
Abstract
Identifying the environmental factors that determine the occurrence of invasive species is essential in defining and implementing effective control campaigns. Here, we applied multi-season occupancy models to analyze American mink (Neogale vison) track data collected using 121 floating rafts, as a function of factors occurring at multiple spatial scales. Our overall aim was to identify those factors that determine the use, colonization or abandonment of rafts by free ranging individuals found in western Macedonia, Greece. We found that increasing values of shrubs and rock cover at the micro-habitat scale were positively associated with the species’ probability of raft use, as was the density of medium-sized rivers at the landscape scale. Colonization was found to increase with increasing amounts of shrub and reed cover; however, both variables were not informative. Conversely, the distance from the nearest fur farm was highly informative in predicting raft abandonment by the species. Effective control actions may require removal by trapping along rocky or densely vegetated riverbanks or lake shores located in the vicinity of the established fur farms in the area. Habitat management, although possible, may be difficult to implement due to the ability of the species to adapt. Finally, fur farms should maximize security and establish an early warning and rapid eradication system in case of future escapes.
Similar content being viewed by others

Introduction
Over the past two centuries, human activities have led to a significant surge in the translocation of species outside their native range1,2,3. While many of these species play a crucial role in our global economy, the negative impacts of nonnative species invasions on native biodiversity are profound4,5. In Europe, the rate of invasion by nonnative species is expected to increase in the near future5. As a consequence, comprehending the mechanisms and patterns of biotic invasions has been the focus of many recent studies.
Native to North America, the American mink (Neogale vison) is a generalist, territorial, semiaquatic carnivore6. The species was first introduced to Europe in the 1920s as a furbearer7 and has since colonized many parts of the world through accidental and deliberate releases8. Among European countries, Greece holds one of the largest fur industries, with mink being bred for their fur since the 1970s9. Currently, there are more than 100 mink fur farms operating in the general region of western Macedonia10.
Feral mink populations are believed to have been established in western Macedonia following deliberate releases by so-called animal rights activists. More specifically, in 2010, approximately 50,000 mink were released, followed by another 10,000 in the year ahead. While most individuals were either recaptured or killed by passing vehicles, an unknown number of individuals became naturalized and were able to spread to the surrounding landscape through the Aliakmonas River valley. Despite the immense amount of scientific evidence suggesting that the American mink may cause conservation problems for local species11,12,13,14,15, no measures were taken at that time to prevent further spread or protection of native fauna. For example, mink may decimate entire colonies and populations of ground nesting birds, including several species of the Anatidae and Rallidae family16, many of which can be found in the general region of western Macedonia. Other affected species include reptiles and amphibians commonly found in riparian ecosystems7,17, whereas it may also affect directly or indirectly other aquatic predators such as the river otter (Lutra lutra)18,19, which is also present in the wider region. However, it was not until 2019 that a LIFE project (LIFE18 NAT/GR/000430) addressed this issue, following the example of many other European countries that have control policies and eradication campaigns focused on American mink7.
Although the ecology of mink outside its native range is well documented, information is lacking in southeastern Europe20. Previous studies have shown that mink can occupy areas as far as ~ 1 km away from water sources21, moving along banks with dense vegetation and complex ground structures where they can hide or den in, while they avoid open areas and high slopes22,23,24,25,26,27. In addition, mink presence may be influenced by biotic factors such as prey availability, competition and predation28,29,30. However, due to its ability to adapt to novel environments, the factors associated with its presence may vary among different regions. In Greece, the species has been poorly studied. In a recent study, Galanaki and Kominos31 investigated mink distribution. However, their findings were based on opportunistic data and offered little to no information regarding the factors driving mink presence in the region. In addition, Vada et al.20 concluded that in order to align management objectives and coordinate mink control across countries, mink monitoring should follow a standardized protocol at the continental scale.
In this study, we applied multi-season occupancy models to analyze American mink detection/non-detection data collected using 121 floating rafts, as a function of factors occurring at multiple spatial scales. Our overall aim was to identify those factors that determine the use, colonization or abandonment of rafts by free ranging individuals found in western Macedonia, Greece, and provide management recommendations for controlling the species.
Methods
Study area
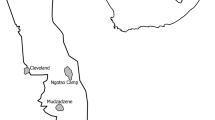
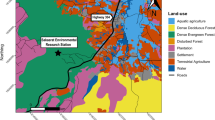
The study was carried out between 2021 and 2023 within the Aliakmon River Basin and the Prespa Sub-basin. The Aliakmon River Basin included approximately 140 km stretches of river as well as lake Orestias. whereas the Prespa Sub-basin only included the Lake Mikri Prespa (Fig. 1). The Aliakmon River Basin covers an area of approximately 11,000 km2 and contains Greece’s largest river, the Aliakmon River32. The area is principally covered by forests and seminatural areas (57%), followed by agricultural areas (37.6%), urban areas (3.3%), water and wetlands (2.1%). Similarly, the Prespa Sub-basin covers an area of 347 km2 and is principally covered by forests (62%), followed by water (23.5%), agricultural land (7.7%), pastures (1.4%), and urban areas (1.1%)33.

