
Abstract
Flooding is a common and critical disaster in agriculture, because it causes defects in plant growth and even crop loss. An increase in herbivore populations is often observed after floods, which leads to additional damage to the plants. Although molecular mechanisms underlying the plant responses to flooding have been identified, how plant defence systems are affected by flooding remains poorly understood. Herein, we show that submergence deactivates wound-induced defence against herbivore attack in Arabidopsis thaliana. Submergence rapidly suppressed the wound-induced expression of jasmonic acid (JA) biosynthesis genes, resulting in reduced JA accumulation. While plants exposed to hypoxia in argon gas exhibited similar reduced wound responses, the inhibitory effects were initiated after short-term submergence without signs for lack of oxygen. Instead, expression of ethylene-responsive genes was increased after short-term submergence. Blocking ethylene signalling by ein2-1 mutation partially restored suppressed expression of several wound-responsive genes by submergence. In addition, submergence rapidly removed active markers of histone modifications at a gene locus involved in JA biosynthesis. Our findings suggest that submergence inactivates defence systems of plants, which would explain the proliferation of herbivores after flooding.
Similar content being viewed by others


Introduction
Flooding is a widespread agricultural disaster, which causes significant damage and loss to crops. Submergence is a representative event of floods in which water covers aerial plant tissues, leading to oxygen deprivation1,2. As oxygen acts as a terminal electron acceptor in the oxidative phosphorylation process during mitochondrial respiration, hypoxia causes energy deficit, which is the major reason for reduced growth and death in plants exposed to hypoxic conditions2,3. In addition to hypoxia-induced plant damage, outbreak of herbivores is another phenomenon observed after flooding. Both continuous and periodic flooding trigger an increase in herbivore populations in rice4. Elevated ovipositional and feeding activities of insect herbivores are also observed in flooded plants including citrus5,6. However, the effects of flooding on plant defence responses to herbivore attack remain largely unknown.
Plant defence against herbivores involves the sensing of mechanical wounding, which is a hallmark of herbivore attack. Early signals, including reactive oxygen species (ROS), Ca2+, and glutamate, are produced in the wounded area7,8. Glutamate triggers an increase in intracellular Ca2+ levels through GLUTAMATE RECEPTOR-LIKE proteins, which are cation-permeable ion channels8. The increase in cytosolic Ca2+ is propagated to nearby tissues, which activates jasmonic acid (JA) biosynthesis8,9. The expression of genes involved in JA biosynthesis, including those encoding LIPOXYGENASE (LOX), ALLENE OXIDE SYNTHASE (AOS), ALLENE OXIDE CYCLASE (AOC), and OXOPHYTODIENOATE-REDUCTASE (OPR), is induced within 1 h of wounding10,11,12. The accumulated JA-Ile directly binds to the F-box protein CORONATINE INSENSITIVE 1 (COI1), which forms a functional SCFCOI1-JAZ co-receptor complex to trigger the degradation of JASMONATE ZIM-DOMAIN (JAZ) proteins13. The activation of the downstream JA signalling by JAZ degradation induces defence responses, including the accumulation of defensive proteins and toxic secondary metabolites14,15.
Studies on the plant adaptive mechanisms in response to submergence have revealed that plants have developed two distinctive growth strategies to survive submergence. To escape from the water, plants focus their energy into upward growth of stems, which is a beneficial strategy under shallow and short-term submergence conditions16,17. However, under long-term submergence, plants tend to reduce growth to save energy and carbohydrate reserves for tolerance16. Ethylene plays pivotal roles in both strategies under submergence conditions. Submergence rapidly increases ethylene concentrations in plant cells, because water restricts diffusion of the ethylene gas2. Accumulated ethylene induces shoot elongation, adventitious root formation, soluble carbohydrate breakdown, and ethanolic fermentation18,19,20,21. ETHYLENE INSENSITIVE 2 (EIN2) is a key factor in ethylene signalling by which it protects proteasomal degradation of transcription factors to transmit ethylene signals22. EIN2-deficient mutants exhibit reduced expression of hypoxia-inducible genes and decreased survival after submergence23,24,25, showing its critical role on hypoxia tolerance.
Hypoxia affects a broad range of gene expressions by changing histone modifications to regulate transcriptional activity. In human cultured cells, hypoxia alters histone-3 lysine-4 trimethylation and H3K36me3, possibly by inhibiting Jumanji-C histone demethylase26. A study on the histone modifications in rice showed that submergence changes histone methylation and acetylation at ADH1 and PDC1 loci, which are responsive to submergence27. Notably, while H3K4 methylation is altered upon short-term submergence, H3 acetylation is increased after long-term submergence. These results indicate that plants recognize duration of submergence and modify transcriptional activity of key genes for survival under submergence conditions.
In this work, we found that submergence rapidly inactivates wound-induced JA biosynthesis in Arabidopsis. Plants that had undergone submergence exhibited reduced wound responses and increased susceptibility to herbivores. Rapid ethylene responses upon submergence were shown to be partially responsible for decreased wound responses. Submergence also triggered changes of histone methylation and acetylation at the OPR3 locus. Our results indicate that submergence inactivates plant wound responses, which possibly explains outbreaks of herbivores after flooding.
Results
Submergence inhibits wound-induced gene expressions
Although the early signals of the wound responses and downstream JA signalling mechanisms have been identified, wound-sensing mechanisms are not well understood. Plant leaves are surrounded by the epidermal cells; thus, internal aeration is tightly controlled by the stomatal apertures28. As wounding disrupts the enclosed structures in plant tissues, we hypothesized that an abrupt increase in aeration in plant tissues might be an initial cue for sensing mechanical wounding. To limit aeration in wounded tissues, we submerged the entire seedlings and wounded the cotyledons and leaves immediately in the water. We chose JAZs and OPR3 genes as representative wound-responsive JA-responsive genes and a JA biosynthesis gene, respectively29,30. The expression of JAZ and OPR3 genes was analysed to monitor plant wound responses. Whereas the expression levels of JAZ7, JAZ10, and OPR3 were significantly elevated after wounding in the air (JAZ7, P = 0.00005, Hedges′ g = 5.80; JAZ10, P = 0.000005, Hedges′ g = 7.63; OPR3, P < 1.0 × 10−7, Hedges′ g = 14.4), the induction was largely suppressed when wounding was done in water (Fig. 1a). We hypothesized that, if plants cannot sense wounds in the water, re-aeration would lead to recovery of the wound responses. However, expression of the wound-responsive genes was still suppressed following re-aeration (Fig. 1b). In addition, when seedlings were wounded in the air after submergence, wound responses were not recovered (Fig. 1c). Wound-induced gene expression was also significantly suppressed after submergence in adult plants grown on soil for 3 weeks (Supplementary Fig. 1; JAZ7, P = 0.0073, Hedges′ g = 2.68; JAZ10, P = 0.0067, Hedges′ g = 2.72; OPR3, P = 0.013, Hedges′ g = 2.41). We examined the promoter activity of JAZ10 gene using pJAZ10:GUS seedlings; the results showed that wound-induced GUS expression was largely down-regulated after submergence (Fig. 1d). Time-course expression analysis after wounding showed that expression of the JAZ7, JAZ10, and OPR3 genes was largely decreased particularly at 1 h after wounding in the seedlings that had undergone submergence (Fig. 1e). Increasing submergence duration of more than 1 h resulted in slight additional suppression in the OPR3 expression (Fig. 1f). Together, these data suggest that while abrupt aeration would not be the initial cue for wound sensing, experience of submergence negatively affects plant wound responses.

