
Abstract
Although giant fennel is recognized as a “superfood” rich in phytochemicals with antioxidant activity, research into the antibacterial properties of its fruits has been relatively limited, compared to studies involving the root and aerial parts of the plant. In this study, seven solvents—acetone, methanol, ethanol, ethyl acetate, chloroform, water, and hexane—were used to extract the chemical constituents of the fruit of giant fennel (Ferula communis), a species of flowering plant in the carrot family Apiaceae. Specific attributes of these extracts were investigated using in silico simulations and in vitro bioassays. High-performance liquid chromatography equipped with a diode-array detector (HPLC–DAD) identified 15 compounds in giant fennel extract, with p-coumaric acid, 3-hydroxybenzoic acid, sinapic acid, and syringic acid being dominant. Among the solvents tested, ethanol demonstrated superior antioxidant activity and phenolic and flavonoid contents. F. communis extracts showed advanced inhibition of gram-negative pathogens (Escherichia coli and Proteus mirabilis) and variable antifungal activity against tested strains. Molecular docking simulations assessed the antioxidative, antibacterial, and antifungal properties of F. communis, facilitating innovative therapeutic development through predicted compound–protein interactions. In conclusion, the results validate the ethnomedicinal use and potential of F. communis. This highlights its significance in natural product research and ethnopharmacology.
Similar content being viewed by others


Introduction
The genus Ferula belongs to the Apiaceae family of perennial herbaceous plants1. It includes approximately 150 species well-suited to diverse Mediterranean and continental climates and grow spontaneously in Morocco2. With a documented history of medicinal use, its pharmacological effects are well-established in both human and veterinary practices3. Six Ferula species have been observed in Morocco: Ferula communis, Ferula cossoniana, Ferula gouliminensis, Ferula sauvagei, Ferula atlantica, and Ferula tingitana4. Known as giant fennel (Ferula communis L.) (la grande Ferule in French and l-bubāl in Arabic) is referred to as ūffal, tuffalt in the Berbère language, where its fruit is sold as a vegetable in Moroccan marketplaces (souks)5. F. communis is renowned for its gum resin (l-fāsūẖ)6, harvested from the rootstock after stripping and incision7. The fruit of F. communis is primarily used as a vegetable, prepared by steaming or ashing, and subsequently chopped and seasoned with olive oil, salt, and pepper8. F. communis grazing is generally avoided by herds, as it can cause poisoning, except during droughts.
This genuine variety is characterized by the synthesis of numerous hydroxycoumarins, including ferulenol9. Recent studies highlight the potential of isolated compounds from this genus as chemopreventive agents and in overcoming resistance to anticancer drugs8,10. Traditionally, plants of this genus have been used to treat various diseases9. Moreover, they exhibit anticonvulsant, stimulant, and expectorant properties11. The genus Ferula is categorized based on the prevalence of oleo-gum resins (Asafoetida) and their roles in natural and conventional medications12. Asafoetida has been used as a tranquilizer to decrease blood pressure. It is commonly used in Indian food and traditional medicine, particularly within the framework of Indian medical structures such as Ayurveda8. Asafoetida, a spice with centuries of medicinal use, is employed for therapeutic use. In the Middle Atlas, unexpanded inflorescences of Ferula serve as anthelmintics9. Polyphenols have well-defined antioxidant properties linked to the inhibition of food oxidation10. In addition, these antioxidants effectively block or inhibit the formation and propagation of free radicals13. Therefore, they are of interest in several fields, including nutrition, cosmetology, and the food industry14,15,16. The antioxidant activities of polyphenols can be evaluated in vitro and in vivo using simple experiments17.
Solvent extraction is useful for the recovery of antioxidant compounds from plant materials. Extraction yields and antioxidant activities vary depending on the extraction method and solvents used18. Polar solvents have gained prominence in the recovery of polyphenols from plant matrices19. In particular, aqueous solvent mixtures containing ethanol, methanol, and acetone are effective. Ethanol has gained recognition as a suitable solvent for extracting polyphenols and is safe for human consumption11. Conversely, methanol is efficient in extracting low-molecular-weight polyphenols20, whereas aqueous acetone has been identified as more suitable for the extraction of greater-molecular-weight flavanols12.
Despite the well-established historical uses of F. communis, further scientific studies are needed to explore the activities of its extracts. To the best of our knowledge, comprehensive investigations on the biological activity of wild Moroccan F. communis fruits are yet to be published. This highlights the novelty and significance of our research, addressing critical knowledge gaps by examining the biochemical constituents of F. communis and investigating the effects of various solvents (methanol, ethanol, water, acetone, hexane, ethyl acetate, and chloroform) on antioxidant activity. The antioxidant, antibacterial, and antifungal activities of wild Moroccan F. communis L. were evaluated against multidrug-resistant pathogenic strains.
Methods
Plant material
Ferula communis was harvested at the end of January from Sefrou Province, Morocco (33.6957°N, 4.3716°W) and subsequently identified by Professor Bari Amina, a botanist in the Department of Biology, Faculty of Sciences Dhar El Mehraz, Sidi Mohamed Ben Abdellah University, where a voucher specimen (FC0522) was deposited at the herbarium. Notably, no approval was required to collect F. communis from Morocco for research purposes. The fruits used in this study were dried for 15 d in ambient air in the laboratory.
Extract preparation
The dried fruits (2 g) of F. communis L. were powdered using an electric mixer (Blender Waring®) and mixed with 20 mL of solvent. Various solvents including methanol, ethanol, water, hexane, acetone, ethyl acetate, and chloroform were used individually. Macerates were filtered through Whatman No. 1 filter paper (Merck KGaA, Darmstadt, Allemagne), and the obtained extracts were stored in sterile sample tubes at 4 °C.
High-performance liquid chromatography with diode-array detection analysis
Extracts of F. communis were prepared at concentrations of 50 mg/mL and filtered through 0.45 μm microfilters. Phenolic compounds were characterized using high-performance liquid chromatography (HPLC) connected to a UV detector (210–400 nm). Notably, 20 μL of extract was injected through a (C18) reverse phase column (250 × 4.6 mm, 5 μm) with an elution gradient of A (water/0.5% phosphoric acid) and B (methanol) at a flow rate of 1 mL/min. Compounds were identified by comparing retention times with authentic standards. Quantification was performed by comparison with external calibration curves and expressed as mg extract equivalents/100 g of dry matter. Analyses were performed in triplicate, and all calibration curves showed good linearity (R2 > 0.99).