Study area and site locations of the 121 floating rafts used to track American mink presence in western Macedonia, Greece, 2021–2023.
Floating raft surveys
American mink detection/non-detection data were gathered from track surveys carried out with the use of mink floating rafts, following pre-established protocols34, and in accordance with Greece animal welfare legislation. A detailed description of the floating raft can be found in Reynolds et al.34. In brief, a floating raft acts as a monitoring device by recording footprints on a clay cartridge. A total of 150 rafts were established within the study area, however data from 121 rafts were included in the analysis. Rafts were spaced at approximately 1 km intervals (mean = 823 m; range: 183–3965 m) along the Aliakmon River and its tributaries, as well as around Lakes Orestias and Mikri Prespa. Spacing between rafts was based on three different criteria; increase the probability of an individual being captured35,36, previous knowledge of mink home range size in Mediterranean regions37, and ease of access by the observers. Rafts were left in place for the entire study period. All tracks from each visit were photographed by the observers and sent to experts for identification. Data were recorded as mink presence or absence, depending on whether we found an imprint of mink paws on each clay cartridge during each visit.
Raft surveys were conducted in four distinct primary periods: September–November 2021, April–May 2022, October–December 2022, and March–April 2023. Each primary period was concluded within a time window of 25 days, during which rafts were set to record mink tracks for five consecutive days, followed by a 5-day trapping campaign at selected rafts (with recorded mink presence and immediate adjacent ones), and so on and so forth. Hence, during each primary period we recorded mink presence or absence in three distinct secondary periods, each of which lasted five consecutive days (total of 15 days of tracking required for each primary period).
Ethical statement
All procedures carried out in this study were conducted in accordance with Greece animal welfare legislation, approved by the General Secretariat of Natural Environment and Water, Ministry of Environment and Energy, Hellenic Republic (Licence number: 75935/1924/16-12-2020) and conducted under the authority of the project licence (LIFE18 NAT/GR/000430).
Environmental covariates
To better understand the associations between habitat features and mink occupancy, we followed a multiscale approach that included micro-habitat, local and landscape scale variables. At the micro-habitat scale, we used a 10 m transect along each of the main cardinal directions to estimate vegetation cover and cover of different ground components at each raft location21,26. Vegetation cover was estimated as the percent cover of three different strata (Table 1), with data recorded every 2 m. The percent cover of different ground components included bare soil, rocks, grass, shrubs and reeds, with data recorded every 50 cm (Table 1). Finally, we extracted elevation values at each site using a hand-held GPS device. Local and landscape variables were created using ArcGIS Desktop: Release 1038 and cartographic data from the Copernicus Land Monitoring Services. Variables included the percent cover of different land uses, river density and distance to the nearest fur farms (Table 1). Local variables referred to a distance class of 250 m around each raft, whereas landscape variables referred to a distance class of 1 km around each raft. All of the above variables were also used to account for temporal variation in colonization and extinction, along with some yearly covariates which included the effect of primary period (S), whether trapping was attempted or not (trap), and the number of individuals caught (catch) (Table 1). Finally, to account for imperfect detection, we included four observation-level covariates (Table 1). Prior to model fitting, all variables were checked for collinearity and were standardized to a mean of zero and a variance of one. Because variables “Strata 1” and percent (%) shrub cover (SH) were strongly correlated (Pearson’s r > 0.6), variable “Strata 1” was excluded from further analyses.
Analysis framework and dynamic occupancy models
We used multi-season dynamic occupancy models to investigate patterns of mink raft use after accounting for imperfect detection39. However, for these models to apply, three main assumptions must be met39 which are clarified below. According to the first assumption, there should be no false positive detections of the target species. In this study, all animal tracks were photographed and sent to experts for identification and hence false positive detections of minks should not be considered an issue. Based on the second assumption, sample units (rafts) are assumed to be independent. Since the study focused on mink tracks detection rather than detection of the species itself, the detection probability of tracks is independent on whether minks are in close proximity to the raft locations or not, and thus the second assumption could be considered true. Finally, according to the third assumption the status of a sampling unit (raft) should remain unchanged during primary periods40. In our case, the terms “occupancy” and “extinction” need to be clarified41,42,43. Occupancy estimates should be interpreted in terms of rafts used by the species; that is rafts where mink were present at some point during or prior to the survey period, as opposed to rafts exclusively inhabited during the survey period. Additionally, since disappearance from a raft implies abandonment rather than local extinction, the probability of extinction should be interpreted as probability of raft abandonment. Thus, we hereafter use the terms “use” instead of “occupancy” and “abandonment” instead of “extinction”.
