective autophagy in coupling energy availability with TSC pathogenesis and suggest a potential new therapeutic strategy to treat people with TSC.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others

References
Saxton, R. A. & Sabatini, D. M. mTOR signaling in growth, metabolism, and disease. Cell 169, 361–371 (2017).
Hay, N. & Sonenberg, N. Upstream and downstream of mTOR. Genes Dev. 18, 1926–1945 (2004).
Ben-Sahra, I. & Manning, B. D. mTORC1 signaling and the metabolic control of cell growth. Curr. Opin. Cell Biol. 45, 72–82 (2017).
Costa-Mattioli, M. & Monteggia, L. M. mTOR complexes in neurodevelopmental and neuropsychiatric disorders. Nat. Neurosci. 16, 1537–1543 (2013).
Crino, P. B. mTOR: a pathogenic signaling pathway in developmental brain malformations. Trends Mol. Med. 17, 734–742 (2011).
Chan, J. A. et al. Pathogenesis of tuberous sclerosis subependymal giant cell astrocytomas: biallelic inactivation of TSC1 or TSC2 leads to mTOR activation. J. Neuropathol. Exp. Neurol. 63, 1236–1242 (2004).
Magri, L. et al. Sustained activation of mTOR pathway in embryonic neural stem cells leads to development of tuberous sclerosis complex-associated lesions. Cell Stem Cell 9, 447–462 (2011).
Way, S. W. et al. Loss of Tsc2 in radial glia models the brain pathology of tuberous sclerosis complex in the mouse. Hum. Mol. Genet. 18, 1252–1265 (2009).
Zhou, J. et al. Tsc1 mutant neural stem/progenitor cells exhibit migration deficits and give rise to subependymal lesions in the lateral ventricle. Genes Dev. 25, 1595–1600 (2011).
Zordan, P. et al. Tuberous sclerosis complex-associated CNS abnormalities depend on hyperactivation of mTORC1 and Akt. J. Clin. Invest. 128, 1688–1706 (2018).
Knobloch, M. et al. Metabolic control of adult neural stem cell activity by Fasn-dependent lipogenesis. Nature 493, 226–230 (2013).
Rafalski, V. A. & Brunet, A. Energy metabolism in adult neural stem cell fate. Prog. Neurobiol. 93, 182–203 (2011).
Shyh-Chang, N., Daley, G. Q. & Cantley, L. C. Stem cell metabolism in tissue development and aging. Development 140, 2535–2547 (2013).
Stoll, E. A. et al. Neural stem cells in the adult subventricular zone oxidize fatty acids to produce energy and support neurogenic activity. Stem Cells 33, 2306–2319 (2015).
Xie, Z., Jones, A., Deeney, J. T., Hur, S. K. & Bankaitis, V. A. Inborn errors of long-chain fatty acid beta-oxidation link neural stem cell self-renewal to autism. Cell Rep. 14, 991–999 (2016).
Duvel, K. et al. Activation of a metabolic gene regulatory network downstream of mTOR complex 1. Mol. Cell 39, 171–183 (2010).
Hu, C. J., Wang, L. Y., Chodosh, L. A., Keith, B. & Simon, M. C. Differential roles of hypoxia-inducible factor 1alpha (HIF-1alpha) and HIF-2alpha in hypoxic gene regulation. Mol. Cell Biol. 23, 9361–9374 (2003).
Porstmann, T. et al. SREBP activity is regulated by mTORC1 and contributes to Akt-dependent cell growth. Cell Metab. 8, 224–236 (2008).
Laplante, M. & Sabatini, D. M. An emerging role of mTOR in lipid biosynthesis. Curr. Biol. 19, R1046–R1052 (2009).
Valvezan, A. J. et al. mTORC1 couples nucleotide synthesis to nucleotide demand resulting in a targetable metabolic vulnerability. Cancer Cell 32, 624–638 (2017). e625.
Levine, B. & Kroemer, G. Autophagy in the pathogenesis of disease. Cell 132, 27–42 (2008).
Mizushima, N., Levine, B., Cuervo, A. M. & Klionsky, D. J. Autophagy fights disease through cellular self-digestion. Nature 451, 1069–1075 (2008).
Mizushima, N. & Komatsu, M. Autophagy: renovation of cells and tissues. Cell 147, 728–741 (2011).
Ng, S., Wu, Y. T., Chen, B., Zhou, J. & Shen, H. M. Impaired autophagy due to constitutive mTOR activation sensitizes TSC2-null cells to cell death under stress. Autophagy 7, 1173–1186 (2011).
Chen, Y., Wei, H., Liu, F. & Guan, J. L. Hyperactivation of mammalian target of rapamycin complex 1 (mTORC1) promotes breast cancer progression through enhancing glucose starvation-induced autophagy and Akt signaling. J. Biol. Chem. 289, 1164–1173 (2014).
Di Nardo, A. et al. Neuronal Tsc1/2 complex controls autophagy through AMPK-dependent regulation of ULK1. Hum Mol. Genet. 23, 3865–3874 (2014).
Parkhitko, A. et al. Tumorigenesis in tuberous sclerosis complex is autophagy and p62/sequestosome 1 (SQSTM1)-dependent. Proc. Natl Acad. Sci. USA 108, 12455–12460 (2011).