Ten-day-old seedlings were used. Error bars indicate ±SD. Letters indicate groups that are statistically significantly different from each other (P < 0.05, Tukey’s test). a Effects of submergence on wound responses. Cotyledons and leaves of the Col-0 seedlings were left in the air (air) or transferred into the water (sub), wounded immediately and then harvested after 1 h incubation. Three biological replicates were averaged. b Re-aeration does not recover wound responses. The Col-0 seedlings were wounded in the water immediately after submergence and incubated for 1 h. Submerged seedlings were then re-aerated for 1 h before they were harvested (Sub re-air) or were left in water for another hour before harvest (Sub). Three biological replicates were averaged. c Submergence inhibits wound responses of re-aerated seedlings. The Col-0 seedlings were submerged for 1 h without wounding and then transferred to air. Re-aerated seedlings were immediately wounded and then harvested after 1 h. Three biological replicates were averaged. d Promoter activity of the JAZ10 gene. Ten-day-old pJAZ10:GUS transgenic seedlings were submerged and wounded as described in c. Seedlings were fixed with acetone 1 h after the wounding. Intensities of GUS signals were measured using ImageJ software. Four biological replicates were averaged. Scale bars indicate 0.1 cm. e Time-course expression of genes after wounding. Seedlings were submerged for 1 h and then transferred to air. Re-aerated seedlings were wounded and then harvested at the indicated time points. Three biological replicates were averaged. f Effects of submergence time on wound responses. Seedlings were submerged for the indicated time points and then transferred to air. Re-aerated seedlings were wounded immediately and then harvested after 1 h. Three biological replicates were averaged.
Submergence suppresses JA biosynthesis to reduce defence against herbivores
The analysed wound-responsive genes were related to JA biosynthesis and JA signalling12,31. To determine whether the submergence affects JA biosynthesis or the JA signalling, we treated plants with methyl JA (MeJA) after submergence. The expression of JAZ7, JAZ10, and OPR3 genes was largely induced by MeJA treatment, as reported previously32 (Fig. 2a). Notably, the expression of JAZ7 was not altered and that of JAZ10 was reduced only slightly after submergence. In contrast, MeJA-induced OPR3 expression was largely suppressed after submergence, similar to the results observed upon wounding. These data suggest that submergence affects JA biosynthesis. We next analysed the expression of other genes involved in the JA biosynthesis pathways (Fig. 2b). The wound-induced expression of genes encoding AOCs, AOS, and LOXs, which play key roles in JA biosynthesis33, was significantly reduced after submergence except for LOX6 (Fig. 2c and Supplementary Fig. 2; AOC2, P < 1.0 × 10−7, Hedges′ g = 20.1; LOX2, P < 1.0 × 10−7, Hedges′ g = 15.4; AOC1, P = 0.000074, Hedges′ g = 5.31; AOC3, P = 0.00056, Hedges′ g = 4.00; AOC4, P = 0.0055, Hedges′ g = 4.25; AOS, P = 0.0000096, Hedges′ g = 7.00; LOX3, P = 0.0000053, Hedges′ g = 7.55; LOX4, P = 0.0026, Hedges′ g = 3.17). Consistently, JA accumulation upon wounding was also decreased after submergence (Fig. 2d), indicating that submergence affects the transcription of JA biosynthesis genes to inhibit wound-induced JA accumulation.

Ten-day-old seedlings were used. Error bars indicate ±SD. Letters indicate groups that are statistically significantly different from each other (P < 0.05, Tukey’s test). a Expression of JA-responsive genes after MeJA treatment. The Col-0 seedlings were submerged for 1 h and then transferred to the air. Re-aerated seedlings were treated with 25 μM MeJA. Whole seedlings were harvested 1 h after the treatment. Three biological replicates were averaged. b JA biosynthesis pathways. 13(S)-HPOT, (9Z,11E,15Z,13S)-13-hydroperoxy-9,11,15-octadecatrienoic acid; 12-oxo-PDA, 12-oxo-10,15(Z)-octadecatrienoic acid; OPC-8:0, 3-oxo-2(2′(Z)-pentenyl)-cyclopentane-1-octanoic acid. c Expression of JA biosynthesis genes after the submergence. The Col-0 seedlings were submerged and wounded as described in Fig. 1c. Three biological replicates were averaged. d JA accumulation after the wounding. The Col-0 seedlings were submerged and wounded as described in Fig. 1c. Five biological replicates were averaged. e Herbivore-feeding assay. Four-week-old Col-0 plants were submerged for 1 h and then transferred to the air. Two caterpillars (larval stage L1) of the Brassicaceae specialist P. rapae were released onto a single reaerated 4-week-old plant. Larval weights were measured before and 5 days after the caterpillar feeding (n = 8~24). The box plot shows median and quartiles. Scale bars indicate 0.5 cm.
It is well known that JA mediates plant defence against herbivore attacks34. As our data showed that submergence reduces wound-induced JA biosynthesis, we examined plant defence responses after submergence using Brassicaceae specialist, Pieris rapae caterpillars. Soil-grown plants were submerged and then transferred to the air before herbivore feeding. We put ~20 caterpillars on 10 plants, but only collected 8~9 caterpillars after 5 days, possibly because several of them moved away from the plants. Larvae fed on the plants that had undergone submergence showed a significantly higher weight than those fed on control plants (Fig. 2e; P < 1.0 × 10−7; Hedges′ g = 3.72), indicating that defence against herbivores is suppressed after submergence.