Based on the above, we modelled the probability of raft use for the first primary period (ψ1), the probability of raft colonization (γ; i.e. the probability of an unused raft at time t becoming used at time t + 1) and raft abandonment (ε), and mink detection probability (p). In order to reduce the effect of uninformative parameters44, we developed a limited number of candidate models following a hierarchical four-stage approach45. First, we modelled p through a set of models that included each of the observation-level covariates (Table 1), as well as a null model in which no covariates were included (Supplementary Table S1). Second, we used the top ranked p model to develop a set of models examining ψ1 for each spatial scale (Supplementary Tables S2–S4). We then used the covariates included in the top ranked model from each spatial scale to develop a new list of candidate models for ψ1 (Supplementary Table S5). Third, we used the top ranked ψ1 model from the previous step to construct a list of candidate models examining γ for each spatial scale and for a set of yearly covariates that included the effect of primary period (S), attempted trapping (trap), number of individuals trapped (catch), as well as a null model (Supplementary Tables S6–S9). We then used the covariates from the above top ranked models to create a new list of candidate models for γ (Supplementary Table S10). Finally, we used the top ranked γ model from the previous step to create a set of models examining ε for each spatial scale and for the set of available yearly covariates (Supplementary Tables S11–S14). Using the covariates of the above top ranked models, we created a new list of candidate models for ε (Supplementary Table S15). The top ranked model from this step was the selected as the final model on which the interpretation of our results was based. Model fit of the final model was tested using a parametric bootstrap goodness-of-fit test based on Pearson’s χ2, where P > 0.05 indicates adequate model fit46,47. All statistical analyses were carried out in R48, with the function colext in the package unmarked46.
Results
Throughout the duration of the study, mink tracks were recorded at least once at 65 floating rafts, whereas a total of 28 individuals were trapped and culled. More specifically, mink tracks were recorded at 38 rafts and 18 individuals were trapped during the first primary period. During the second period mink tracks were recorded at 18 rafts and two individuals were trapped, followed by 27 rafts with mink tracks and five trapped individuals during the third period. Finally, during the fourth and final period, mink tracks were recorded at 18 rafts and three individuals were caught. The rafts used by the species during each period, as well as the rafts in which individuals were trapped are shown in Fig. 2a–d.

Location of used and unused floating rafts, as well as rafts with American mink captures during (a) the first, (b) the second, (c) the third, and (d) the fourth primary period that took place in western Macedonia, Greece, 2021–2023.
Based on the results of the final model (Table 2), American mink detectability was influenced by the month during which surveys were conducted. More specifically, detectability was greater during the months of September (\(\hat{\beta }\) = 1.287, S.E. = 0.487) and October (\(\hat{\beta }\) = 0.768, S.E. = 0.323), whereas during April, detectability was the lowest (\(\hat{\beta }\) = -1.395, S.E. = 0.255). Detection probabilities for each month are shown in Fig. 3. The percent amount of shrub (\(\hat{\beta }\) = 1.726, S.E. = 0.587) and rock cover (\(\hat{\beta }\) = 1.244, S.E. = 0.481) at the micro-habitat scale, as well as the density of medium-sized rivers (\(\hat{\beta }\) = 1.985, S.E. = 0.762) at the landscape scale, were found to have a positive effect on the probability of a raft being used by the species (Fig. 4a, b, c). Confidence intervals of all three variables did not span zero, suggesting that they were highly informative (Table 2). Conversely, the probability of a raft being colonized by the species increased with increasing cover of shrubs and reeds at the micro-habitat scale; however, both these variables exhibited wide confidence intervals (Table 2), suggesting that the probability of raft colonization was not well predicted at higher values of shrub and reed cover. Finally, the probability of a raft being abandoned by the species was positively associated with the distance to the nearest mink fur farm (\(\hat{\beta }\) = 1.345, S.E. = 0.52), with longer distances resulting in greater probabilities of raft abandonment (Fig. 5). Confidence intervals for this variable did not span zero (Table 2), suggesting that the probability of raft abandonment was well predicted at longer distances from fur farms. The final multi-season occupancy model from which the results were drawn had an adequate model fit according to Pearson’s χ2 (P = 0.110).

Predicted relationship between survey months and detection probabilities (± 95% CI) of American mink in western Macedonia, Greece, 2021–2023.

Predicted relationships between (a) shrub cover, (b) rock cover, and (c) medium river density and the probabilities of initial raft use by the American mink in western Macedonia, Greece, 2021–2023. Shaded areas represent confidence intervals (95%) of the estimations.