Lamark, T. & Johansen, T. Aggrephagy: selective disposal of protein aggregates by macroautophagy. Int. J. Cell Biol. 2012, 736905 (2012).
Kim, I., Rodriguez-Enriquez, S. & Lemasters, J. J. Selective degradation of mitochondria by mitophagy. Arch. Biochem. Biophys. 462, 245–253 (2007).
Kubli, D. A. & Gustafsson, A. B. Mitochondria and mitophagy: the yin and yang of cell death control. Circ. Res. 111, 1208–1221 (2012).
Palikaras, K. & Tavernarakis, N. Mitophagy in neurodegeneration and aging. Front. Genet. 3, 297 (2012).
Singh, R. et al. Autophagy regulates lipid metabolism. Nature 458, 1131–1135 (2009).
Liu, K. & Czaja, M. J. Regulation of lipid stores and metabolism by lipophagy. Cell Death Differ. 20, 3–11 (2013).
Martinez-Lopez, N. et al. Autophagy in the CNS and periphery coordinate lipophagy and lipolysis in the brown adipose tissue and liver. Cell Metab. 23, 113–127 (2016).
Hara, T. et al. Suppression of basal autophagy in neural cells causes neurodegenerative disease in mice. Nature 441, 885–889 (2006).
Komatsu, M. et al. Loss of autophagy in the central nervous system causes neurodegeneration in mice. Nature 441, 880–884 (2006).
Wang, C. et al. Elevated p62/SQSTM1 determines the fate of autophagy-deficient neural stem cells by increasing superoxide. J. Cell Biol. 212, 545–560 (2016).
Wang, C., Liang, C. C., Bian, Z. C., Zhu, Y. & Guan, J. L. FIP200 is required for maintenance and differentiation of postnatal neural stem cells. Nat. Neurosci. 16, 532–542 (2013).
Kim, J., Kundu, M., Viollet, B. & Guan, K. L. AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat. Cell Biol. 13, 132–141 (2011).
Kwiatkowski, D. J. et al. A mouse model of TSC1 reveals sex-dependent lethality from liver hemangiomas, and up-regulation of p70S6 kinase activity in Tsc1 null cells. Hum. Mol. Genet. 11, 525–534 (2002).
Sun, S. et al. Constitutive activation of mTORC1 in endothelial cells leads to the development and progression of lymphangiosarcoma through VEGF autocrine signaling. Cancer Cell 28, 758–772 (2015).
Wang, C., Yeo, S., Haas, M. A. & Guan, J. L. Autophagy gene FIP200 in neural progenitors non-cell autonomously controls differentiation by regulating microglia. J. Cell. Biol. 216, 2581–2596 (2017).
Sharma, M. et al. Subependymal giant cell astrocytoma: a clinicopathological study of 23 cases with special emphasis on proliferative markers and expression of p53 and retinoblastoma gene proteins. Pathology 36, 139–144 (2004).
Chen, S. et al. Distinct roles of autophagy-dependent and -independent functions of FIP200 revealed by generation and analysis of a mutant knock-in mouse model. Genes. Dev. 30, 856–869 (2016).
Liu, J. et al. Beclin1 controls the levels of p53 by regulating the deubiquitination activity of USP10 and USP13. Cell 147, 223–234 (2011).
Sancak, Y. et al. Ragulator-Rag complex targets mTORC1 to the lysosomal surface and is necessary for its activation by amino acids. Cell 141, 290–303 (2010).
Nigrovic, L. E., Kimia, A. A., Shah, S. S. & Neuman, M. I. Relationship between cerebrospinal fluid glucose and serum glucose. N. Engl. J. Med. 366, 576–578 (2012).
Kimmelman, A. C. & White, E. Autophagy and tumor metabolism. Cell Metab. 25, 1037–1043 (2017).
Galluzzi, L., Pietrocola, F., Levine, B. & Kroemer, G. Metabolic control of autophagy. Cell 159, 1263–1276 (2014).
Son, J. et al. Glutamine supports pancreatic cancer growth through a KRAS-regulated metabolic pathway. Nature 496, 101–105 (2013).
Chakrabarti, P., English, T., Shi, J., Smas, C. M. & Kandror, K. V. Mammalian target of rapamycin complex 1 suppresses lipolysis, stimulates lipogenesis, and promotes fat storage. Diabetes 59, 775–781 (2010).
Tuunanen, H. et al. Trimetazidine, a metabolic modulator, has cardiac and extracardiac benefits in idiopathic dilated cardiomyopathy. Circulation 118, 1250–1258 (2008).
Zacharowski, K., Blackburn, B. & Thiemermann, C. Ranolazine, a partial fatty acid oxidation inhibitor, reduces myocardial infarct size and cardiac troponin T release in the rat. Eur. J. Pharmacol. 418, 105–110 (2001).
Huang, S. C. et al. Cell-intrinsic lysosomal lipolysis is essential for alternative activation of macrophages. Nat. Immunol. 15, 846–855 (2014).
Mayer, N. et al. Development of small-molecule inhibitors targeting adipose triglyceride lipase. Nat. Chem. Biol. 9, 785–787 (2013).