Hypoxia inhibits wound responses after re-aeration
Wounding triggers the accumulation of early signalling molecules in the wounded leaves; thereafter, the signals are propagated to other tissues, leading to JA accumulation7,8,9. Therefore, we analysed whether submergence affects early wound responses. Hydrogen peroxide (H2O2) is one of the initial signals of wounding7, but submergence did not alter H2O2 accumulation after wounding (Supplementary Fig. 3a). Increase in the concentration of cytosolic Ca2+ is another important event in the early wound responses9. We analysed the expression of TCH2 and TCH3, which are molecular markers of cytosolic Ca2+ increase35. The expression of these genes was induced upon wounding, but submergence without wounding also elevated the expression of TCH2 and TCH3 (Supplementary Fig. 3b). Wounding after submergence did not lead to additional increase in gene expressions. As the wound-induced cytosolic accumulation of Ca2+ is controlled by glutamate8, we measured glutamate levels using 35S:CHIB-iGluSnFR glutamate reporter plants, which have been previously described8. As with molecular Ca2+ markers, there was an accumulation of glutamate after submergence (Supplementary Fig. 3c). Again, wounding did not further elevate glutamate levels after submergence. If saturation of glutamate accumulation and the corresponding Ca2+ increase diminished wound responses in submerged plants, then high concentrations of glutamate treatment should have similar results. However, the wound-induced expression of JAZ10 and OPR3 genes was still observed in both the mock- and glutamate-treated plants (Supplementary Fig. 3d). Submergence-mediated decrease in the expression of wound-responsive genes was also observed after glutamate treatment. Together, these data indicate that suppression of wound responses after submergence is not directly related with early wound signals.
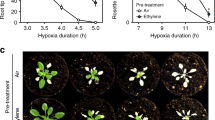
As the solubility of oxygen is lower in water than in air, hypoxia responses are induced in submerged plants36. To investigate whether the reduction of wound responses after submergence was related to oxygen deprivation, we built a system for incubation of plants under a continuous flow of argon gas (Fig. 3a). Incubation of the seedlings in a transparent box filled with argon gas for 2 h triggered induction of a gene encoding ALCOHOL DEHYDROGENASE 1 (ADH1), a molecular marker for hypoxia36 (Fig. 3b). After argon gas treatment, seedlings were transferred to ambient air and then wounded. The expression of JAZ10 and OPR3 genes was analysed 1 h after wounding. Expression of wound-responsive genes was significantly reduced after hypoxia (Fig. 3c; JAZ10, P = 0.0082, Hedges′ g = 2.62; OPR3, P = 0.0000048, Hedges′ g = 7.69), suggesting that oxygen availability is related with suppression of plant wound responses.

Ten-day-old seedlings were used. Error bars indicate ±SD. a Schematic illustration of the experiment. The Col-0 seedlings were incubated in the transparent glove box filled with Ar gas for 2 h. Seedlings were then transferred to the ambient air and wounded. b Expression of the ADH1 gene. Whole seedlings treated as described in a were harvested before and 1 h after the wounding. Expression of the ADH1 gene was analysed as a hypoxia marker. Three biological replicates were averaged. Letters indicate groups that are statistically significantly different from each other (P < 0.05, Tukey’s test). c Expression of the wound-responsive genes. Whole seedlings treated as described in a were harvested 1 h after the wounding. The JAZ10 and OPR3 genes were analysed. Letters indicate groups that are statistically significantly different from each other (P < 0.05, Tukey’s test).
Ethylene is involved in submergence-mediated suppression of wound responses
For detailed examination of the effects of submergence on plant wound responses, we submerged seedlings for different time durations before wounding. Submergence for even 1 min resulted in inhibition of wound responses and more than 10 min of submergence strongly suppressed the expression of wound-responsive genes (Fig. 4a). Less than 10 min of submergence is not sufficient to induce hypoxia in plants under light conditions37,38. In this work, plants were submerged in the light. In agreement with previous studies, expression of a hypoxia marker gene, ADH1, was not induced during short-term submergence (Fig. 4a). We thus speculated that hypoxia is not the main reason for suppression of wound responses by short-term submergence. To find clues, we analysed additional JA-responsive genes in the plants that had undergone submergence. Although expression of JR1 and VSP1 genes exhibited similar pattern to that of JAZ10, expression of PR genes was not largely altered by both wound and submergence in our work (Supplementary Fig. 4). Notably, expression of ethylene-responsive factors (ABR1 and ERF4) and ethylene- and JA-responsive PDF1.2 was significantly induced by 1 h submergence11,39,40 (Fig. 4b and Supplementary Fig. 4; ABR1, P = 0.0000022, Hedges′ g = 13.2; ERF4, P = 0.00039, Hedges′ g = 5.18; PDF1.2, P = 0.033, Hedges′ g = 3.67). Ethylene is a key phytohormone that mediates plant submergence responses2. To examine cross-talks between ethylene signals and wound responses, we investigated wound responses after submergence using ethylene-insensitive ein2-1 mutant. Wound-induced expression of AOC2, JAZ7, and VSP1 genes after submergence was partially restored in ein2-1 mutant, but other JA-biosynthesis and JA-responsive genes were not significantly altered by ein2-1 mutation (Fig. 4c). These data suggest that ethylene partially mediates early submergence responses to inhibit plant wound responses, but there are other unknown regulators in these processes.

Ten-day-old seedlings were used. Error bars indicate ±SD. a Effects of submergence time on wound responses. The Col-0 seedlings were submerged for the indicated time periods and then transferred to the air. Re-aerated seedlings were wounded immediately and harvested 1 h after the treatment. For ADH1, the Col-0 seedlings were harvested right after 1 h submergence. Three biological replicates were averaged. Letters indicate groups that are statistically significantly different from each other (P < 0.05, Tukey’s test). NS, not significant. b Expression of wound- and ethylene-responsive genes. The Col-0 seedlings were submerged and wounded as described in Fig. 1c. Three biological replicates were averaged. Letters indicate groups that are statistically significantly different from each other (P < 0.05, Tukey’s test). c Expression of wound-responsive genes in ein2-1 mutant. The Col-0 and ein2-1 seedlings were submerged and wounded as described in Fig. 1c. Three biological replicates were averaged. Letters indicate groups that are statistically significantly different from each other (P < 0.05, Tukey’s test).