Predicted relationship between distance to the nearest fur farm and probability of local raft abandonment by the American mink in western Macedonia, Greece, 2021–2023. Shaded areas represent confidence intervals (95%) of the estimations.
Discussion
The ability of invasive species to successfully spread and establish in an area depends on their spatial and temporal responses to environmental features49. Occupancy models offer a great opportunity to estimate the factors that determine an invasive species’ presence in space and time, and their outputs can be utilized in designing and guiding management actions21,50,51,52.
Floating rafts constitute one of the most widespread methods for surveying American mink presence in riparian and coastal areas35,36,53,54,55. According to Reynolds et al.34 and Reynolds et al.56, floating rafts are much more efficient at tracking American mink than alternative methods; however, detection may vary by season. Indeed, the probability of detecting mink in our study area varied by month, with September and October exhibiting the highest probabilities. During this period, juvenile individuals disperse, and animals re-settle in territories57, whereas in late October, populations are generally stable58, which might explain the higher detection estimates. Similarly, Roy et al.59 reported a higher rate of trapped mink during this time. Conversely, individuals were less likely to be detected during the months of April and May. These months correspond to the gestation, parturition and weaning periods, during which mink activity is at its lowest60,61. When accounting for imperfect detection, mink occupancy within our study area was estimated to be 67.59%.
Initial use of rafts by the American mink within the study area was found to be mainly driven by features of its immediate habitat. This may reflect the species’ opportunistic behaviour or may be due to its small ranging ability60,62. Based on our results, mink was more likely to use rafts established along riverbanks and lake shores with more shrubs and rocks. These patterns have been previously described in several other studies22,23,25,26,27,62,63 and may reflect the species’ requirements in nesting and in avoiding predation or competition24,25,64,65,66. Yamaguchi et al.25 and Schüttler et al.26 found that the species avoids exposed areas and mainly uses areas with dense vegetation, which provide more cover and nesting opportunities57,64. In addition, the species may favour such areas because they offer more feeding opportunities27. According to Torre and Diaz67 and Torre et al.68, in Mediterranean ecosystems, shrubs support a greater diversity and density of small mammals, many of which may serve as prey for the mink. Similarly, rocky areas can be used for denning when the availability of other nesting types is limited63 or to avoid predation63,69. According to Brainerd et al.70, Zalewski71, Achterberg et al.72 and Stier73, mustelids usually favour tree cavities for nesting. However, when availability is low, the species may seek alternative sites, such as areas with rocks or boulders63. Elevated rock cavities resemble trees, as both are inaccessible to terrestrial predators69, whereas dens located on the ground between rocks provide protection against predators with digging abilities63. Hence, predation risk is likely to be a significant factor influencing the choice of cover and den sites for the species25,57,74,75. Because many of the minks’ natural predators and competitors can be found across our study area, individuals may be forced to select densely vegetated or rocky riparian areas6,25,57,74.
In addition, the probability of initial raft use by the species was found to be positively associated with the density of medium-sized rivers at the landscape scale. Zabala et al.76 reported that mustelids show a preference for secondary rivers with sufficient riparian vegetation cover, while they avoid areas with polluted waters or modified riverbeds. Moreover, Sidorovich and Macdonald18 reported that mustelids avoid large, fast-flowing rivers, while Garcia et al.27 concluded that because secondary rivers are characterized by the existence of islets within the watershed, these may serve as resting sites for the species25,74. Our study area does not include large rivers (Stahler order > 6); therefore, the positive association between mink raft use and medium-sized rivers may be because this river category is characterized by continuous water flow throughout the year, in contrast to smaller rivers in the area, which are characterized by seasonal flow and shallow water depth.
The probability of a vacant raft being colonized (used) between surveys was found to be positively associated with the percent cover of shrubs and reeds. However, none of these variables were informative. Previous studies have shown that colonization may be affected by stream size, water depth and flow regime55,77,78. In addition, colonization may be closely related to habitat features that increase denning and resting site availability78 or may be closely related to prey distribution and urbanization79. Conversely, the distance to the nearest fur farm within our study area was very informative in predicting the probability of raft abandonment by the species. Moreover, almost 70% of all mink presence sites were within a 5 km radius from nearby fur farms, whereas 90% of all individuals captured were also within a 5 km radius from nearby farms. According to Bonesi and Palazón7, fur farms are considered the main source of American mink reintroduction. With that in mind and given that mink are very elusive species80, eradication campaigns are often unsuccessful. The number of minks escaping from fur farms located within the study area is unknown; however, reducing the number of individuals introduced into the wild may help control the successful establishment and spread of American minks in the wider region81. Nonetheless, both wild and introduced individuals illustrate similar feeding habits and exhibit the same ability to hunt their prey82. In addition, individuals introduced from fur farms may act as a source of increasing genetic diversity and adaptation for wild populations83.