Long, J. Z. et al. Selective blockade of 2-arachidonoylglycerol hydrolysis produces cannabinoid behavioral effects. Nat. Chem. Biol. 5, 37–44 (2009).
Ouimet, M. et al. Autophagy regulates cholesterol efflux from macrophage foam cells via lysosomal acid lipase. Cell Metab. 13, 655–667 (2011).
Dubland, J. A. & Francis, G. A. Lysosomal acid lipase: at the crossroads of normal and atherogenic cholesterol metabolism. Front. Cell Dev. Biol. 3, 3 (2015).
Zhang, Y. et al. Coordinated regulation of protein synthesis and degradation by mTORC1. Nature 513, 440–443 (2014).
Schmidt, E. V. The role of c-myc in cellular growth control. Oncogene 18, 2988–2996 (1999).
Morita, M. et al. mTORC1 controls mitochondrial activity and biogenesis through 4E-BP-dependent translational regulation. Cell Metab. 18, 698–711 (2013).
Wei, H. et al. Suppression of autophagy by FIP200 deletion inhibits mammary tumorigenesis. Genes Dev. 25, 1510–1527 (2011).
Yang, S. et al. Pancreatic cancers require autophagy for tumor growth. Genes Dev. 25, 717–729 (2011).
Guo, J. Y. et al. Activated Ras requires autophagy to maintain oxidative metabolism and tumorigenesis. Genes Dev. 25, 460–470 (2011).
Nie, D. et al. Tsc2-Rheb signaling regulates EphA-mediated axon guidance. Nat. Neurosci. 13, 163–172 (2010).
Djouadi, F., Bonnefont, J.-P., Munnich, A. & Bastin, J. Characterization of fatty acid oxidation in human muscle mitochondria and myoblasts. Mol. Genet. Metab. 78, 112–118 (2003).
Yeo, S. K., Wen, J., Chen, S. & Guan, J. L. Autophagy differentially regulates distinct breast cancer stem-like cells in murine models via EGFR/Stat3 and Tgfbeta/Smad signaling. Cancer Res. 76, 3397–3410 (2016).
Acknowledgements
We thank our colleagues Y. Zheng, Q. Lu, M. Czyzyk-Krzeska, D. Hui and T. Cunningham, and our laboratory members G. Doerman and B. Peace. Research was supported by National Institutes of Health (NIH) grants NS094144 and CA211066 to J.L.G. and NS103981 to C.W.
Author information
Authors and Affiliations
Contributions
C.W., J.-L.G.: project concept, research design, data analysis, manuscript writing. C.W.: experiments. C.W., M.A.H.: mouse model analyses, OCR assay. J.W.: immunoblots. P.S., D.R.P.: fatty acid β-oxidation assay. S.Y.: FACS analysis. F.Y., T.O.: generated Atg7 KO in WT and Tsc2 KO MEFs. S.C.: constructed Atg5 KO in Rheb-CA MEFs.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Analyses of autophagy, mTORC1 signalling and NSCs from SVZ tissues and neurospheres.
a, Brain size (upper panel) and H&E staining of sagittal sectioned brain (lower panel) of Ctrl, Tsc1GFAPcKO, 2cKO and Fip200GFAP cKO mice at P21. Five independent experiments gave similar results. b, H&E staining of sagittal sectioned brain at two positions of Ctrl, Tsc1GFAPcKO, 2cKO and Fip200GFAP cKO mice at P0. Mean ± s.e. of the number of SVZ cells in Ctrl, Tsc1GFAPcKO, 2cKO and Fip200GFAP cKO mice at P0 shown on the right. n = 5 independent experiments. c,d, The levels of p62, Fip200 and Tsc1 (c) and LC3 (d) in isolated SVZ tissue of Ctrl, Tsc1GFAPcKO, 2cKO and Fip200GFAPcKO mice treated with or without CQ for 14 d. Three independent experiments gave similar results. e, Means ± s.e. of the autophagy flux (calculated as LC3-II with CQ divided by LC3-II without CQ as shown in b). n = 3 independent experiments. f,g, Immunofluorescence of p62 (f), p4EBP1 (phosphorylated at T37/46) (g) and DAPI in SVZ of Tsc1GFAPcKO and 2cKO mice at P21. Bottom panels in g show details of p4EBP1+ cells in boxed area. h, Mean ± SE of the percentage of p4EBP1+cells in SVZ of Ctrl, Tsc1GFAP cKO, 2cKO, and FIP200GFAP cKO mice at P21. n = 4 independent experiments. i,j, Mean ± SE of the number of GFAP+Sox2+NSC (i) and the number of GFAP+Nestin+BrdU+ cells of total BrdU+ cells (j) in SVZ of Ctrl, Tsc1GFAP cKO, 2cKO, and FIP200GFAP cKO mice at P21. n = 6 independent experiments. k–m, Mean ± SE of the number (k) and size (l,m) of primary (k,l) and secondary (m) neurospheres from SVZ cells of Ctrl, Tsc1GFAP cKO, 2cKO, and FIP200GFAP cKO mice at P21. n = 4 (k), 3 (l,m) independent experiments. n, Lysates from primary neurospheres of Ctrl, Tsc1GFAPcKO, 2cKO, and FIP200GFAPcKO mice examined by immunoblotting with indicated antibodies. Three independent experiments gave similar results. o, The level of LC3 in neurospheres of Ctrl, Tsc1GFAPcKO, 2cKO, and FIP200GFAPcKO mice treated with or without BafA1. Three independent experiments gave similar results. The dotted lines indicated the boundaries of SVZ with LV (f,g). E: ependymal layer; LV: lateral ventricle; ST: striatum; SVZ: subventricular zone. Bar = 100 mm. Data were analyzed by one-way ANOVA with Tukey’s post-hoc test (b,e,h–m).