Antioxidant activity
2,2-Diphenyl-1-picrylhydrazyl free radical scavenging activity
The total free radical scavenging capacity of various extracts of F. communis fruit was estimated according to a previously reported method14. Extracts (50 µL) at concentrations ranging from 12.5 to 50 mg/mL were added to 825 μL of ethanolic 2,2-diphenyl-1-picrylhydrazyl (DPPH). The mixture was shaken vigorously and maintained at room temperature in the dark for 1 h. The absorbance of the DPPH radical only served as the blank. All measurements were performed in triplicate. The ability to scavenge DPPH radicals was calculated using Eq. (1):
2,2-Azinobis (3-ethyl-benzothiazoline-6-sulfonic acid) radical scavenging activity
Free radical scavenging activity of wild F. communis fruit extracts were determined using an 2,2ʹ-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid (ABTS) free radical cation decolorization assay15. Briefly, 50 μL of various dilutions of each ethanolic extract or gallic acid (used as a positive control) were added to 825 μL of ABTS radical cation solution. Thereafter, the solutions were incubated at room temperature in the dark for 6 min. Absorption was measured at 734 nm using a UV/Vis spectrophotometer. Absorption of a blank sample containing the same amounts of ethanol and ABTS solution served as the negative control. The inhibition (%) of absorbance was calculated using Eq. (1), and IC50 values were determined graphically and expressed as mg ATBS equivalents/mL. All analyses were performed in triplicate.
Reducing power
Reducing power (RP) was determined by combining 50 μL of each F. communis fruit extract with 200 μL of 0.2 M sodium phosphate buffer (pH 7.6) and 200 μL of 1% potassium ferricyanide16. The mixture was stored at 50 °C for 20 min. Two hundred microliters of 10% trichloroacetic acid was added, and the mixture was centrifuged at 2.217×g for 10 min. The supernatant was mixed with 600 μL of distilled water and 120 μL of 0.1% ferric chloride. Ascorbic acid (0.073–1.238 mg/mL) was used as a standard to obtain a calibration curve (R2 = 0.9996). A graph was plotted using the absorbance at 700 nm versus sample concentration. RP is defined as the extract concentration yielding 0.5% absorbance (EC50) and expressed in mg/mL. All measurements were performed in triplicate.
Total antioxidant capacity
The total antioxidant capacity (TAC) of extracts were evaluated using the phosphomolybdenum method17. Briefly, 50 µL of various extracts was mixed with 1 mL of reagent solution (sulfuric acid 0.6 M, 28 mM sodium phosphate, and 4 mM ammonium molybdate). The absorbance of the sample was recorded against a blank at 695 nm after 90 min of incubation in a water bath at 95 °C using a Perkin Elmer Lambda 40 UV/Vis spectrophotometer. Results are expressed in mg of ascorbic acid equivalents (AAE) per g of dry plant (mg AAE/g DW), as the mean of three triplicate ± standard deviation (SD).
Total polyphenols and flavonoids in F. communis
Total phenolic content
The total phenolic content (TPC) was determined using a colorimetric method based on the Folin–Ciocalteu reagent18. First, 50 μL of gallic acid extract or standard solution was mixed with 450 μL of Folin–Ciocalteu reagent solution (10%), shaken on a vortex, and incubated for 5 min. Thereafter, 450 μL of a Na2CO3 solution (75 g L−1) was added, and the mixture was shaken again. Following a 2 h incubation at room temperature (25 + 1 °C), absorption was measured at 760 nm using a spectrophotometer. The gallic acid calibration curve displayed high linearity with a standard curve equation (R2 = 0.9994). The results are expressed in mg gallic acid equivalent (GAE)/g DW. All analyses were performed in triplicate.
Flavone/flavanol content
An aliquant of 500 μL of extracts or quercetin, used as the positive control, was mixed with 500 μL of AlCl3 (2%). After incubation for 1 h at ambient temperature, the absorbance of the mixture was measured at 420 nm using a UV/Vis spectrophotometer. Quercetin (0.003–0.5 mg/L) was used to obtain a standard curve (R2 = 0.9998), and the results were expressed as milligrams of quercetin equivalent (QE) per gram of sample (mg QE/g). All tests were performed in triplicate19.
Antimicrobial activity of the ethanol, acetone, and water extracts of F. communis
Microbial strains
The antimicrobial activities of F. communis extracts in ethanol (EtOH), acetone (AcE), or water (AqE) were determined against four fungal strains, including Candida albicans (ATCC 10231), Aspergillus niger (MTCC 282), Aspergillus flavus (MTCC 9606), and Fusarium oxysporum (MTCC 9913), and four bacterial strains, including Staphylococcus aureus (ATCC 6633), Escherichia coli (K12), Bacillus subtilis (DSM 6333), and Proteus mirabilis (ATCC 29906). All microbial strains were provided by the Laboratory of Biotechnology, Environment, Agri-Food, and Health, Faculty of Sciences, Dhar El Mahraz, Sidi Mohammed Ben Abdellah University, Fez, Morocco.
Assessment of antimicrobial activity
The antimicrobial activities of the aqueous, ethanol, and acetone extracts of F. communis were evaluated using the disk diffusion method11. Briefly, Petri dishes containing Mueller–Hinton (MH) culture media and malt extract (ME) were individually inoculated with each of the four bacterial strains in combination with C. albicans, using the double layer method. Ten-fold dilutions were prepared in sterile saline (0.9% NaCl) from fresh cultures grown in MH and ME media, reaching a turbidity of 0.5 McFarland (106–108 CFU/mL). Thereafter, 100 µL was added to tubes containing 5 mL soft agar (0.5% agar), and the inoculated tubes were placed into Petri dishes containing MH and EM media. Whatman no. 4 filter paper discs (6 mm in diameter) were impregnated with 20 μL of the AqE, EtOH, and AcE extracts of F. communis. For A. niger, A. flavus, and F. oxysporum, the antifungal activity was evaluated by confrontation in ME medium. To assess the efficacy of the negative and positive controls, F. communis extracts containing the conventional antibiotics, oxacillin for bacterial strains and fluconazole for fungal pathogens, were tested in a similar manner. Petri dishes inoculated with bacteria and fungi were incubated at 37 °C and 30 °C, respectively20. The diameters of inhibition and inhibition percentages were calculated 24 h post-inoculation (hpi) for the bacterial strains, 48 hpi for C. Albicans, and 7 d post-inoculation for F. oxysporum, A. niger, and A. flavus12,21.
Minimum inhibitory concentration
The minimum inhibitory concentration (MIC) of the AqE, EtOH, and AcE extracts of F. communis against the four bacterial and four fungal pathogens were determined using the microdilution method described by Sarker et al.22. Microplates were prepared under aseptic conditions, and each sterile 96-well microplate was labeled. Thereafter, 100 µL of EtOH, AcE, or AqE extracts of F. communis in 2% DMSO was pipetted into the first column of the plate. All other wells contained 50 µL sterile MH for bacterial pathogens and 50 µL of sterile EM for fungal pathogens. Serial dilutions were made using a multichannel pipette; finally, 40 µL of microbial suspension of each strain (106 to 108 CFU/mL) was added to each well. After a 24-h incubation period for bacteria, 48-h period for C. albicans, and 7-d period for A. niger, A. flavus, and F. oxysporum, the MIC endpoint was quantified calorimetrically (TTC, 0.2% (w/v))23.