Conclusions
Knowledge of the environmental parameters that influence the occupancy dynamics of invasive species is fundamental in planning and implementing effective management actions84. Management options to mitigate the negative impacts caused by American mink introduction and establishment may include direct population control (e.g., removal trapping) or habitat modification8. However, managing the species at a small spatial scale is likely to be more practical and effective than managing it at the landscape level57,85. In addition, management actions should consider identifying target areas, both for efficiency and economy but also for protecting biodiversity8. According to our results, management actions such as trapping American mink individuals could be more effective if they are focused in areas within a 5 km radius from the nearest fur farms and along rocky or densely vegetated riverbanks or lake shores. In addition, trapping efforts during autumn are likely to be more effective since during this time, the species has a high detection probability. On the other hand, mink control may involve habitat management by reducing the suitability of habitats used by the species for nesting and hiding and improving the suitability of habitats for other competing species, such as the otter (Lutra lutra). However, in this case, and due to the opportunistic behaviour and the species’ ability to adapt to various conditions and environments, such actions may be more difficult to implement8. While our study focuses on mink raft use rather than occupancy, we believe that our findings are of great importance to managers, since floating rafts are among the main methods used to record, monitor, and control the American mink. Future studies should focus on investigating the factors that may affect the species’ ability to colonize new areas, such as water quality78 and prey distribution29, which were not addressed in the current study but have been shown to be of great importance. Finally, the composition of free-ranging individuals should be investigated in future studies in order to distinguish between wild-born and farm-born individuals. In the latter case, and if most individuals are a product of recent escapes, management measures should focus on maximizing fur farm security and establishing an early warning and rapid eradication system.
Data availability
The datasets analysed during the current study are available from the corresponding author (D.E.B.) upon reasonable request.
References
Hulme, P. E., Pyšek, P., Nentwig, W. & Vilà, M. Will threat of biological invasions unite the European Union? Science 1979(324), 40–41 (2009).
Tittensor, D. P. et al. A mid-term analysis of progress toward international biodiversity targets. Science 1979(346), 241–244 (2014).
Blackburn, T. M., Dyer, E., Su, S. & Cassey, P. Long after the event, or four things we (should) know about bird invasions. J. Ornithol. 156, 15–25 (2015).
Vitousek, P. M., D’antonio, C. M., Loope, L. L., Rejmanek, M. & Westbrooks, R. Introduced species: A significant component of human-caused global change. N. Z. J. Ecol. 21(1), 1–16 (1997).
Sala, O. E. et al. Global biodiversity scenarios for the year 2100. Science 1979(287), 1770–1774 (2000).
Larivière, S. Mustela vison. Mamm. Spec. https://doi.org/10.2307/3504420 (1999).
Bonesi, L. & Palazon, S. The American mink in Europe: Status, impacts, and control. Biol. Conserv. 134, 470–483 (2007).
Macdonald, D. W. & Harrington, L. A. The American mink: The triumph and tragedy of adaptation out of context. N. Z. J. Zool. 30, 421–441 (2003).
Semos, N., Dotas, V. & Bampidis, V. Development strategies for the fur farming industry in Greece. Open J. Bus. Manag. 9, 2683–2696 (2021).
Henriksen, B. I. F., Møller, S. H. & Malmkvist, J. Animal welfare measured at mink farms in Europe. Appl. Anim. Behav. Sci. 248, 105587 (2022).
Rushton, S. P., Barreto, G. W., Cormack, R. M., Macdonald, D. W. & Fuller, R. Modelling the effects of mink and habitat fragmentation on the water vole. J. Appl. Ecol. 37, 475–490 (2000).
Põdra, M., Gómez, A. & Palazón, S. Do American mink kill European mink? Cautionary message for future recovery efforts. Eur. J. Wildl. Res. 59, 431–440 (2013).
Põdra, M. & Gómez, A. Rapid expansion of the American mink poses a serious threat to the European mink in Spain. Mammalia 82, 580–588 (2018).
Mori, E. & Mazza, G. Diet of a semiaquatic invasive mammal in northern Italy: Could it be an alarming threat to the endemic water vole?. Mamm. Biol. 97, 88–94 (2019).
Brzeziński, M., Żmihorski, M., Nieoczym, M., Wilniewczyc, P. & Zalewski, A. The expansion wave of an invasive predator leaves declining waterbird populations behind. Divers. Distrib. 26, 138–150 (2020).
Ferreras, P. & Macdonald, D. W. The impact of American mink Mustela vison on water birds in the upper Thames. J. Appl. Ecol. 36, 701–708 (1999).
Hammershøj, M., Thomsen, E. A. & Madsen, A. B. Diet of free-ranging American mink and European polecat in Denmark. Acta Theriol. (Warsz) 49, 337–347 (2004).