Extended Data Fig. 2 Fip200 KO and Fip200-4A knock-in mutant rescued defective phenotypes in Tsc1GFAPcKO SVZ.
a,b, Immunofluorescence of DCX (a), GFAP (b) and DAPI in SVZ of Tsc1GFAPcKO and 2cKO mice at P21. Four independent experiments gave similar results. c–f, Mean ± s.e. of the percentage of β-III tubulin+ cells (c), NeuN+ cells (d), GFAP+ cells (e) and MBP+ cells (f) from differentiated neurospheres of Ctrl, Tsc1GFAPcKO, 2cKO and Fip200GFAPcKO mice. n = 4 (c,d), 5 (e,f) independent experiments. g, Frequency of SEN-like structure in Ctrl, Tsc1GFAPcKO, 2cKO, 2cKI, and Fip200GFAPcKO mice at P7, P14 and P28. The animal numbers used at different postnatal stages are indicated. (h–l) Immunofluorescence of pS6RP (h), nestin and Ki67 (i), GFAP and Sox2 (j), NG2 (k), DCX and GFAP (l) in SEN-like lesion in Tsc1GFAPcKO brain at P21. Three independent experiments gave similar results. m, Immunofluorescence of pS6RP and DAPI in SVZ of Tsc1GFAPcKO and 2cKI mice at P21. Five independent experiments gave similar results. n, Mean ± s.e. of the number of pS6RP+cells in SVZ of Ctrl, Tsc1GFAPcKO, 2cKI and Fip200GFAP cKI mice at P21. n = 5 independent experiments. o, H&E staining of SVZ from Ctrl, 2cKI and Fip200GFAP cKI mice at P28. Five independent experiments gave similar results. p, Mean ± s.e. of SVZ cell number of Ctrl, Tsc1GFAPcKO, 2cKI and Fip200GFAPcKI mice at P21. n = 5 independent experiments. q,r, Mean ± s.e. of the number of GFAP+Nestin+ NSCs (q) and GFAP+Sox2+ NSC (r) in SVZ of Ctrl, Tsc1GFAPcKO, 2cKI and Fip200GFAPcKI mice at P21. n = 5 independent experiments. The dotted lines indicated the boundaries of SVZ or SEN-like lesion with LV. E, ependymal layer; LV, lateral ventricle; SEN, subependymal nodule-like lesion; ST, striatum; SVZ, subventricular zone. Scale bar, 100 μm (a,b,m,o), 40 μm (h–l). Data were analysed by one-way ANOVA with Tukey’s post-hoc test (c–f,n,p–r) or Chi-square test (g).
Extended Data Fig. 3 Autophagy sustained mTORC1 hyperactivation in Tsc-deficient MEFs under energy stresses.
a, Lysates from WT and Tsc2 KO MEFs treated with DMSO or BafA1 examined by immunoblots with antibodies for LC3 and actin. Three independent experiments gave similar results. b, Mean ± s.e. of the number of LC3 puncta of DMSO or Spautin1 treated WT and Tsc2 KO MEFs in normal medium and glucose-free medium (without FBS) with or without BafA1 for 2 hours. Spautin1 was preincubated for 12 h. n = 6–12 independent experiments for at least 200 cells. c, Lysates from DMSO- or Spautin1-treated WT MEFs and Tsc2 KO MEFs under normal, glucose-free and amino-acid-free conditions (without FBS) for 2 h were examined with indicated antibodies. Spautin1 was preincubated for 12 h. Three independent experiments gave similar results. d, Immunofluorescence of mTOR, LAMP2 and DAPI in WT MEF, Tsc2 KO MEF and Tsc2/Atg7 2KO MEF under normal medium (without FBS), glucose-free medium (without FBS) or glucose-free medium with 10 μM Spautin1 (without FBS) for 2 h. Spautin1 was preincubated for 12 h. Four independent experiments gave similar results. e, Mean ± s.e. of the Pearson correlation coefficient (PCC) of mTOR co-localization on LAMP2+ structure in WT MEF, Atg7 KO MEF, Tsc2 KO MEF and 2KO MEF under normal medium, glucose-free medium and glucose-free medium (without FBS) with Spautin1 for 2 h. n = 5 independent experiments for at least 200 cells. f, Lysates from WT MEFs and Tsc2 KO MEFs treated with DMSO or Spautin1 were examined by immunoblots with antibody for LC3 and vinculin. BafA1 was used to block autophagosome degradation, and Spautin1 was preincubated for 12 h before experiments. Three independent experiments gave similar results. Mean ± s.e. of the relative level of LC3II of DMSO- or Spautin1-treated WT MEFs and Tsc2 KO MEFs with or without BafA1 for 2 h on the right. n = 3 independent experiments. g, Lysates from WT MEFs, Atg7 KO MEFs (2 independent clones), Tsc2 KO MEFs and 2KO MEFs (2 independent clones) were examined by immunoblots with antibodies as indicated. Three independent experiments gave similar results. h, Lysates were extracted from WT MEF, Atg7 KO MEF (clone 1), Tsc2 KO MEF and 2KO MEF (clone 1) in normal medium, glucose-free medium and 2DG-supplemented medium (without FBS) for 2 h. The levels of phosphorylated S6K, total S6K, phosphorylated S6RP, total S6RP and vinculin were examined. Three independent experiments gave similar results. Scale bar, 10 μm. Data were analysed by two-tailed Student’s t test (b,f) or Pearson’s correlation coefficient (e).