Molecular docking
To elucidate the antioxidant, antibacterial, and antifungal effects of F. communis, major compounds in their extracts, including syringic acid (C7), 3-hydroxybenzoic acid (C8), and p-coumaric acid (C12), were docked to the active sites of NADPH oxidase, FimH lectin, and CYP51 proteins encoded in the Protein Data Bank (PDB) using 2CDU, 4XO8, and 5TZ1, respectively. Molecular docking was performed via the following steps: (1) Three-dimensional structures of targeted proteins were extracted from the Research Collaboratory for Structural Bioinformatics (RCSB) PDB basis (https://www.rcsb.org/); (2) Proteins in PDB files were prepared using Discovery Studio 2021 software (https://www.3ds.com/products-services/biovia/products/molecular-modeling-simulation/biovia-discovery-studio/)24,25, removing all co-crystallized ligands and water molecules, and adding the charges of Gasteiger; (3) The examined ligands (C7, C8, and C12) and the prepared proteins were converted from PDB to PDBQT files using Autodock software version 4.2 (https://doi.org/10.1186/1758-2946-3-12)26; (4) Grid boxes were centralized on each targeted protein, maximizing dimensions in three directions (OX, OY, and OZ) with a spacing of 0.375 Å, as shown in Table 1. The produced complexes were visualized using Discovery Studi 2021 software27.
Statistical analyses
All results are expressed as the mean of triplicate experiments ± SD. The significance of the difference between means was tested using a two-way analysis of variance (ANOVA). Tukey’s multiple range tests (p < 0.05) were performed using GraphPad Prism 8.0.1 (Graph Pad Software Inc., San Diego, United States), and correlations were added using Origin Pro-2023.
Plant collection approval
No approval is needed to collect Ferula communis in Morocco for research purposes.
IUCN policy statement
The collection of plant material complies with relevant institutional, national, and international guidelines and legislation.
Results and discussion
Extraction yield
The water/raw material ratio is a critical factor affecting extraction yield of active components from plant materials. Furthermore, the recovery of phenolic and flavonoid compounds is considerably influenced by polarity28. The purpose of the experiments in this study was to assess the effect of solvents on the extraction yield.
The results presented in Table 2 indicate that polar protic solvents had better extraction yields. Among all solvents used, ethanol and methanol achieved significantly higher extraction yields, with rates of 15.25 ± 1.0% and 15 ± 1.2%, respectively. This suggested that ethanol and methanol are optimal solvents for maximizing the extraction yield of F. communis fruit, followed by water, acetone, and ethyl acetate with yields of 7.9 ± 0.25%, 2.6 ± 0.05%, and 2.3 ± 0.02%, respectively. However, hexane and chloroform achieved the lowest yields, with rates of 0.9 ± 0.012% and 0.6 ± 0.03%, respectively.
The polarity of the ethanol system is highly stable for the extraction of flavonoids and betacyanin glycosides, such as isorhamnetin-3-O-rutinoside, betanin, isobetanin, and betanidine, and polar components, such as phospholipids, polysaccharides, and lipoproteins20. Acetone and water are common extractants of proanthocyanidins and tannins12.
Our experiments show that the selection of an appropriate extraction solvent depends on the type and polarity of the constituents in the plant material.
HPLC analysis
Fifteen phenolic compounds, including gallic acid, caffeic acid, catechin, 4-hydroxybenzoic acid, catechin hydrate, succinic acid, syringic acid, 3-hydroxybenzoic acid, naringin, cinnamic acid, ferulic acid, p-coumaric acid, sinapic acid, quercetin 3-O-β-d-glucoside, and rutin (Fig. 1), were identified and quantified using HPLC–DAD. In a previous study, various phenolic compounds, such as resorcinol, ferulic acid, syringic acid, and coumarin, are identified as the predominant components in F. communis fruits from various geographical origins. Compounds were identified according to relative retention times based on reference standards and reported as percentage areas (Table 3)7.

HPLC–DAD chromatogram of F. communis extract fruits (FC-Ext) at 320 nm using the following standards: Gallic acid (1), Caffeic acid (2), Catechin (3), 4-Hydroxybenzoic acid (4), Catechin hydrate (5), Succinic acid (6), Syringic acid (7), 3-hydroxybenzoic acid (8), Naringin (9), Cinnamic acid (10), Ferulic acid (11), p-Coumaric acid (12), Sinapic acid (13), Quercetin 3-O-β-d-glucoside (14), Rutin (15).
In a study focusing on F. communis rhizomes and employing a bioautography-guided isolation technique, three antibacterial sesquiterpenes were successfully isolated and characterized. These compounds were identified as daucane 14-(O-hydroxycinnamoyloxy)-dauc-4,8-diene, 2a-acetyl-6a-(benzoyl) jaeschkeanadiol, and 2a-acetyl-6a-(p-anisoyl) jaeschkeanadiol29. Additionally, ferulenol, a constituent of F. communis, has garnered attention owing to its various pharmacological effects, including antimicrobial, anticoagulant, antiproliferative, and antiappetitant activities. Recently, Gliszczynska et al., explored sesquiterpene coumarins, focusing on ferulenol, as promising lead compounds for drug discovery30. Notably, this diverse array of secondary metabolites substantially contributes to the antioxidant capabilities of F. communis while simultaneously conferring a broad spectrum of pharmacological and biological functionalities31. Phenolic acids, known for their various biological applications, are the primary polyphenols produced by plants and serve as precursors for bioactive molecules used in the therapeutic, cosmetic, and food industries. The health-promoting effects of these phenolic compounds are diverse and may include anticancer, anti-inflammatory, antioxidant, antimicrobial, and anti-aging properties32.
Total phenolic content
Phenolic compounds are the primary phytochemicals responsible for the biological activities of Ferula3. Moreover, the solvent influences the quantity and selectivity of the extracted components33. Phenolic compounds and other phytochemicals containing hydroxyl groups are primarily soluble in polar solvents34. As shown in Fig. 2a, our results present the effects of different extraction solvents (ethanol, methanol, water, hexane, ethyl acetate, and chloroform) on the total polyphenol content of F. communis. The ethanol extract exhibited the highest total polyphenol content (62.20 ± 0.11 mg GAE/g DW), followed by the methanol (60.82 ± 0.32 mg GAE/g DW) and aqueous (44.04 ± 0.22 mg GAE/g DW) extracts. Acetone, ethyl acetate, chloroform, and hexane showed lower values at 19.76 ± 1.19, 15.24 ± 0.60, 13.94 ± 0.92 and 6.93 ± 0.43 mg GAE/g DW, respectively. Comparing the TPC of the solvents, the order established was: ethanol > methanol > water > acetone > ethyl acetate > chloroform > hexane. The extract of Tunisian F. communis fruit exhibited the highest TPC (422 mg GAE/g DW), followed by flowers (207.21 mg GAE/g DW) and stems (129.86 mg GAE/g DW)35. In addition, Gamal and Atraimki quantified the total phenolics in n-butanol and ethyl acetate extracts (44.7–55.8 mg GAE/g DW)36.