Sidorovich, V. & Macdonald, D. W. Density dynamics and changes in habitat use by the European mink and other native mustelids in connection with the American mink expansion in Belarus. Neth. J. Zool. 51, 107–126 (2001).
Manas, S. et al. Aleutian mink disease parvovirus in wild riparian carnivores in Spain. J. Wildl. Dis. 37, 138–144 (2001).
Vada, R. et al. Feral American mink Neogale vison continues to expand its European range: Time to harmonise population monitoring and coordinate control. Mamm. Rev. 53, 158–176 (2023).
Crego, R. D., Jimenez, J. E. & Rozzi, R. Potential niche expansion of the American mink invading a remote island free of native-predatory mammals. PLoS One 13, e0194745 (2018).
Allen, A. W. Western Energy and Land Use Team, Division of Biological Services, Research and Development, Fish and Wildlife Service, US Department of the Interior. Habitat Suitability Index Models: Mink (1984).
Previtali, A., Cassini, M. H. & Macdonald, D. W. Habitat use and diet of the American mink (Mustela vison) in Argentinian Patagonia. J Zool 246, 443–486 (1998).
Bonesi, L., Dunstone, N. & O’Connell, M. Winter selection of habitats within intertidal foraging areas by mink (Mustela vison). J. Zool. 250, 419–424 (2000).
Yamaguchi, N., Rushton, S. & Macdonald, D. W. Habitat preferences of feral American mink in the Upper Thames. J. Mammal. 84, 1356–1373 (2003).
Schüttler, E., Ibarra, J. T., Gruber, B., Rozzi, R. & Jax, K. Abundance and habitat preferences of the southernmost population of mink: Implications for managing a recent island invasion. Biodivers. Conserv. 19, 725–743 (2010).
García, P., Arévalo, V. & Lizana, M. Characterisation of den sites of American mink Neovison vison in central Spain. Wildl. Biol. 16, 276–282 (2010).
Carlsson, N. O. L., Jeschke, J. M., Holmqvist, N. & Kindberg, J. Long-term data on invaders: When the fox is away, the mink will play. Biol. Invasions 12, 633–641 (2010).
Wolff, P. J., Taylor, C. A., Heske, E. J. & Schooley, R. L. Habitat selection by American mink during summer is related to hotspots of crayfish prey. Wildl. Biol. 21, 9–17 (2015).
Hodder, D. P., Larsen, K. W. & Crowley, S. M. The role of environmental variables and sympatric meso-carnivores on the detection and occupancy of American mink during winter. Hystrix 28, 16–20 (2017).
Galanaki, A. & Kominos, T. The distribution of American mink (Neovison vison) in Greece. Mammalia 86, 57–65 (2022).
Ministry of Environment—Special Water Secretariat. River Basin Management Plan of Western Macedonia (2014).
Ministry of Environment—Special Water Secretariat. Special Management Plan for the Sub-basin of Prespa in the River Basin of Prespa (GR01) (2014).
Reynolds, J. C., Short, M. J. & Leigh, R. J. Development of population control strategies for mink Mustela vison, using floating rafts as monitors and trap sites. Biol. Conserv. 120, 533–543 (2004).
Porteus, T., Short, M., Richardson, S. & Reynolds, J. Empirical development of strategy for the control of invasive American mink by trapping. Eur. J. Wildl. Res. 58, 403–413 (2012).
Reynolds, J. C., Richardson, S. M., Rodgers, B. J. E. & Rodgers, O. R. K. Effective control of non-native American mink by strategic trapping in a river catchment in mainland Britain. J. Wildl. Manage. 77, 545–554 (2013).
Melero, Y., Palazón, S., Revilla, E., Martelo, J. & Gosálbez, J. Space use and habitat preferences of the invasive American mink (Mustela vison) in a Mediterranean area. Eur. J. Wildl. Res. 54, 609–617 (2008).
ESRI. Desktop Release 10 (Environmental Systems Research Institute, 2011).
MacKenzie, D. I., Nichols, J. D., Hines, J. E., Knutson, M. G. & Franklin, A. B. Estimating site occupancy, colonization, and local extinction when a species is detected imperfectly. Ecology 84, 2200–2207 (2003).
Rota, C. T., Fletcher, R. J. Jr., Dorazio, R. M. & Betts, M. G. Occupancy estimation and the closure assumption. J. Appl. Ecol. 46, 1173–1181 (2009).
Dagtekin, D., Ertürk, A., Sommer, S., Ozgul, A. & Soyumert, A. Seasonal habitat-use patterns of large mammals in a human-dominated landscape. J. Mammal. 105, 122–133 (2024).
Louvrier, J. et al. Mapping and explaining wolf recolonization in France using dynamic occupancy models and opportunistic data. Ecography 41, 647–660 (2018).