Extended Data Fig. 4 Analysis of the role of autophagy, TORC1 hyper-activation, ATP content and AMPK targeting phosphorylation in Tsc-deficient neurospheres, MEF, 293 cells and SVZ.
a, Lysates extracted from WT and Tsc2 KO MEF in normal, glucose-free and 2DG-supplemented media (without FBS) for 2 h were examined by immunoblot as indicated. Cells were preincubated with DMSO or 10 μM Spautin1 for 12 h. Three independent experiments gave similar results. b, Lysates were extracted from WT, Atg7 KO, Tsc2 KO and 2KO MEF in normal, glucose-free and 2DG-supplemented media (without FBS) for 2 h. The levels of phosphorylated Raptor, total Raptor, phosphorylated ACC and total ACC were examined. Three independent experiments gave similar results. c, Mean ± s.e. of the ATP content of Tsc2 KO MEFs in normal, glucose-free and 2DG supplemented media (without FBS) supplemented with 10 μM Spautin1 or 20 μM CQ for 2 h was shown. The autophagy inhibitors were preincubated for 12 h. n = 3–9 independent experiments. d, Mean ± s.e. of the ATP content of Tsc2 KO MEFs and 2KO MEFs in normal, glucose-free and 2DG-supplemented media (without FBS) for 2 h. n = 7–8 independent experiments. e, Lysates from Ctrl, Fip200 KD, Tsc1 KD, and 2KD 293 cells were examined by immunoblots with indicated antibodies. Four independent experiments gave similar results. f,g, Means ± s.e.. of the protein levels of Tsc1 (f) and Fip200 (g) in Ctrl, Fip200 KD, Tsc1 KD, and 2KD 293 cells were shown. n = 4 independent experiments. h, The level of LC3-II was examined in Ctrl, Fip200 KD, Tsc1 KD, and 2KD 293 cells treated with or without BafA1. Three independent experiments gave similar results. i, Mean ± s.e. of the level of LC3-II in Ctrl, Fip200 KD, Tsc1 KD and 2KD 293 cells treated with or without BafA1was shown. n = 3 independent experiments. j, Lysates were prepared from Ctrl, Fip200 KD, Tsc1 KD and 2KD 293 cells treated with 25 mM 2DG (without FBS) for 2 h and analysed by immunoblot with antibodies as indicated. Three independent experiments gave similar results. k,l, Mean± s.e. of ATP-related and maximum OCR in WT and Tsc2 KO MEFs under normal or glucose-free conditions (k) and normal or 2DG treatment conditions (l). n = 9–11 (k) and 4 (l) independent experiments. m,n, Mean ± s.e. of ATP-related and maximum OCR in WT MEFs, Atg7 KO MEFs, Tsc2 KO MEFs and 2KO MEFs under glucose-free conditions (m) and 2DG-treatment conditions (n). n = 8 independent experiments. Data were analysed using two-tailed Student’s t test (c,d,i,k–n), one-way ANOVA with Tukey’s post hoc test (f,g).
Extended Data Fig. 5 Energy stressed Tsc-deficient cells activated lipolysis through autophagy.