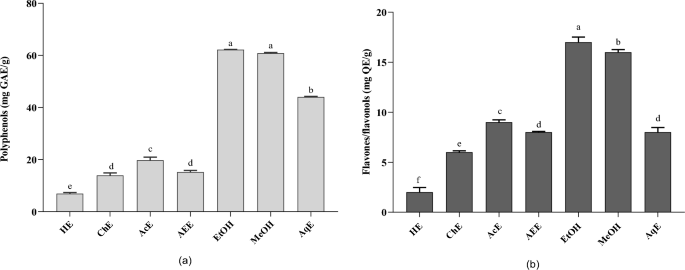
Effect of different solvent extractions on the (a) polyphenol content; (b) flavone/flavanol content; results sharing the same letter in the same test are not significantly different according to Tukey’s multiple range test (p < 0.05). Data are the means of three replicates. HE, Hexane extract; ChE, chloroform extract; AEE, ethyl acetate extract; EtOH, ethanolic extract; MeOH, methanol extract; AqE, aqueous extract.
The extraction efficiencies varied considerably among the solvents, resulting in differences in the concentrations of total phenolics. The TPC of the extracts is influenced by the polarity of the extracting solvent and the solubility of chemical constituents in the solvent37. Solvents with greater polarity extracted more TPC29. Thus, more polar solvents such as ethanol, methanol, and acetone extracted more TPC. Ethanol extracts have the highest TPC, followed by methanol and acetone extracts17. This is consistent with a previous study, in which methanol and ethyl acetate extracted greater amounts of phenolic compounds than the other two37. These results confirm the critical role of extraction solvents in modifying the TPC recovery from plant material. Furthermore, based on the above results, it is clear that the ethanol and methanol extracts of wild F. communis fruit examined in this study exhibited comparable or greater efficiency of TPC extraction than previously reported.
Total flavonoid content
The total flavonoid content (TFC) from F. commnis exhibited a trend similar to that of TPC (Fig. 2b). The ethanol extract of Ferula exhibited the highest extraction yields of flavonoids (17.09 mg QE/g DW), followed by the methanol extract with 16.91 ± 0.26 mg QE/g DW, and the acetone extract (9.31 ± 0.24 mg GAE/g DW). Aqueous, ethyl acetate, chloroform, and hexane showed lower extraction yields with values of 9.31 ± 0.24, 8.97 ± 0.47, 8.68 ± 0.08, 6.23 ± 0.13, and 2.39 ± 0.47 mg GAE/g DW, respectively. Owing to their polarities, the extraction of TFC significantly varied among the solvents. For the TFC, the extracts were classified in decreasing order of efficiency: ethanol > methanol > acetone > water > ethyl acetate > chloroform > hexane. The methanol extracts of various aerial parts of F. communis L. showed the highest concentration in flowers, followed by fruits and stems, with values of 48.77, 14.23, and 13.37 mg QE/g DW, respectively. Our results, reflecting varying TFC amounts extracted by different solvents, can explain the presence of diverse groups of soluble flavonoids in F. communis fruits with distinct polarities. In addition, reported variations in bioactive compound values for Ferula in the literature may stem from factors such as varietal differences, environmental conditions, agronomic practices, and analytical methods.
Antioxidant activity of F. communis
Antioxidant activity was not assessed using a single antioxidant test30; rather, several in vitro procedures, including the DPPH, ABTS, RP, and TAC assays (Fig. 3), were used to assess antioxidant activity. As antioxidant test models vary in different respects, it is difficult to directly compare results9,34. Therefore, performing several screens using ex vivo models and in vivo studies is essential31. DPPH is a nitrogenous organic radical with free electrons, and is an easy and valuable spectrophotometric method for screening or measuring antioxidant activity38. When antioxidants react with DPPH, stable free radicals pair with hydrogen donors and are reduced to DPPH-H28. ABTS is a target molecule used to assess the reactivity of antioxidant samples in the presence of peroxides15. The radical cation of ABTS (ABTS+) is synthesized by oxidizing ABTS with potassium permanganate, potassium persulfate, or 2,2′-azo-bis (2-amidinopropane)39. The ABTS assay reduces labor time, material cost, and sample volume, while decreasing color40. The RP assay is based on the principle that substances with a reducing potential react with potassium ferricyanide (Fe3+) to form potassium ferrocyanide (Fe2+)41, consequently reacting with ferric chloride to form a ferric-ferrous complex with an absorption maximum at 700 nm42.
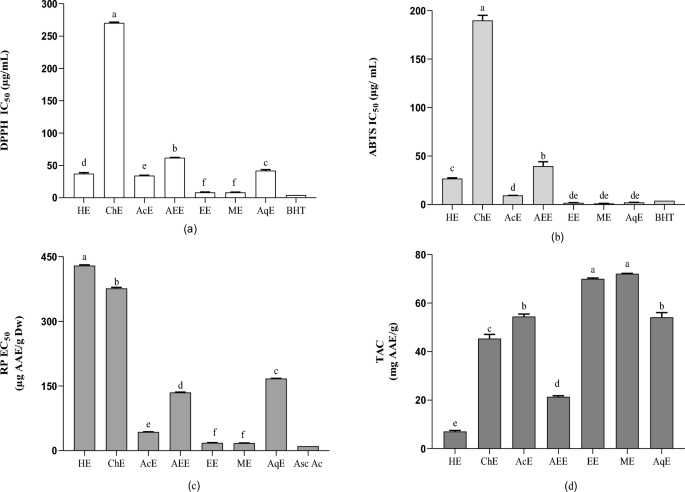
Antioxidant activities: (a) IC50 of the DPPH test; (b) IC50 of the ABTS test; (c) EC50 of reducing power; (d) the content of total antioxidant capacity (TAC). Each response is the average of triplicate with standard error. Values in the same column followed by different letters are significantly different according to Tukey’s multiple range test (p < 0.05). HE, Hexane extract; ChE, chloroform extract; AEE, ethyl acetate extract; EtOH, ethanolic extract; MeOH, methanolic extract; AqE, aqueous extract; IC50, the extract concentration provides 50% inhibition; DPPH, 2,2‐diphenyl‐1‐picrylhydrazyl; ABTS, 2,2′-azino-bis (3-ethylbenzothiazoline-6-sulphonic acid); TAC, total antioxidant capacity.
The greatest DPPH scavenging capacity was demonstrated by the methanol extract with an IC50 value of 8.09 µg/mL, followed by the ethanol, acetone, and aqueous extracts with IC50 values of 8.25, 34.23, and 37.28 μg/mL, respectively. The scavenging efficacy decreased in the following order: ChE < AEE < AqE < AcE < EtOH < MeOH. These results demonstrated that highly polar solvents, including ethanol and methanol, are effective for the extraction of antioxidants with efficient free radical scavenging properties and are more effective than intermediate-polarity solvents, such as acetone and hexane, but not chloroform (Fig. 3a).
The results of this study are consistent with those of previous studies wherein potent antioxidant activities were observed for both ethyl acetate (IC50 = 4.4 ± 0.1 μg/mL) and n-butanol extracts36. The antioxidant activity assessed by measuring the scavenging power of DPPH on gum resin of the Algerian F. communis L. harvested in the Sahara had an IC50 of 5.08 mg/mL43. Roots of another species, Ferula gummosis Boiss, was evaluated for antioxidant activity using the DPPH assay and displayed an IC50 = 579.6 ± 19.4 μg/mL44. The RP of F. communis extracts was highest in the MeOH extract, with an EC50 of 17.63 µg, followed by the ethanol, acetone, and ethyl acetate extracts, with EC50 values of 17.63, 43.4, and 135 µg AAE/g DW, respectively. Hexane exhibited the lowest RP with an EC50 of 429.3 μg AAE/g DW. The high RP of the F. communis methanol extract is probably due to clusters of hydroxyls in phenolic compounds, serving as electron donors. Therefore, antioxidants act as both reducers and activators45. The RP of a compound can considerably indicate its antioxidant potential46. However, no study has evaluated the RP of F. communis fruits.