Roda, F. et al. Wolf scat detection dog improves wolf genetic monitoring in new French colonized areas. J. Vertebr. Biol. 69, 20101–20102 (2021).
Arnold, T. W. Uninformative parameters and model selection using Akaike’s Information Criterion. J. Wildl. Manage. 74, 1175–1178 (2010).
Ramírez-Cruz, G. A. & Ortega-Álvarez, R. Identifying management guidelines to control the invasive House Sparrow (Passer domesticus) within natural protected areas through the estimation of local colonization and extinction probabilities. Biol. Invasions 23, 3767–3776 (2021).
Fiske, I. & Chandler, R. Unmarked: An R package for fitting hierarchical models of wildlife occurrence and abundance. J. Stat. Softw. 43, 1–23 (2011).
Kéry, M. & Chandler, R. Dynamic occupancy models in unmarked (2012). http://cran.r-project.org/web/packages/unmarked/vignettes/colext.pdf Accessed 20 April 2015
R Core Team, R. R: A language and environment for statistical computing (2013).
Catford, J. A., Jansson, R. & Nilsson, C. Reducing redundancy in invasion ecology by integrating hypotheses into a single theoretical framework. Divers. Distrib. 15, 22–40 (2009).
Bled, F., Royle, J. A. & Cam, E. Hierarchical modeling of an invasive spread: The Eurasian collared-dove Streptopelia decaocto in the United States. Ecol. Appl. 21, 290–302 (2011).
Yackulic, C. B. et al. Neighborhood and habitat effects on vital rates: Expansion of the Barred owl in the Oregon coast ranges. Ecology 93, 1953–1966 (2012).
Santulli, G., Palazón, S., Melero, Y., Gosálbez, J. & Lambin, X. Multi-season occupancy analysis reveals large scale competitive exclusion of the critically endangered European mink by the invasive non-native American mink in Spain. Biol. Conserv. 176, 21–29 (2014).
Harrington, L. A., Harrington, A. L. & Macdonald, D. W. Estimating the relative abundance of American mink Mustela vison on lowland rivers: evaluation and comparison of two techniques. Eur. J. Wildl. Res. 54, 79–87 (2008).
Bryce, R. et al. Turning back the tide of American mink invasion at an unprecedented scale through community participation and adaptive management. Biol. Conserv. 144, 575–583 (2011).
Schooley, R. L., Cotner, L. A., Ahlers, A. A., Heske, E. J. & Levengood, J. M. Monitoring site occupancy for American mink in its native range. J. Wildl. Manage. 76, 824–831 (2012).
Reynolds, J. C., Porteus, T. A., Richardson, S. M., Leigh, R. J. & Short, M. J. Detectability of American mink using rafts to solicit field signs in a population control context. J. Wildl. Manage. 74, 1601–1606 (2010).
Dunstone, N. The Mink (Poyser, 1993).
Yamaguchi, N. & Macdonald, D. W. The burden of co-occupancy: Intraspecific resource competition and spacing patterns in American mink, Mustela vison. J. Mammal. 84, 1341–1355 (2003).
Roy, S., Reid, N. & McDonald, R. A. A review of mink predation and control in Ireland. in Irish Wildlife Manuals vol. 40 (National Parks and Wildlife Service, Department of the Environment, Heritage and Local Government, 2009).
Dunstone, N. & Birks, J. D. S. The comparative ecology of coastal, riverine and lacustrine mink Mustela vison in Britain. Zeitschrift für Angew. Zool. 72, 59–70 (1985).
Bonesi, L. & Macdonald, D. W. Evaluation of sign surveys as a way to estimate the relative abundance of American mink (Mustela vison). J. Zool. 262, 65–72 (2004).
Lundy, M. G. & Montgomery, W. I. A multi-scale analysis of the habitat associations of European otter and American mink and the implications for farm scale conservation schemes. Biodivers. Conserv. 19, 3849–3859 (2010).
Birks, J. D. S., Messenger, J. E. & Halliwell, E. C. Diversity of den sites used by pine martens Martes martes: A response to the scarcity of arboreal cavities?. Mamm. Rev. 35, 313–320 (2005).
Halliwell, E. C. & Macdonald, D. W. American mink Mustela vison in the Upper Thames catchment: Relationship with selected prey species and den availability. Biol. Conserv. 76, 51–56 (1996).
McDonald, R. A. Resource partitioning among British and Irish mustelids. J. Anim. Ecol. 71, 185–200 (2002).
Bonesi, L., Harrington, L. A., Maran, T., Sidorovich, V. E. & Macdonald, D. W. Demography of three populations of American mink Mustela vison in Europe. Mamm. Rev. 36, 98–106 (2006).