a, Lysates were extracted from DMSO- or Spautin1-treated WT and Tsc2 KO MEFs supplemented with 20 mM glutamine under normal, glucose-free (upper panels), or 2DG-treatment conditions (lower panels) for 2 h. The levels of phosphorylated S6K and total S6K were examined by immunoblot with antibodies as indicated. Three independent experiments gave similar results. b,c, Mean ± s.e. of the ammonia content (b) and the glycogen content (c) in WT and Tsc2 KO MEFs under normal and glucose-free media (for c, with or without BafA1 treatment) for 2 h. n = 6 (b) and 2 (c) independent experiments. d, Mean ± s.e. of the content of FFAs in Ctrl, Tsc1 KD, 2KD and Fip200 KD 293 cells under normal and glucose-free media. n = 6 independent experiments. e, Immunofluorescence for lipid droplets, LC3 and DAPI in DMSO-, BafA1- and Spautin1-treated Tsc2 KO MEFs under normal or glucose-free conditions (without FBS) for 2 h. Four to nine independent experiments for at least 200 cells gave similar results. f, Immunofluorescence for LDs, LAMP2 and DAPI in DMSO or Spautin1-treated WT and Tsc2 KO MEFs under normal or glucose-free conditions (without FBS) for 2 h. Six to ten independent experiments for at least 200 cells gave similar results. g, Mean ± s.e. of ATP content in CQ or Spautin1-treated Tsc2 KO MEFs under normal conditions and supplemented with BSA or BSA–palmitate for 2 h. n = 3–6 independent experiments. h–l, Representative images of LDs (indicated by Oil Red O staining) in SVZ of Ctrl (H and K), Tsc1GFAPcKO (i,l), and 2cKO (j) mice treated with (k,l) or without (h–j) ETO at P21. Five independent experiments gave similar results. m, Mean ± s.e. of the percentage of cells with lipid droplets in SVZ of Ctrl, Tsc1GFAPcKO and 2cKO mice treated with or without ETO at P21. n = 5 independent experiments. Scale bars, 10 μm.Data were analysed by two-tailed Student’s t test (b,d,g) or one-way ANOVA with Tukey’s post-hoc test (c,m).
Extended Data Fig. 6 Regulation of activation of mTORC1, AMPK and ATP content by inhibitors for β-oxidation and AMPK in energy-stressed Tsc-deficient cells.
a–d, Lysates were extracted from WT MEFs and Tsc2 KO MEFs in normal medium, glucose-free medium and 2DG-supplemented medium (without FBS) for 2 h. Cells were preincubated with β-oxidation inhibitors of Rano (a,b) and TMZ (c,d) for 12 hours and followed by 2 hours treatment. In a and c, the levels of phosphorylated Raptor, total Raptor, phosphorylated ACC, total ACC and vinculin were examined. In b and d, the levels of phosphorylated S6K, total S6K, phosphorylated S6RP, total S6RP and vinculin were examined. Three independent experiments gave similar results. e, Immunofluorescence of mTOR, LAMP2 and DAPI in ETO and Rano treated Tsc2 KO MEFs under normal and glucose-free medium for 2 h. Five independent experiments for at least 200 cells gave similar results. f, Mean ± s.e. of the Pearson correlation coefficient of mTOR localization on LAMP2+ structure in Tsc2 KO MEF treated with ETO or Rano under glucose-free conditions. n = 5 independent experiments. g, Lysates from vehicle-, ETO- or Compound C (CC)-treated Tsc2 KO MEFs in normal, glucose-free and 2DG supplemented media (without FBS) for 2 h were examined by immunoblot using antibodies as indicated. Cells were preincubated with ETO with or without CC for 24 h. Three independent experiments gave similar results. h, Mean ± s.e. of the ATP content of β-oxidation inhibitors of ETO-, TMZ- and Rano-treated WT MEFs in normal, glucose-free and 2DG media (without FBS) for 2 h. The β-oxidation inhibitors were preincubated for 12 h. n = 3–6 independent experiments. i, Mean ± s.e. of ATP content in ETO-treated Ctrl and Tsc1 KD 293 cells under glucose deprivation conditions for 2 h. n = 5 independent experiments. Scale bar,10 μm. Data were analysed by two-tailed Student’s t test (h,i) or Pearson’s correlation coefficient (f).
Extended Data Fig. 7 Lipophagy released FFAs in energy-stressed mTORC1 hyperactivated Tsc-deficient cells.
a,b, Mean ± s.e. of glycerol release from (a) and the number of LDs in (b) WT MEFs under normal and glucose-free media (without FBS) for 2 h supplemented with DMSO, orlistat, atglistat or JZL were shown. n = 3 (a) and 5 (b) independent experiments. c, Immunofluorescence of LAMP2, mTOR and DAPI in Orlistat-treated WT and Tsc2 KO MEFs under glucose-free medium (without FBS) for 2 h. Insets in details on right panels. Three independent experiments gave similar results. d, Lysates from WT and Tsc2 KO MEFs were examined by immunoblot of LAL and actin. Three independent experiments gave similar results. e, Lysates from scrambled shRNA and two individual LAL shRNAs infected WT MEF were examined by immunoblot of LAL and actin. Three independent experiments gave similar results. f,g, Mean ± s.e. of the glycerol release (f) and FFA content (g) from scrambled-shRNA- and LAL-shRNA-#1-nfected WT and Tsc2 KO MEF in normal and glucose-free media for 2 h were shown. n = 3 (f) and 6 (g) independent experiments. h,i, Mean ± s.e. of the number of LDs (h) and the content of TG (i) in scrambled-shRNA- and LAL-shRNA-#1-infected Tsc2 KO MEFs under normal medium and glucose-deprivation conditions. n = 5 