Submergence removes active histone modifications at the OPR3 locus
Hypoxia triggers changes of histone modifications in both animals and plants26,27. We therefore analysed histone modifications at the OPR3 locus using chromatin immunoprecipitation (ChIP) assays (Fig. 5a). H3K4me2 and acetylation of histone H3 (H3Ac) are considered as markers of active transcription41,42. ChIP assays showed that H3K4me2 was significantly decreased by submergence in both mock- and wound-treated seedlings (Fig. 5b; P6-mock, P < 1.0 × 10−7, Hedges′ g = 13.7; P6-wound, P = 0.0000001, Hedges′ g = 15.4). However, changes of H3K4me2 in mock-treated seedlings would be a technical error, because H3K4me2 was also reduced by submergence at the control ACTIN 7 (ACT7) locus (Fig. 5b). Otherwise, H3Ac was significantly suppressed by submergence in both mock- and wound-treated seedlings (P4-mock, P = 0.0000001, Hedges′ g = 10.8; P4-wound, P < 1.0 × 10−7, Hedges′ g = 11.6), while that did not exhibit any change in the control regions (Fig. 5c). We also analysed trimethylation of histone H3 lysine 9 (H3K9me3), which is associated with heterochromatin43, but the results did not yield meaningful data (Supplementary Fig. 5). These results suggest that submergence epigenetically regulates the OPR3 gene to inactivate gene expression before wounding. To find genes responsible for submergence-mediated histone modifications, we analysed expression of the OPR3 gene in the mutants that are deficient in a histone deacetylase (axe1-5)44, a histone demethylase (jmj30-2)45, and histone methyltransferases (atx1-2 atx2-1)46. However, expression of the OPR3 gene in these mutants was similar to that in Col-0 seedlings (Supplementary Fig. 6). We next treated Col-0 seedlings with trichostatin A (TSA), which is a histone deacetylase inhibitor, to examine the role of histone deacetylases in these responses. The expression of the OPR3 and JAZ10 genes in the seedlings that had undergone submergence was not altered by TSA treatment until 1 h after wounding (Fig. 5d). However, wounding slightly but significantly elevated expression of OPR3 and JAZ10 genes at 2 h after wounding in TSA-treated seedlings (OPR3, P = 0.0019, Hedges′ g = 24.1; JAZ10, P = 0.0000002, Hedges′ g = 21.5), whereas changes upon wounding were not significant in mock-treated seedlings (Fig. 5d). These results suggest that the histone deacetylase affects submergence-mediated inhibition at late stage of wound responses. All together, our findings suggest that even a short-term experience of submergence triggers repression of plant defence responses against herbivores. Active histone modifications at the OPR3 locus would enable wound-induced gene expression in plants grown under ambient air (Fig. 5e). However, submergence induces ethylene and other unknown responses to inhibit JA biosynthesis. In addition, submergence removes active histone modifications; thus, subsequent wounding cannot efficiently induce the expression of JA biosynthesis genes, resulting in reduced defence against herbivores (Fig. 5e).

a Genomic structure of the OPR3 gene. Black boxes, exons; white boxes, untranslated regions. Sequence elements used for ChIP assays are annotated as P1 to P7. b, c Histone modifications at OPR3 locus. Ten-day-old Col-0 seedlings were submerged for 1 h and then transferred to the air. Re-aerated seedlings were wounded immediately and harvested 1 h after the treatment. Anti-H3K4me2 (b) and anti-H3Ac (c) antibodies were used for immunoprecipitation. Three to four biological replicates were averaged. Error bars indicate ±SD. Letters indicate groups that are statistically significantly different from each other (P < 0.05, Tukey’s test). It is noteworthy that the data were statistically analysed separately for individual sequence elements by one-way ANOVA. Individual data points were marked in IP. IP, immunoprecipitation; NC, negative control. d Effects of TSA on wound-induced gene expressions after submergence. The Col-0 seedlings grown on DMSO- or TSA-containing MS-agar media were submerged for 1 h and then transferred to the air. Re-aerated seedlings were wounded and harvested at the indicated time points after the treatment. Three biological replicates were averaged. Letters indicate groups that are statistically significantly different from each other (P < 0.01, Tukey’s test). Error bars indicate ±SD. It is noteworthy that DMSO is used for mock treatment. e Schematic model for wound-induced defence responses after the submergence. In the ambient air, wounding induces expression of JA biosynthesis genes for defence against herbivores. When plants undergo submergence, ethylene signalling was activated and histone modifications were changed from the active to inactive states. There would be other unknown regulators (X) activated by submergence to inhibit wound responses. These events would block wound-induced gene expressions, resulting in reduced herbivore resistance.
Discussion
Many studies have identified the molecular signalling pathways for a single abiotic or biotic stress, but how previous environmental stresses affect upcoming biotic challenge is largely unknown. This work found that experience of submergence causes negative effects on plant resistance to herbivores by suppressing JA biosynthesis. Why do plants inactivate defence systems against herbivores after submergence? Previous studies have reported that JA antagonizes salicylic acid (SA) signalling. Activation of JA signalling results in inhibition of SA-dependent gene expressions in tobacco47. In addition, JA-insensitive coi1 mutant exhibits enhanced expression of SA-dependent genes and resistance to Pseudomonas syringae47. Thus, it is possible that the JA signalling pathways are inhibited in plants to improve SA-dependent pathogen resistance after submergence. In support of this idea, a recent study has reported that prior submergence enhances resistance to bacterial pathogen P. syringae by upregulating expression of a WRKY22 gene in Arabidopsis38. WRKY22 is a SA-responsive gene and high expression of WRKY22 causes negative effects on defence against aphids48. Further research on the JA-SA cross-talks would provide important clues regarding the molecular mechanisms on submergence-mediated suppression of plant wound responses.
A recent study has reported that AOS and hydroperoxide lyase (HPL)-mediated metabolic changes confer waterlogging tolerance in Arabidopsis49. Waterlogging induces production of 12-oxo-PDA (12-oxo-10,15(Z)-octadecatrienoic acid), which is a precursor of JA. However, JA content is not changed by waterlogging stress49. Our study also showed that submergence does not affect JA levels in the absence of wound stress (Fig. 2d). It is likely that plants induce JA-independent metabolic changes for tolerance to waterlogging stress by regulating AOS and HPL activities. On the other hand, plants reduce wound responses by suppressing JA biosynthesis possibly for increasing JA-antagonistic responses including pathogen resistance after submergence.