The results of the ABTS assay showed significant variations among the extracts (Fig. 3b). The ethanol extract expressed the highest percentage of ABTS free radical inhibition (IC50 = 1.88 ± 0.07 μg/mL). Alternatively, the chloroform extract (ChE) showed the lowest ABTS radical scavenging capacity (IC50 = 189.87 μg/mL). The activity (ABTS) results for F. communis extracts have not been previously published.
Reduced antioxidant activity, as determined by the TAC assay (Fig. 3d), was similar to the activities determined by the other tests. The MeOH and EtOH extracts exhibited the highest values (72.15 ± 0.11 and 70.04 ± 0.34 mg AAE/g, respectively). In addition, the hexane sample showed the least RP (7.05 ± 0.46 g AAE/g). The methanol, ethanol, acetone, and aqueous extracts showed powerful and significant correlations in the four antioxidant tests (Table 4), possibly related to their phytochemical constituents, primarily phenolic components. These results supported the investigation of the antimicrobial activity of these extracts.
Antimicrobial studies
Antibacterial activity
The antimicrobial activities of F. communis extracts, including EtOH, AqE, and AcE, were evaluated against the examined microorganisms. EtOH and AcE extracts exhibited significant antibacterial activity compared to the reference antibiotic, oxacillin, as determined by the inhibition zone diameter and MIC values (Tables 5, Fig. 4). The AcE extract displayed the highest inhibitory activity against P. mirabilis with an inhibition diameter of 19.00 ± 1.00 mm and an MIC of 2.50 ± 0.00 mg/mL, followed by E. coli with 11.50 ± 1.50 mm of inhibition zone and an MIC of 2.50 ± 0.00 mg/mL. The extract also exhibited inhibitions of 11.00 and 9.00 mm against S. aureus and B. subtilis, respectively.

Visual observations of the inhibitory effects of Ferula communis extracts against pathogenic bacteria, including Staphylococcus aureus ATCC6633, Escherichia coli K12, Bacillus subtilis DSM6333, and Proteus mirabilis ATCC29906.
The EtOH extract, which contained the largest amounts of TPC and TFC, also exhibited the greatest TAC and was the most effective against all bacteria. The greatest activity against E. coli, with an inhibition diameter of 14.00 ± 1.00 mm, an MIC of 0.312 ± 0.00 mg/mL, and the least zone of 9.00 ± 0.00 mm, was observed for S. aureus. AcE was more potent than EtOH for both extracts. Both extracts showed bactericidal activity against the four tested bacterial strains, whereas the aqueous extract of F. communis did not inhibit any of the examined strains. These differences in inhibition diameters can be attributed to the chemical composition of F. communis, and its antibacterial activity may primarily be because of the compounds in these extracts (EtOH, AqE, and AcE). The results showed that the ethanol and acetone extracts inhibited the four bacterial strains, with the ethanol extract having the best antibacterial activity.
Resorcinol, ferulic and syringic acids, and coumarin are renowned for their antibacterial activity. Compared to streptomycin sulfate, the antibacterial activities of the constituents extracted from the rhizome of F. communis revealed that the sesquiterpene 14-(hydroxycinnamoyloxy)-dauc-4,8-diene possesses vigorous activity against S. aureus, B. subtilis, E. faecalis, and S. durans7. The 80% ethanol-solubilized fraction of F. communis exhibited the greatest antibacterial effects on all assayed bacteria, whereas the aqueous-solubilized fractions did not exhibit any effect on the clinical isolates of Strep. pyogenes and Strep. pneumonia47. Alternatively, the isolated oil of Tunisian Ferula lutea was tested for its antimicrobial activity using disc diffusion and microdilution assays against six gram-positive and five gram-negative bacteria and eight species of the genus Candida48. Oil from flowers of F. lutea exhibited antibacterial and anticandidal activity, with an MIC of 39 g/mL against S. aureus, S. epidermidis, and E. coli, and an MIC of 156 g/mL against C. albicans. The type of solvent used considerably affected the antibacterial activity of extracts of Ferula inflorescence. These antibacterial compounds were marginally more abundant in gram-negative bacteria than in gram-positive bacteria. This result is likely due to the shape and composition of the cell walls of gram-positive bacteria, which have a thick layer of peptidoglycan with teichuronic acid and glycopolymers such as teichoic acids, making them less sensitive to the action of plant extracts49.
Antifungal activity
In in vitro disk diffusion assays, the antifungal activities of F. communis extract against C. albicans, A. niger, A. flavus, and F. oxysporum were compared to that of the fungicide fluconazole. AcE exhibited significant activity against F. oxysporum, with a percent inhibition of 20.6 ± 1.4% and an MIC value of 5.0 ± 0.0 mg/mL. Additionally, it showed a percentage of inhibition of 19.0 ± 1.0% and an MIC value of 5 mg/mL against A. niger. The EtOH extract resulted in a small zone of inhibition of only 12.50 ± 2.50 mm) against C. albicans with an MIC of 2.50 mg/mL (Table 6, Fig. 5). Additionally, the ethanol extract exhibited an inhibition percentage of 26.0 ± 2.0% and an MIC of 2.5 mg/mL against A. niger. The percent inhibition of F. oxysporum was 13.3 ± 2.6 with an MIC of 5 mg/mL. The aqueous F. communis extract demonstrated antifungal activities against C. albicans with a 13.00 ± 1.00 mm of inhibition zone and an MIC of 5 mg/mL for MIC, as this extract has no effect against A. niger and F. oxysporum. However, the three F. communis extracts did not exhibit antifungal effects against A. flavus.

Visual observations of the inhibitory effects of Ferula communis extracts against pathogenic fungi, including Candida albicans ATCC10231, Aspergillus niger MTCC282, Aspergillus flavus MTCC9606, and Fusarium oxysporum MTCC9913.
Studies on the activity of F. communis extracts are scarce. More detailed information on the antimicrobial activities, especially the antifungal properties of F. communis, is required. The in vitro antibacterial activities of F. communis root extracts on colonies and conidia of Botryotinia fuckeliana, Penicillium digitatum, P. expansum, Monilinia laxa, M. fructigena, and Aspergillus spp were determined50; minimal antifungal activity on colony growth was observed, whereas root extracts exhibited the greatest effects on the growth of fungal colonies. However, the root extract was unable to inhibit conidia germination. These results are encouraging because the extract was not a pure product; therefore, the antimicrobial activity may be owing to several compounds related to the presence of bioactive metabolites.