independent experiments. j, Mean ± s.e. of ATP-related OCR of scrambled-shRNA- and LAL-shRNA-#1-infected WT and Tsc2 KO MEFs under glucose deprivation conditions. n = 4 independent experiments. k, Mean ± s.e. of the ATP content of scrambled-shRNA- and LAL-shRNA-#1-infected Tsc2 KO MEFs in normal, glucose-free and 2DG media with or without supplement of BSA–palmitate for 2 h were shown. n = 3 independent experiments. l, Lysates from scrambled-shRNA- and LAL-shRNA-#1-infected WT and Tsc2 KO MEFs in normal, glucose-free and 2DG-supplemented media for 2 hours. The levels of phosphorylated S6K, total S6K, phosphorylated S6RP, total S6RP and vinculin were examined by Western blot as indicated. Three independent experiments gave similar results. m–o, Immunofluorescence of pS6RP (m), pACC (n), pAMPK (o) and DAPI in SVZ of Tsc1GFAPcKO mice treated with vehicle, 2DG, CQ and CQ + 2DG. Five independent experiments gave similar results. p, Immunofluorescence of pAKT and DAPI in SVZ of Tsc1GFAPcKO mice treated with vehicle, 2DG, CQ and CQ + 2DG. Five independent experiments gave similar results. q, Mean ± s.e. of the percentage of pAKT+ cells in SVZ of Tsc1GFAPcKO mice treated with vehicle, 2DG, CQ and CQ + 2DG. n = 5 independent experiments. r, Immunofluorescence of pERK and DAPI in SVZ of Tsc1GFAPcKO mice treated with vehicle, 2DG, CQ and CQ + 2DG. Five independent experiments gave similar results. s, Mean ± s.e. of the percentage of pERK+ cells in SVZ of Tsc1GFAPcKO mice treated with vehicle, 2DG, CQ and CQ + 2DG. n = 5 independent experiments. Dotted lines indicated the boundaries between SVZ and LV. E, ependymal layer; LV, lateral ventricle; ST, striatum; SVZ, subventricular zone. Scale bar, 10 μm (c); 100 μm (m–p,r). Data were analysed by two-tailed Student’s t test (a,b,f–k), one-way ANOVA with Tukey’s post-hoc test (q,s).
Extended Data Fig. 8 AMPK activation stimulated autophagy and lipophagy was required in Tsc-deficient cells. Postnatal developmental defects of cerebral cortex in Tsc1GFAPcKO mice were rescued in 2cKO mice.
a, Mean ± s.e. of the number of GFAP+Sox2+ NSCs in SVZof Ctrl mice treated with CQ+2DG and Tsc1GFAPcKO mice treated with vehicle, 2DG, CQ and CQ + 2DG. n = 6 independent experiments. b, Mean ± s.e. of the number of GFAP+Nestin+BrdU+ cell of total BrdU+ cells (b) in SVZ of Ctrl mice treated with CQ+2DG and Tsc1GFAPcKO mice treated with vehicle, 2DG, CQ and CQ + 2DG. n = 6 independent experiments. c, Mean ± s.e. of the number of GFAP+Sox2+ NSCs in SVZ of Tsc1GFAPcKO mice treated with vehicle, 2DG, ETO, Rano, TMZ, ETO + 2DG, Rano + 2DG and TMZ + 2DG are shown. n = 5 independent experiments. d, Mean ± s.e. of the number of GFAP+Nestin+BrdU+ cells of total BrdU+ cells in SVZ of Ctrl and Tsc1GFAP cKO mice treated with vehicle, 2DG, ETO, Rano, TMZ, ETO + 2DG, Rano + 2DG and TMZ + 2DG. n = 5 independent experiments. e,f, Mean ± s.e. of the number of DCX+ cells (e) and NeuN+ cells (f) in SVZ of Ctrl and Tsc1GFAPcKO mice treated with vehicle, 2DG, ETO, Rano, TMZ, ETO+2DG, Rano+2DG and TMZ+2DG. n = 5 independent experiments. g, Lysates from WT and Tsc2 KO MEFs in normal or glucose-free media were examined by immunoblots with antibodies as indicated. Three independent experiments gave similar results. h, Lysates from WT MEFs, AMPKα1 KO MEFs (2 independent clones), Tsc2 KO MEFs and 2KO MEFs (2 independent clones) were examined by immunoblots with antibodies, as indicated. Three independent experiments gave similar results. i, Lysates were extracted from WT MEFs, AMPKα1 KO MEFs, Tsc2 KO MEFs and 2KO MEFs in glucose-free media with or without BafA1 for 2 h. The levels of LC3-II and vinculin were examined. Three independent experiments gave similar results. j, H&E staining of sagittal sectioned brain indicating hydrocephaly of Tsc1GFAPcKO lateral ventricle, but not in Ctrl, 2cKO, 2cKI and Fip200GFAP cKO mice at P21. Arrows indicated SEN-like structures in Tsc1GFAPcKO brain. Three independent experiments gave similar results. k, Mean ± s.e. of brain weight of Ctrl, Tsc1GFAPcKO, 2cKO and Fip200GFAP cKO mice at P7, P14 and P21. n = 3 independent experiments. l, H&E staining of sagittal sectioned brain indicating thicker cortex of Tsc1GFAPcKO brain, but not in Ctrl, 2cKO, 2cKI and Fip200GFAP cKO mice at P21. The sagittal sections for middle of the brain (middle position) and 1 mm lateral to the middle position were stained. The dashed lines indicated the boundaries of cortex and CC. Four independent experiments gave similar results. m–o, Mean ± s.e. of cortex thickness of Ctrl, Tsc1GFAPcKO, 2cKO and Fip200GFAPcKO mice at two different section positions (m,n) at P7, P14, and P21, and P21 2cKI (o) mice at middle position. n = 4 (m,n) and 5 (o) independent experiments. p–r, Mean ± s.e. of cortex cell size (p) and cortex cell number (q) of Ctrl, Tsc1GFAPcKO, 2cKO and Fip200GFAP cKO mice by H&E staining at P21, and cortex TUNEL-positive cell number (r) at P7 and P21. n = 4 (p, q) and 3 (r) independent experiments. CC, corpus callosum; hippo, hippocampus; I-VI indicates layers of cerebral cortex. Scale bar, 50 μm. Data were analysed by one-way ANOVA with Tukey’s post-hoc test (a–f,o–r) or one-way ANOVA (k,m,n,r).