Ethylene is a key phytohormone in plant submergence responses. Diffusion rate of ethylene in water is lower than that in ambient air, thus submergence induces rapid ethylene responses in plants2,36. Our data suggested that the ethylene response is involved in submergence-mediated suppression of wound responses. Wound-induced expressions of JA-biosynthesis genes and JA-responsive genes were significantly suppressed by submergence in Col-0 seedlings, but those of several genes including AOC2, JAZ7, and VSP1 were partially restored in the ethylene-insensitive ein2-1 mutant. EIN2 is a key molecule in ethylene signalling. In the presence of ethylene, the ethylene receptors deactivate a negative regulator CONSTITUTIVE TRIPLE RESPONSE 1 to relieve suppression of EIN222. EIN2 protects downstream transcription factors including EIN3 and EIN3-LIKE 1 (EIL1) from the proteasomal degradation mediated by EIN3-BINDING F BOX PROTEIN 1 (EBF1) and EBF222,50. Therefore, it is possible that EIN2 mediates submergence-induced ethylene signalling to block JA biosynthesis. In support of this idea, previous studies have reported the JA-ethylene cross-talks at the molecular level. EIN3/EIL1 transcription factors physically interact with MYC2, a key transcription factor in JA signalling pathways51. The EIN3/EIL1-MYC2 interaction suppresses transcriptional activities of both EIN3/EIL1 and MYC2, showing antagonistic cross-talks between JA and ethylene. Because MYC2 activates JA-responsive promoters of JA biosynthesis genes52, the ethylene-mediated suppression of MYC2 could be the reason for the reduced expression of JA biosynthesis genes after submergence.
Our data showed that submergence rapidly suppresses plant wound responses with no signs of oxygen deprivation. These results indicate a crosstalk between wound responses and early signals of submergence rather than oxygen deprivation. Mutation of EIN2 partially restored wound responses after submergence, but still the expression of wound-responsive genes was largely reduced (Fig. 4c). Previous studies proposed that EIN2-independent pathways would be involved in ethylene signalling. Ethylene-insensitive phenotype of the ein2 mutant is restored by reducing JA levels53. Also, EIN2 TARGETING PROTEIN 1 (ETP1) and ETP2-deficient mutants exhibit constitutive stabilization of EIN2 but are still responsive to ethylene54, raising the possibility that an EIN2-independent ethylene pathway could be related with submergence-mediated suppression of wound responses. Otherwise, submergence could activate ethylene-independent regulators to inhibit wound responses. It is possible that the synergistic action of EIN2 and other unidentified regulators is required to fully suppress wound responses upon submergence.
Submergence-mediated removal of active histone modification at the OPR3 locus led us to investigate upstream regulators for the epigenetic regulation. However, the key genes for the histone modifications under submergence conditions were not identified. TSA treatment assays showed that histone deacetylases are at least partially involved in suppression of late stage wound responses after submergence (Fig. 5d), but the effects of TSA were not enough to fully explain molecular mechanisms of a crosstalk between submergence and wound responses. These results suggest that multiple histone modifications might be redundantly involved in these responses. Genetic analyses using mutants deficient in multiple histone modifications would be required to find clues for the detailed mechanisms.
Methods
Plant materials and growth conditions
Arabidopsis thaliana ecotype Columbia (Col-0) was used for the assays. The atx1-2 atx2-1 (N71692) seeds were obtained from the Nottingham Arabidopsis Stock Centre (NASC, Nottingham, UK). The ein2-1 and axe1-5 seeds were a gift from Dr. Young-Joon Park and previously described44,55. The jmj30-2 seeds were a gift from Dr. Pil Joon Seo and previously described45. Seeds were surface-disinfected using 70% ethanol and then incubated at 4 °C for 3 days. After stratification, seeds were transferred to a growth room set at 24 °C and at 50% humidity. Seedlings were grown on half-strength Murashige and Skoog-agar (MS-agar) plates containing 0.7% agar without sucrose under long days (16 h light and 8 h dark cycles). Plates were placed horizontally in the growth room. White light with an intensity of about 100 μmol m−2 s−1 was applied using fluorescent FL40EX-D tubes (Focus, Bucheon, Korea).
Submergence treatment
Col-0 seedlings grown on MS-agar plates for 10 days under long days were used. Lids of the plates were opened before submergence. Seedlings were submerged in distilled water for the indicated time points in the figure legends. During submergence, plates were placed horizontally in the water-filled transparent box without lids. Submergence treatment was performed in the growth room in light. After submergence, plates were transferred to air and waters in the plates were poured out and lids were closed. To analyse wound responses, cotyledons and first leaves of the seedlings were wounded with scissors right after submergence.
Reverse-transcription quantitative PCR
Total RNA was extracted from the seedlings after appropriate treatments using RNeasy Plant Mini Kit (QIAGEN) with an on-column DNase I treatment. Reverse-transcription reaction was performed to produce cDNA using TOPscript cDNA Synthesis Kit (Enzynomics). Reverse-transcription quantitative PCR (RT-qPCR) reactions were carried out in 96-well plates with a CFX Connect Real-Time PCR Detection System (Bio-Rad) using TOPreal qPCR PreMIX (Enzynomics). The primers used in RT-qPCR reactions were listed in Supplementary Table 1. A reference gene UBC21 (AT5G25760) was included as an internal control for normalization. The comparative ΔΔCT method was used to evaluate relative quantities of each amplified product in the samples. The threshold cycle (CT) was automatically determined for each reaction by the system set with default parameters.
Histochemical assays
A 2112 bp fragment containing the promoter and the 5′-untranslated region of the JAZ10 gene was fused to the 5′-end of the β-glucuronidase (GUS)-coding gene in the vector pCAMBIA1305.229. The fusion construct was transformed into Col-0 plants. We obtained three independent second generation (T2) pJAZ10:GUS transgenic plants and confirmed wound-responsive GUS expression in all transgenic lines. A single line was used for histochemical GUS assays to observe effects of submergence. Transgenic seedlings grown on MS-agar plates for 10 days under long days were submerged for 1 h and then transferred to air. First leaves of reaerated plants were wounded and incubated at 24 °C under the light for 1 h and then fixed in 90% acetone for 20 min on ice. Seedlings were washed twice with rinsing solution containing 50 mM sodium phosphate pH 7.2, 0.5 mM K3Fe(CN)6, and 0.5 mM K4Fe(CN)6. Fixed seedlings were incubated at 37 °C for 16 h in staining solution containing 2 mM X-Gluc (Duchefa) in the rinsing solution. After the staining, seedlings were incubated in 70% ethanol for 4 h. For 3,3′-diaminobenzidine (DAB) staining, seedlings were submerged for 1 h and then transferred to air. First leaves of reaerated plants were wounded in the air. After 30 min, seedlings were incubated in DAB staining solution containing 1 mg/ml DAB, 10 mM Na2HPO4, and 0.05% (v/v) Tween 20 at 24 °C for 16 h in darkness. After staining, DAB staining solution was removed and the seedlings were incubated in 70% ethanol for 4 h. Plant materials were mounted on slide glasses and were photographed using Nikon D5600 digital camera (Nikon).