Torre, I. & Díaz, M. Small mammal abundance in Mediterranean post-fire habitats: A role for predators?. Acta Oecol. 25, 137–142 (2004).
Torre, I., Jaime-González, C. & Díaz, M. Habitat suitability for small mammals in Mediterranean landscapes: How and why shrubs matter. Sustainability 14, 1562 (2022).
Webster, J. A. A review of the historical evidence of the habitat of the pine marten in Cumbria. Mamm. Rev. 31, 17–31 (2001).
Brainerd, S. M. et al. Pine marten (Martes martes) selection of resting and denning sites in Scandinavian managed forests. Ann. Zool. Fennici. 32, 151–157 (1995).
Zalewski, A. Factors affecting selection of resting site type by pine marten in primeval deciduous forests (Bialowieza National Park, Poland). Acta Theriol. (Warsz) 42, 271–288 (1997).
Achterberg, C., Bestman, M. & Wijsman, H. J. W. Inventarisatie van boommarternestbomen op de Utrechtse Heuvelrug 1992–1999. Lutra 43, 93–100 (2000).
Stier, N. Tagesverstecke des Baummarders (Martes martes L.) in Südwest-Mecklenburg. Beiträge zur Jagd-und Wildforschung 25, 165–182 (2000).
Zabala, J., Zuberogoitia, I. & Martínez-Climent, J. A. Winter habitat preferences of feral American mink Mustela vison in Biscay, Northern Iberian Peninsula. Acta Theriol. (Warsz) 52, 27–36 (2007).
Harrington, L. A. & Macdonald, D. W. Spatial and temporal relationships between invasive American mink and native European polecats in the southern United Kingdom. J. Mammal. 89, 991–1000 (2008).
Zabala, J., Zuberogoitia, I. & Martínez-Climent, J. A. Factors affecting occupancy by the European mink in south-western Europe/Facteurs affectant l’habitat du vison en Europe sud-occidentale. Mammalia 70, 193–201 (2006).
Ahlers, A. A. et al. Summer precipitation predicts spatial distributions of semiaquatic mammals. PLoS One 10, e0135036 (2015).
Holland, A. M., Schauber, E. M., Nielsen, C. K. & Hellgren, E. C. River otter and mink occupancy dynamics in riparian systems. J. Wildl. Manage. 83, 1552–1564 (2019).
Ahlers, A. A., Heske, E. J. & Schooley, R. L. Prey distribution, potential landscape supplementation, and urbanization affect occupancy dynamics of American mink in streams. Landsc. Ecol. 31, 1601–1613 (2016).
Medina-Vogel, G., Barros, M., Monsalve, R. & Pons, D. J. Assessment of the efficiency in trapping North American mink (Neovison vison) for population control in Patagonia. Rev. Chile. Historia Nat. 88, 1–12 (2015).
Jeschke, J. M. & Strayer, D. L. Determinants of vertebrate invasion success in Europe and North America. Glob. Change Biol. 12, 1608–1619 (2006).
Rørbæk, R. W., Andersen, T. A., Pertoldi, C., Jørgensen, A. & Pagh, S. Diet of free ranging American mink (Neovison vison) in Denmark. Animals 13, 461 (2023).
Pagh, S. et al. Methods for the identification of farm escapees in feral mink (Neovison vison) populations. PLoS One 14, e0224559 (2019).
MacKenzie, D. I. et al. Occupancy Estimation and Modeling: Inferring Patterns and Dynamics of Species Occurrence (Elsevier, 2017).
Boitani, L. Carnivore introductions and invasions: Their success and management. In Carnivore Conservation, 123–144 (1999).
Acknowledgements
This study was part of the LIFE “ATIAS” project and has received funding from the European Union’s LIFE programme (agreement number: LIFE18 NAT/GR/000430), with the contribution of the Hellenic Green Fund (181.1/2020). We are very grateful to many people from the Forest Services of the Decentralized Administration of Epirus-West Macedonia and the Decentralized Administration of Macedonia and Thrace, the Hunting Federation of Macedonia and Thrace, and Homeotech who helped with the fieldwork during the various parts of this study. Authors would also like to thank two anonymous reviewers for their insightful suggestions, which helped improve the manuscript.
Author information
Authors and Affiliations
Contributions
D.E.B. conceptualization, funding acquisition, writing—review, and editing. C.T.T. and K.N.M. data curation, data analyses, visualization, writing—original draft. E.G.K. writing—review and editing. All authors critically reviewed the manuscript and approved the final version of the manuscript for publication.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bakaloudis, D.E., Thoma, C.T., Makridou, K.N. et al. Occupancy dynamics of free ranging American mink (Neogale vison) in Greece. Sci Rep 14, 9973 (2024). https://doi.org/10.1038/s41598-024-60542-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-60542-4
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