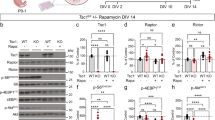
Extended Data Fig. 9 Postnatal developmental defects of cerebral cortex in Tsc1GFAPcKO mice were rescued in 2cKO mice and by rapamycin treatment.
a, Immunofluorescence of pS6RP, NeuN and DAPI in cortex of Ctrl, Tsc1GFAPcKO, 2cKO and Fip200GFAP cKO mice at P21. Inset shows details of pS6RP and NeuN in boxed area. Four independent experiments gave similar results. b,c, Mean ± s.e. of the percentage of pS6RP+NeuN+ of NeuN+ cells (b) and the cell size of NeuN+ cells (c) in the cortex of Ctrl, Tsc1GFAP cKO, 2cKO, and Fip200GFAP cKO mice at P21. n = 4 independent experiments, at least 300 cells counted. d, Immunofluorescence of GFAP and DAPI in SVZ of Ctrl, Tsc1GFAPcKO, 2cKO and Fip200GFAPcKO mice at P21. Four independent experiments gave similar results. e, Mean ± s.e. of the relative percentage of GFAP+ cells in the cortex of Ctrl, Tsc1GFAPcKO and 2cKO mice at P21. n = 4 independent experiments, at least 300 cells counted. f, Immunofluorescence of pS6RP, NeuN and DAPI in the cortex of rapamycin treated Ctrl and Tsc1GFAP mice at P21. Inset shows details of pS6RP and NeuN in boxed area. The dashed lines indicated the boundaries of cortex and CC. Four independent experiments gave similar results. g, Mean ± s.e. of the percentage of pS6RP+NeuN+ of NeuN+ cells in the cortex of rapamycin treated Ctrl andTsc1GFAPcKO mice at P21. n = 4 independent experiments, at least 300 cells counted. h, Mean ± s.e. of the brain weight of rapamycin treated Ctrl andTsc1GFAPcKO mice at P21. n = 3 independent experiments. i, H&E staining of sagittal sectioned brain indicating cortex of Ctrl, Tsc1GFAPcKO, 2cKO, 2cKI and Fip200GFAPcKO mice at P21. The sagittal sections for middle of the brain (middle position) and 1 mm lateral to the middle position were stained. The dashed lines indicated the boundaries of cortex and CC. Four independent experiments gave similar results. j, Mean ± s.e. of cortex thickness of Ctrl, Tsc1GFAPcKO, 2cKO and Fip200GFAP cKO mice at two different section positions at P21. n = 3 (middle) or 4 (–1 mm) independent experiments. CC, corpus callosum; hippo, hippocampus; I-VI indicates layers of cerebral cortex. Scale bar, 50 μm. Data were analysed by one-way ANOVA with Tukey’s post-hoc test (b,c,e,s,h,j).
Supplementary information
Supplementary Information
Supplementary Table 1 and Discussion
Source data
Source Data Fig. 1
Original images for immunoblot presented in main and supplementary figures.
Source Data Fig. 2
Original images for immunoblot presented in main and supplementary figures.
Source Data Fig. 3
Original images for immunoblot presented in main and supplementary figures.
Source Data Fig. 4
Original images for immunoblot presented in main and supplementary figures.
Source Data Fig. 5
Original images for immunoblot presented in main and supplementary figures.
Rights and permissions
About this article

Cite this article
Wang, C., Haas, M.A., Yang, F. et al. Autophagic lipid metabolism sustains mTORC1 activity in TSC-deficient neural stem cells. Nat Metab 1, 1127–1140 (2019). https://doi.org/10.1038/s42255-019-0137-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s42255-019-0137-5
This article is cited by
-
Autophagy inhibition prevents lymphatic malformation progression to lymphangiosarcoma by decreasing osteopontin and Stat3 signaling
Nature Communications (2023)
-
The regulatory role of lipophagy in central nervous system diseases
Cell Death Discovery (2023)
-
Functional Analysis of a Novel Immortalized Murine Microglia Cell Line in 3D Spheroid Model
Neurochemical Research (2023)
-
Crosstalk between autophagy inhibitors and endosome-related secretory pathways: a challenge for autophagy-based treatment of solid cancers
Molecular Cancer (2021)
-
Amino acid levels determine metabolism and CYP450 function of hepatocytes and hepatoma cell lines
Nature Communications (2020)