Hypoxia treatment
Transparent glove box containing two holes for gas flow (GBI) was used for hypoxia treatment. Col-0 seedlings grown on MS-agar plates at 24 °C for 10 days under long days were placed in the glove box with lid open and the pressure was held at 0.6 MPa with flow of 99.999% (v/v) argon. Seedlings were incubated in the box for 2 h under light conditions and then were wounded in ambient air.
Pharmacological treatment
For MeJA treatment, Col-0 seedlings grown on MS-agar plates at 24 °C for 10 days under long days were submerged for 1 h and then sprayed with 25 μM MeJA (Sigma-Aldrich/Merck). For glutamate treatment, Col-0 seedlings grown on MS-agar plates were submerged for 1 h and then sprayed with 100 mM l-glutamate (Sigma-Aldrich/Merck). For TSA treatment, Col-0 seedlings were grown on MS-agar plates for 2 days. Seedlings were then transferred to other MS-agar plates containing 5 μM TSA and further grown for 8 days. As TSA is dissolved in dimethyl sulfoxide (DMSO), DMSO-treated seedlings were used as a mock control. Seedlings were submerged for 1 h and then transferred to air. Re-aerated seedlings were wounded and then harvested at the annotated time points after wounding.
Confocal microscopy
The iGluSnFR fragment (from Addgene plasmid #41732) was amplified by PCR and inserted into pBA002 vector containing CaMV 35S promoter and NOS terminator. The 63 bp of chitinase (AT3G12500) signal peptide was fused in-frame to the 5′-end of the iGluSnFR fragment, forming 35S:CHIB-iGluSnFR construct8. The construct was transformed into Col-0 plants. We obtained three independent second generation (T2) 35S:CHIB-iGluSnFR transgenic plants and confirmed wound-responsive iGluSnFR fluorescence signals in all transgenic lines. A single line was used for detailed analysis to observe effects of submergence. Transgenic seedlings were submerged for 1 h and then wounded in the air. After 5 min, seedlings were placed on slide glasses and were subjected to fluorescence imaging using an LSM 800 confocal microscope (Carl Zeiss). Fluorescence images were analysed using ZEN 2.5 LITE software. Fluorescence intensities were measured using ImageJ software.
ChIP assays
Col-0 seedlings grown on MS-agar plates at 24 °C for 10 days under long days were used for ChIP assays. Seedlings were submerged in distilled water for 1 h and then transferred to air. Cotyledons and first leaves of reaerated plants were wounded in air. After 1 h, seedlings were vacuum infiltrated with 1% (v/v) formaldehyde for cross-linking. The cross-linking was quenched by adding glycine. Plant materials were then ground in liquid nitrogen and resuspended in nuclear extraction buffer containing 1.7 M sucrose, 10 mM Tris-Cl pH 7.5, 2 mM MgCl2, 0.15% (v/v) Triton X-100, 5 mM β-mercaptoethanol, 0.1 mM phenylmethylsulfonyl fluoride, and protease inhibitor cocktail tablets (Sigma-Aldrich/Merck). Nuclear fraction was isolated by centrifuge at 16,000 × g for 1 h at 4 °C. The nuclear fraction was lysed with lysis buffer containing 50 mM Tris-Cl pH 8.0, 0.5 M EDTA, 1% (w/v) SDS, and protease inhibitor cocktail tablets. Solutions were then sonicated to fragment chromatins into 0.5 to 0.8 kb. The sonicated solutions were centrifuged at 16,000 × g for 10 min at 4 °C. The supernatants were transferred to new tubes and diluted (1:10) in ChIP dilution buffer containing 1.1% (v/v) Triton X-100, 1.2 mM EDTA, 16.7 mM Tris-Cl pH 7.5, and 167 mM NaCl. The 10% (v/v) of the diluted solutions were used as inputs. Anti-H3K4me2, anti-H3Ac, and anti-H3K9me3 antibodies were added for immunoprecipitation of histone-DNA complexes. Chromatin solutions without antibodies were used as negative controls. The precipitates were collected using protein G magnetic beads (Bio-Rad) and then eluted from the beads to isolate DNA fragments. DNAs in inputs, negative controls, and immunoprecipitated samples were purified using Wizard® SV gel and PCR clean-up system (Promega). Genomic DNA enriched in the chromatin preparations was analysed using qPCR. The qPCR signals derived from the negative controls and immunoprecipitated samples were normalized using those derived from the inputs. The primers used are listed in Supplementary Table 1.
Herbivore-feeding assays
Col-0 plants grown in soil at 24 °C for 4 weeks under long days were submerged in distilled water for 1 h before releasing Brassicaceae specialist P. rapae caterpillars (Larval stage L1) (Hampyeongnabi). Larval weights were measured before and 5 days after. Soft paintbrush was used to move caterpillars. Twenty caterpillars were released to each experiment and seven to ten survived caterpillars were weighed after 5 days.
Statistics and reproducibility
All statistical methods are annotated in the figure legends. Each experiment was repeated at least twice with similar results. The numbers of biological replicates in each assays are also indicated in the figure legends. ImageJ software was used for quantification of GUS, DAB, and iGluSnFR signals. One-way analysis of variance with post hoc Tukey’s test was performed using Rstudio software. For gene expression analysis, three biological replicates with ~10 seedlings for each replicate were used. For analysing GUS signals, four biological replicates with a single leaf for each replicate were used. For measuring JA content, five biological replicates with ~100 mg of seedlings for each replicate were used. For herbivore-feeding assay, weights of 8 to 24 caterpillars were analysed. For ChIP assays, four biological replicates with ~20 seedlings for each replicate were used. Note that three replicates were used for analysing P7 locus as a negative control. Bar and box plots were made using Microsoft Excel. Individual data points were included in all plots. In box plots, each box extends from the first quartile to the third quartile values of the data, and centre lines show median values. Whiskers indicate 1.5 times the interquartile range. Black dots indicate outliers.