Correlation
Multivariate analyses, such as principal component analysis (PCA), can reduce a complex dataset, make it more interpretable, and reveal relationships between or among parameters. In the present study, the correlations between antioxidant content, antioxidant activities, and antioxidant and antibacterial activities were examined. PCA results e(Fig. 6) showed that 91.2% of data variability is attributable to the first two principal components, PC1 (Component 1) and PC2 (Component 2), confirming that the correlation test is considered an excellent tool to reveal any relationship between different parameters studied. The first three PCs explained 100% of the data variance, which was sufficient to illustrate all the variables adequately. In this analysis, if two vectors subtend at a slight angle to each other, the two variables they represent strongly correlate. The plots of the scores for PC1 and PC2 showed a strong positive correlation among TPC, TFC, and TAC. The results of the DPPH, ABTS, and RP assays revealed a strong negative correlation between the bioactive content, as represented by the presence of phenolic and flavones/flavanol compounds, and antioxidant activity. Additionally, ethanol and methanol positively and negatively correlated with hexane, respectively. However, a positive correlation between DPPH and ABTS was observed, indicating that the most potent moieties for scavenging DPPH radicals showed the highest ABTS. The ingenious display of the polar heat map revealed a row and column hierarchical cluster structure in the data matrix (similarity/dissimilarity). This made it possible to group different samples based on their similarities. The analysis based on Pearson’s correlation showed lower concentrations in brown and greater concentrations in dark blue (Fig. 7). Plant extracts of the genus Ferula have been arranged into two groups, one of which is the ethanol and aqueous extract. The polar heatmap also shows the correlation between the antioxidant content and antioxidant and antibacterial activities.

Principal component analysis.

Polar heatmap with dendrogram for antioxidant and antibacterial correlations.
Molecular docking simulations
In investigating potential mechanisms of inhibition via antioxidant, antibacterial, and antifungal activities, nine molecular docking simulations were conducted on three targeted proteins with three major compounds of F. communis: syringic acid (C7), 3-hydroxybenzoic acid (C8), and p-coumaric acid (C12). These compounds were extracted at rates of 12.87, 17.34, and 29.42%, respectively (Fig. 8). Three major compounds were docked to the NADPH oxidase protein from Lactobacillus sanfranciscensis. Encoded by 2CDU.pdb, this protein exhibits binding energies of -5.28, -5.76, and 5.65 kcal/mol. It shares various common intermolecular interactions, forming hydrogen bonds with Ser115, Ala11, Lys134, and Gly12 amino acid residues (AAR) in the A chain. Additionally, it forms carbon–hydrogen bonds toward Thr9 AAR. When three major compounds were docked to the FimH lectin protein from E. coli K-12, encoded in the PDB by 4XO8.pdb, they formed another family of chemical bonds with binding energies of -5.38, -5.47, and -5.01 kcal/mol. Common bonds included one hydrogen bond with Arg92 AAR and one Pi-Pi alkyl bond with Pro91 AAR. This similarly was observed between syringic acid (C7) and p-coumaric acid (C12) (Fig. 9). 3-Hydroxybenzoic acid created another class of chemical bonds, forming six hydrogen bonds with Asp47, Asp54, Asp140, Phe1, Gln133, and Asn46 AAR, in addition to one Pi-alkyl bond with Il13 AAR in the A chain.

Results of intermolecular interactions between the NADPH oxidase protein from Lactobacillus sanfranciscensis (2CDU.pdb) and major compounds of Ferula communis (syringic acid, 3-hydroxybenzoic acid, and p-coumaric acid) in two and three dimensions.

Results of intermolecular interactions between the FimH lectin protein from Escherichia coli K12 (4XO8.pdb) and major compounds of Ferula communis (syringic acid, 3-hydroxybenzoic acid, and p-coumaric acid) in two and three dimensions, produced.
The mechanism of antifungal activities of the three major constituents in F. communis extracts was examined by docking to the crystal structure of sterol 14-alpha demethylase (CYP51) from C. albicans encoded in the PDB by 5TZ1.pdb. The binding energies were -5.88, -6.93, and -6.77 kcal/mol, corresponding to C7, C8, and C12, respectively (Fig. 10). The major compounds of the studied plant produced similar chemical interactions, with three hydrogen bonds created with Arg381, Tyr118, and His468 AAR, and more than one bond resulting in a Cys470 AAR in the A chain of the targeted protein. The docking validation protocol for these nine molecular simulations was successfully evaluated. The three major compounds of the F. communis plant were strongly docked to the active sites of each targeted protein, as confirmed with the assistance of the protein plus server (https://proteins.plus/)51. The active sites of the proteins were defined as co-crystallized ligands (or native ligands) (Fig. 11). Three amino acids, Ala11, Lys134, and Thr9, were active sites for the NADPH oxidase protein. Amino acids Asp47, Asp54, Asp140, Phe1, and Gln133 AARs were the active sites of the antibacterial protein. Arg381, Tyr118, and His468 are the active sites of antifungal proteins. Binding energies in Kcal/mol are primarily negative and do not exceed the − 5.000 kcal/mol threshold, demonstrating molecular stability52. These results are consistent with those of Jeddi et al.53, who demonstrated antioxidant and antibacterial effects toward the same targeted receptors, NADPH oxidase and FimH lectin proteins, for two major compounds of the essential oil extracted from the mill of Lavandula angustifolia. In that study, the major compounds in F. communis extracts were similarly docked to the active sites of each responsible protein, sharing broadly equivalent intermolecular interactions. The biological activity of antifungal molecules from natural sources toward lanosterol 14-alpha demethylase (CYP51) involves similar intermolecular interactions, including the active sites of the CYP51 protein from C. albicans54.

Results of intermolecular interactions between the CYP51 protein from Candida albicans (5TZ1.pdb) and major compounds (syringic acid, 3-hydroxybenzoic acid, and p-coumaric acid) in two and three dimensions.

Active sites of antioxidant (2CDU.pdb), antibacterial (4XO8.pdb), and antifungal (5TZ1.pdb) proteins complexed with flavin-adenine dinucleotide (-A-), heptyl alpha-d-mannopyranoside (-B-), and tetrazole-based antifungal drug candidate VT1161 (VT1), respectively.
Conclusion
In this study, we examined the effects of various solvents on extracts. The extracts demonstrated considerable antioxidant activity across various antioxidant methods, and exhibited antimicrobial activity against bacteria and yeasts. The extract contained moderate amounts of flavonoids and polyphenols. The bioactive substances identified in the non-volatile section of F. communis via the HPLC–DAD study are most likely responsible for these effects. Information on ligand–protein interactions involving antioxidant and antimicrobial proteins has provided valuable insights into binding affinities and interactions. These findings support further exploration via in vivo and in situ experiments, as well as the investigation of other biological activities of this plant.
Data availability
All data generated or analyzed during this study are included in this published article.
References
Nouioura, G. et al. Assessment of the acute and subacute toxicity of the aqueous extract of Moroccan Ferula communis fruit in a mouse model. Saudi Pharm. J. 31(8), 101701. https://doi.org/10.1016/j.jsps.2023.101701 (2023).
Bellakhdar, J. Médecine traditionnelle et toxicologie ouest-sahariennes: contribution à l’étude de la pharmacopée marocaine (1978).