Reporting summary
Further information on experimental design is available in the Nature Research Reporting Summary linked to this paper.
Data availability
All data are available in the main text or the Supplementary Information. All the source data for graphs in Figures and Supplementary Information are presented in Supplementary Data 1–11. The Arabidopsis Genome Initiative locus codes for the genes discussed in this study are as follows: JAZ7 (AT2G34600), JAZ10 (AT5G13220), OPR3 (AT2G06050), AOC2 (AT3G25770), LOX2 (AT3G45140), AOC1 (AT3G25760), AOC3 (AT3G25780), AOC4 (AT1G13280), AOS (AT5G42650), LOX3 (AT1G17420), LOX4 (AT1G72520), LOX6 (AT1G67560), TCH2 (AT5G37770), TCH3 (AT2G41100), ADH1 (AT1G77120), ABR1 (AT5G64750), PDF1.2 (AT5G44420), JR1 (AT3G16470), PR2 (AT3G57260), PR5 (AT1G75040), VSP1 (AT5G24780), ERF4 (AT3G15210), ACT7 (AT5G09810), and UBC21 (AT5G25760).
References
Voesenek, L. A., Colmer, T. D., Pierik, R., Millenaar, F. F. & Peeters, A. J. How plants cope with complete submergence. N. Phytol. 170, 213–226 (2006).
Fukao, T., Barrera-Figueroa, B. E., Juntawong, P. & Pena-Castro, J. M. Submergence and waterlogging stress in plants: a review highlighting research opportunities and understudied aspects. Front. Plant Sci. 10, 340 (2019).
Schmidt, R. R., Weits, D. A., Feulner, C. F. J. & van Dongen, J. T. Oxygen sensing and integrative stress signaling in plants. Plant Physiol. 176, 1131–1142 (2018).
Dyck, V. A. et al. In Brown Planthopper: Threat to Rice Production in Asia (ed. Brady, N. C.) 61–98 (IRRI, 1979).
Nakamura, M., Utsumi, S., Miki, T. & Ohgushi, T. Flood initiates bottom-up cascades in a tri-trophic system: host plant regrowth increases densities of a leaf beetle and its predators. J. Anim. Ecol. 74, 683–691 (2005).
Li, H., Syvertsen, J. P., McCoy, C. W., Stuart, R. J. & Schumann, A. W. Water stress and root injury from simulated flooding and diaprepes abbreviatus root weevil larval feeding in citrus. Soil Sci. 171, 138–151 (2006).
Miller, G. et al. The plant NADPH oxidase RBOHD mediates rapid systemic signaling in response to diverse stimuli. Sci. Signal 2, ra45 (2009).
Toyota, M. et al. Glutamate triggers long-distance, calcium-based plant defense signaling. Science 361, 1112–1115 (2018).
Gilroy, S. et al. ROS, calcium, and electric signals: key mediators of rapid systemic signaling in plants. Plant Physiol. 171, 1606–1615 (2016).
Mosblech, A. et al. Phosphoinositide and inositolpolyphosphate signalling in defense responses of Arabidopsis thaliana challenged by mechanical wounding. Mol. Plant 1, 249–261 (2008).
Suza, W. P. & Staswick, P. E. The role of JAR1 in Jasmonoyl-L: -isoleucine production during Arabidopsis wound response. Planta 227, 1221–1232 (2008).
Heil, M. et al. How plants sense wounds: damaged-self recognition is based on plant-derived elicitors and induces octadecanoid signaling. PLoS ONE 7, e30537 (2012).
Thines, B. et al. JAZ repressor proteins are targets of the SCF(COI1) complex during jasmonate signalling. Nature 448, 661–665 (2007).
Halitschke, R. & Baldwin, I. T. Antisense LOX expression increases herbivore performance by decreasing defense responses and inhibiting growth-related transcriptional reorganization in Nicotiana attenuata. Plant J. 36, 794–807 (2003).
Savatin, D. V., Gramegna, G., Modesti, V. & Cervone, F. Wounding in the plant tissue: the defense of a dangerous passage. Front. Plant Sci. 5, 470 (2014).
Perata, P. & Voesenek, L. A. C. J. Submergence tolerance in rice requires Sub1A, an ethylene-response-factor-like gene. Trends Plant Sci. 12, 43–46 (2007).
Sasidharan, R. et al. Signal dynamics and interactions during flooding stress. Plant Physiol. 176, 1106–1117 (2018).
Drew, M. C. Oxygen deficiency and root metabolism: injury and acclimation under hypoxia and anoxia. Annu. Rev. Plant Physiol. Plant Mol. Biol. 48, 223–250 (1997).
Fukao, T., Xu, K., Ronald, P. C. & Bailey-Serres, J. A variable cluster of ethylene response factor-like genes regulates metabolic and developmental acclimation responses to submergence in rice. Plant Cell 18, 2021–2034 (2006).
Bailey-Serres, J. & Voesenek, L. A. C. J. Flooding stress: acclimations and genetic diversity. Annu. Rev. Plant Biol. 59, 313–339 (2008).
Jackson, M. B. Ethylene-promoted elongation: an adaptation to submergence stress. Annu. Bot. 101, 229–248 (2008).
Ji, Y. & Guo, H. From endoplasmic reticulum (ER) to nucleus: EIN2 bridges the gap in ethylene signaling. Mol. Plant 6, 11–14 (2013).
Peng, H. P., Chan, C. S., Shih, M. C. & Yang, S. F. Signaling events in the hypoxic induction of alcohol dehydrogenase gene in Arabidopsis. Plant Physiol. 126, 742–749 (2001).
Tsai, K. J., Chou, S. J. & Shih, M. C. Ethylene plays an essential role in the recovery of Arabidopsis during post-anaerobiosis reoxygenation. Plant Cell Environ. 37, 2391–2405 (2014).
Hong, C. P., Wang, M. C. & Yang, C. Y. NADPH oxidase RbohD and ethylene signaling are involved in modulating seedling growth and survival under submergence stress. Plants 9, 471 (2020).
Batie, M. et al. Hypoxia induces rapid changes to histone methylation and reprograms chromatin. Science 363, 1222–1226 (2019).
Tsuji, H., Saika, H., Tsutsumi, N., Hirai, A. & Nakazono, M. Dynamic and reversible changes in histone H3-Lys4 methylation and H3 acetylation occurring at submergence-inducible genes in rice. Plant Cell Physiol. 47, 995–1003 (2006).
Franks, P. J. & Farquhar, G. D. The mechanical diversity of stomata and its significance in gas-exchange control. Plant Physiol. 143, 78–87 (2007).
Acosta, I. F. et al. Role of NINJA in root jasmonate signaling. Proc. Natl Acad. Sci. USA 110, 15473–15478 (2013).
Koo, A. J. K., Gao, X., Jones, A. D. & Howe, G. A. A rapid wound signal activates the systemic synthesis of bioactive jasmonates in Arabidopsis. Plant J. 59, 974–986 (2009).
Gfeller, A. et al. Jasmonate controls polypeptide patterning in undamaged tissue in wounded Arabidopsis leaves. Plant Physiol. 156, 1797–1807 (2011).
Pauwels, L. et al. NINJA connects the co-repressor TOPLESS to jasmonate signalling. Nature 464, 788–791 (2010).
Wasternack, C. & Song, S. Jasmonates: biosynthesis, metabolism, and signaling by proteins activating and repressing transcription. J. Exp. Bot. 68, 1303–1321 (2017).
Caarls, L. et al. Arabidopsis JASMONATE-INDUCED OXYGENASES down-regulate plant immunity by hydroxylation and inactivation of the hormone jasmonic acid. Proc. Natl Acad. Sci. USA 114, 6388–6393 (2017).
Braam, J. Regulated expression of the calmodulin-related TCH genes in cultured Arabidopsis cells: induction by calcium and heat shock. Proc. Natl Acad. Sci. USA 89, 3213–3216 (1992).
Liu, F. et al. Global transcription profiling reveals comprehensive insights into hypoxic response in Arabidopsis. Plant Physiol. 137, 1115–1129 (2005).
Vashisht, D. et al. Natural variation of submergence tolerance among Arabidopsis thaliana accessions. N. Phytol. 190, 299–310 (2011).
Hsu, F. C. et al. Submergence confers immunity mediated by the WRKY22 transcription factor in Arabidopsis. Plant Cell 25, 2699–2713 (2013).
Suzuki, K., Suzuki, N., Ohme-Takagi, M. & Shinshi, H. Immediate early induction of mRNAs for ethylene-responsive transcription factors in tobacco leaf strips after cutting. Plant J. 15, 657–665 (1998).
Bäumler, J. et al. AtERF#111/ABR1 is a transcriptional activator involved in the wounding response. Plant J. 100, 969–990 (2019).
Moghaddam, A. M. B. et al. Additive inheritance of histone modifications in Arabidopsis thaliana intra-specific hybrids. Plant J. 67, 691–700 (2011).
Keren, I. & Citovsky, V. Activation of gene expression by histone deubiquitinase OTLD1. Epigenetics 12, 584–590 (2017).
Becker, J. S., Nicetto, D. & Zaret, K. S. H3K9me3-dependent heterochromatin: barrier to cell fate changes. Trends Genet 32, 29–41 (2016).
Murfett, J., Wang, X. J., Hagen, G. & Guilfoyle, T. J. Identification of Arabidopsis histone deacetylase HDA6 mutants that affect transgene expression. Plant Cell 13, 1047–1061 (2001).
Lee, K., Park, O. S. & Seo, P. J. JMJ30-mediated demethylation of H3K9me3 drives tissue identity changes to promote callus formation in Arabidopsis. Plant J. 95, 961–975 (2018).
Xu, M., Leichty, A. R., Hu, T. & Poethig, R. S. H2A.Z promotes the transcription of MIR156A and MIR156C in Arabidopsis by facilitating the deposition of H3K4me3. Development 145, dev152868 (2018).
Kunkel, B. N. & Brooks, D. M. Cross talk between signaling pathways in pathogen defense. Curr. Opin. Plant Biol. 5, 325–331 (2002).
Kloth, K. J. et al. AtWRKY22 promotes susceptibility to aphids and modulates salicylic acid and jasmonic acid signalling. J. Exp. Bot. 67, 3383–3396 (2016).
Savchenko, T., Rolletschek, H., Heinzel, N., Tikhonov, K. & Dehesh, K. Waterlogging tolerance rendered by oxylipin-mediated metabolic reprogramming in Arabidopsis. J. Exp. Bot. 70, 2919–2932 (2019).
Zhao, Q. & Guo, H. W. Paradigms and paradox in the ethylene signaling pathway and interaction network. Mol. Plant 4, 626–634 (2011).
Song, S. et al. Interaction between MYC2 and ETHYLENE INSENSITIVE3 modulates antagonism between jasmonate and ethylene signaling in Arabidopsis. Plant Cell 26, 263–279 (2014).
Wasternack, C. & Hause, B. Jasmonates: biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot. 111, 1021–1058 (2013).
Kim, J., Patterson, S. E. & Binder, B. M. Reducing jasmonic acid levels causes ein2 mutants to become ethylene responsive. FEBS Lett. 587, 226–230 (2013).
Cho, Y. H., Lee, S. & Yoo, S. D. EIN2 and EIN3 in ethylene signaling. Annu. Plant Rev. 44, 169–187 (2012).
Guzmán, P. & Ecker, J. R. Exploiting the triple response of Arabidopsis to identify ethylene-related mutants. Plant Cell 2, 513–523 (1990).
Acknowledgements
We thank Dr. Young-Joon Park and Dr. Pil Joon Seo for sharing seeds, Dr. Kwangsoo Shin for building hypoxia treatment systems. This work was supported by the Basic Research Program provided by the National Research Foundation of Korea (NRF- 2019R1C1C1002045), New Breeding Technologies Development Program (Project number PJ01480201) provided by the Rural Development Administration of Korea, and the KRIBB Research Initiative Program (KGM5372012).
Author information
Authors and Affiliations
Contributions
H.-J.L. conceived and designed the experiments. H.-J.L. performed gene expression analysis, DAB staining, and confocal microscopy. H.-J.L. and J.-S.P. performed ChIP assays. H.-J.L. and S.Y.S. performed herbivore-feeding assay. S.-G.K. and G.L. measured JA content. H.-S.K. and J.H.J. provided experimental tools and equipment. H.-J.L. prepared the manuscript with the contributions of H.-S.K. and H.S.C.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lee, HJ., Park, JS., Shin, S.Y. et al. Submergence deactivates wound-induced plant defence against herbivores. Commun Biol 3, 651 (2020). https://doi.org/10.1038/s42003-020-01376-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s42003-020-01376-4
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