Mrid, R. B. et al. Dittrichia viscosa L. leaves: A valuable source of bioactive compounds with multiple pharmacological effects. Molecules 27(7), 2108. https://doi.org/10.3390/molecules27072108 (2022).
El Alaoui Fares, F., Cauwet, A., Fraigui, O., Lamnaouer, D. & Gorenflot, R. Le Genre Ferula L (apiaées) au Maroc. Revu de cytologie et biologie vegetales-le botaniste, 3–19, (2005).
Bellakhdar, J. Pharmacopée marocaine traditionnelle: Medicine arabe ancienne et savoirs populaires 739–759 (Ibis Press, 1997).
Bellakhdar, J. Plantes médicinales au Maghreb et soins de base: précis de phytothérapie moderne, 386 (2004).
Al-Yahya, M. A., Muhammad, I., Mirza, H. H. & El-Feraly, F. S. Antibacterial constituents from the rhizomes of Ferula communis. Phytother. Res. 12(5), 335–339. https://doi.org/10.1002/(SICI)1099-1573(199808)12:5%3c335::AID-PTR306%3e3.0.CO;2-H (1998).
Golmohammadi, F. Traditional knowledge and economic importance of Ferula assa-foetida in the rural areas of southeastern Iran. Afr. J. Plant Sci. 16(6), 148–156. https://doi.org/10.5897/AJPS2017.1617 (2022).
Nouioura, G. et al. Optimization of a new antioxidant formulation using a simplex lattice mixture design of Apium graveolens L., Coriandrum sativum L., and Petroselinum crispum M. grown in Northern Morocco. Plants 12(5), 1175. https://doi.org/10.3390/plants12051175 (2023).
Nouioura, G. et al. The protective potential of Petroselinum crispum (Mill.) Fuss. on paracetamol-induced hepatio-renal toxicity and antiproteinuric effect: A biochemical, hematological, and histopathological study. Medicina 59(10), 1814. https://doi.org/10.3390/medicina59101814 (2023).
El Barnossi, A., Moussaid, F. & Iraqi Housseini, A. Antifungal activity of Bacillus sp. Gn-A11-18isolated from decomposing solid green household waste in water and soil against Candida albicans and Aspergillus niger. E3S Web Conf. 150, 02003. https://doi.org/10.1051/e3sconf/202015002003 (2020).
Rolta, R., Kumar, V., Sourirajan, A., Upadhyay, N. K. & Dev, K. Bioassay guided fractionation of rhizome extract of Rheum emodi wall as bio-availability enhancer of antibiotics against bacterial and fungal pathogens. J. Ethnopharmacol. 257, 112867. https://doi.org/10.1016/j.jep.2020.112867 (2020).
Alruways, M. W. Antimicrobial effects of Ferula species—an herbal tactic for management of infectious diseases. J. King Saud Univ. Sci. 35(7), 102806. https://doi.org/10.1016/j.jksus.2023.102806 (2023).
Brand-Williams, W., Cuvelier, M. E. & Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 28(1), 25–30. https://doi.org/10.1016/S0023-6438(95)80008-5 (1995).
Re, R. et al. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 26(9–10), 1231–1237. https://doi.org/10.1016/S0891-5849(98)00315-3 (1999).
Oyaizu, M. Studies on product of browning reaction prepared from glucose amine. Jpn. J. Nutr. 44, 307–315 (1986).
Wang, H., Cao, G. & Prior, R. L. Total antioxidant capacity of fruits. J. Agric. Food Chem. 44(3), 701–705. https://doi.org/10.1021/jf950579y (1996).
Singleton, V. L., Orthofer, R. & Lamuela-Raventós, R. M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Methods in Enzymology Vol. 299 152–178 (Elsevier, 1999). https://doi.org/10.1016/S0076-6879(99)99017-1.
SanMiguel-Chávez, R. Phenolic antioxidant capacity: a review of the state of the art. In Phenolic Compounds-Biological Activity (eds Soto-Hernndez, M. et al.) (InTech, 2017). https://doi.org/10.5772/66897.
Mssillou, I. et al. Phytochemical characterization, antioxidant activity, and in vitro investigation of antimicrobial potential of Dittrichia viscosa L. leaf extracts against nosocomial infections. Acta Ecol. Sin. 42(6), 661–669. https://doi.org/10.1016/j.chnaes.2021.09.021 (2022).
Rolta, R. et al. Bioassay guided fractionation of phytocompounds from Bergenia ligulata: A synergistic approach to treat drug resistant bacterial and fungal pathogens. Pharmacol. Res. Mod. Chin. Med. 3, 100076. https://doi.org/10.1016/j.prmcm.2022.100076 (2022).
Sarker, S. D., Nahar, L. & Kumarasamy, Y. Microtitre plate-based antibacterial assay incorporating resazurin as an indicator of cell growth, and its application in the in vitro antibacterial screening of phytochemicals. Methods 42(4), 321–324. https://doi.org/10.1016/j.ymeth.2007.01.006 (2007).
Bouslamti, M. et al. Total polyphenols content, antioxidant and antimicrobial activities of leaves of Solanum elaeagnifolium Cav. from Morocco. Molecules 27(13), 4322. https://doi.org/10.3390/molecules27134322 (2022).
D. Systèmes, Free Download: BIOVIA Discovery Studio Visualizer, Dassault Systèmes. https://discover.3ds.com/discovery-studio-visualizer-download (Accessed 9 Oct 2023). (2020) [En ligne].
Salaria, D. et al. In vitro and in silico analysis of Thymus serpyllum essential oil as bioactivity enhancer of antibacterial and antifungal agents. J. Biomol. Struct. Dyn. 40(20), 10383–10402. https://doi.org/10.1080/07391102.2021.1943530 (2022).
Norgan, A. P., Coffman, P. K., Kocher, J.-P.A., Katzmann, D. J. & Sosa, C. P. Multilevel parallelization of AutoDock 4.2. J. Cheminform. 3(1), 12. https://doi.org/10.1186/1758-2946-3-12 (2011).
El Fadili, M. et al. QSAR, ADME-Tox, molecular docking and molecular dynamics simulations of novel selective glycine transporter type 1 inhibitors with memory enhancing properties. Heliyon 9(2), e13706. https://doi.org/10.1016/j.heliyon.2023.e13706 (2023).
Shekhar, T. C. & Anju, G. Antioxidant activity by DPPH radical scavenging method of Ageratum conyzoides Linn. leaves. Am. J. Ethnomedicine 1(4), 7 (2014).
Aazza, S. Application of multivariate optimization for phenolic compounds and antioxidants extraction from Moroccan Cannabis sativa waste. J. Chem. 2021, 1–11. https://doi.org/10.1155/2021/9738656 (2021).
de Menezes, B. B., Frescura, L. M., Duarte, R., Villetti, M. A. & da Rosa, M. B. A critical examination of the DPPH method: Mistakes and inconsistencies in stoichiometry and IC50 determination by UV–Vis spectroscopy. Anal. Chim. Acta 1157, 338398. https://doi.org/10.1016/j.aca.2021.338398 (2021).
Eivazkhani, F. et al. Effects of N-acetyl-l-cystein antioxidant on ex vivo culture of vitrified premature mouse ovarian tissue. Biopreserv. Biobank. 20(4), 331–339. https://doi.org/10.1089/bio.2021.0147 (2022).
Kumar, N. & Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 24, e00370. https://doi.org/10.1016/j.btre.2019.e00370 (2019).
Mohammad Salamatullah, A. et al. Effects of different solvents extractions on total polyphenol content, HPLC analysis, antioxidant capacity, and antimicrobial properties of peppers (red, yellow, and green (Capsicum annum L.)). Evid. Based Complement. Alternat. Med. 2022, 1–11. https://doi.org/10.1155/2022/7372101 (2022).
Iloki-Assanga, S. B. et al. Solvent effects on phytochemical constituent profiles and antioxidant activities, using four different extraction formulations for analysis of Bucida buceras L. and Phoradendron californicum. BMC Res. Notes 8(1), 396. https://doi.org/10.1186/s13104-015-1388-1 (2015).
Rahali, F. Z. et al. Phytochemical composition and antioxidant activities of different aerial parts extracts of Ferula communis L. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 153(2), 213–221. https://doi.org/10.1080/11263504.2018.1461696 (2019).
Gamal, M. A. B. & Atraiki, R. A. M. Phytochemical constituents of Ferula communis plant extracts and their antimicrobial and antioxidant activity. Lebda Med. J 1, 6–9 (2015).
Babbar, N., Oberoi, H. S., Sandhu, S. K. & Bhargav, V. K. Influence of different solvents in extraction of phenolic compounds from vegetable residues and their evaluation as natural sources of antioxidants. J. Food Sci. Technol. 51(10), 2568–2575. https://doi.org/10.1007/s13197-012-0754-4 (2014).
Gülçin, İ. Antioxidant activity of food constituents: An overview. Arch. Toxicol. 86(3), 345–391. https://doi.org/10.1007/s00204-011-0774-2 (2012).
Thaipong, K., Boonprakob, U., Crosby, K., Cisneros-Zevallos, L. & Hawkins Byrne, D. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 19(6–7), 669–675. https://doi.org/10.1016/j.jfca.2006.01.003 (2006).
Vinci, G., D’Ascenzo, F., Maddaloni, L., Prencipe, S. A. & Tiradritti, M. The influence of green and black tea infusion parameters on total polyphenol content and antioxidant activity by ABTS and DPPH assays. Beverages 8(2), 18. https://doi.org/10.3390/beverages8020018 (2022).
Singhal, M., Paul, A., Singh, H. P., Dubey, S. K. & Gaur, K. Evaluation of Reducing Power Assay of Chalcone Semicarbazones, 8 (2011).
Langley-Evans, S. C. Antioxidant potential of green and black tea determined using the ferric reducing power (FRAP) assay. Int. J. Food Sci. Nutr. 51(3), 181–188. https://doi.org/10.1080/09637480050029683 (2000).
Youmbai, A. et al. Characterization and biological activities of a polysaccharidic extract from Ferula communis L. (Apiaceae) harvested in Sahara, Phytothérapie, Publication ahead of print, (2021) [En ligne]. Disponible sur. https://doi.org/10.3166/phyto-2021-0292.
Ebrahimzadeh, M., Nabavi, S., Nabavi, S. & Dehpour, A. Antioxidant activity of hydroalcholic extract of Ferula gummosa Boiss roots. Eur. Rev. Med. Pharmacol. Sci. 15, 658–664 (2011).
Lobo, V., Patil, A., Phatak, A. & Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 4(8), 118. https://doi.org/10.4103/0973-7847.70902 (2010).
Hossain, M. M., Uddin, M. S., Baral, P. K., Ferdus, M. & Bhowmik, S. Phytochemical screening and antioxidant activity of Ipomoea hederifolia stems: A potential medicinal plant. Asian J. Nat. Prod. Biochem. https://doi.org/10.13057/biofar/f200201 (2022).
Unasho, A. et al. Investigation of antibacterial activities of Albizia gummifera and Ferula communis on Streptococcus pneumoniae and Streptoccus pyogenes. Ethiop. Med. J. 47(1), 25–32 (2009).
Znati, M. et al. Chemical composition and in vitro evaluation of antimicrobial and anti-acetylcholinesterase properties of the flower oil of Ferula lutea. Nat. Prod. Commun. 7(7), 1934578X1200700. https://doi.org/10.1177/1934578X1200700738 (2012).
Bonnet, J. Rôles coopératifs du peptidoglycane et des acides téichoïques dans le remodelage de la paroi et la division cellulaire de Streptococcus pneumoniae, (Université Grenoble Alpes, 2017) [En ligne]. Disponible sur: https://tel.archives-ouvertes.fr/tel-02008878/file/BONNET_2017_archivage.pdf.
Mamoci, E. et al. Chemical composition and in vitro activity of plant extracts from Ferula communis and Dittrichia viscosa against postharvest fungi. Molecules 16(3), 2609–2625. https://doi.org/10.3390/molecules16032609 (2011).
Zentrum für Bioinformatik, Universität Hamburg - Proteins Plus Server, 29 (2022). [En ligne]. Disponible sur: https://proteins.plus/.
Xing, S. et al. Antifungal activity of volatile components from Ceratocystis fimbriata and its potential biocontrol mechanism on Alternaria alternata in postharvest cherry tomato fruit. Microbiol. Spectr. 11(1), e02713-e2722. https://doi.org/10.1128/spectrum.02713-22 (2023).
Jeddi, M. et al. Antimicrobial, antioxidant, α-amylase and α-glucosidase inhibitory activities of a chemically characterized essential oil from Lavandula angustifolia Mill.: In vitro and in silico investigations. Biochem. Syst. Ecol. 111, 104731. https://doi.org/10.1016/j.bse.2023.104731 (2023).
Sama-ae, I., Pattaranggoon, N. C. & Tedasen, A. In silico prediction of antifungal compounds from natural sources towards lanosterol 14-alpha demethylase (CYP51) using molecular docking and molecular dynamic simulation. J. Mol. Graph. Model. 121, 108435. https://doi.org/10.1016/j.jmgm.2023.108435 (2023).
Acknowledgements
The authors extend their appreciation to Researchers Supporting Project (No. RSPD2024R816), King Saud University, Riyadh, Saudi Arabia.
Funding
This research was funded by the Researchers Supporting Project (No. RSPD2024R816), King Saud University, Riyadh, Saudi Arabia.
Author information
Authors and Affiliations
Contributions
Conceptualization, G.N., E.D., and B.L.; methodology, M.B., M.E., and A.B.; software, M.E.; formal analysis, G.N., and A.E.; data curation, G.N., and H.L; writing—original draft preparation, G.N., and J.P.G.; writing—review and editing, G.N., and M.E. writing—review and editing. Supervision, B.L., and E.D. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Nouioura, G., El fadili, M., El Barnossi, A. et al. Comprehensive analysis of different solvent extracts of Ferula communis L. fruit reveals phenolic compounds and their biological properties via in vitro and in silico assays. Sci Rep 14, 8325 (2024). https://doi.org/10.1038/s41598-024-59087-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-59087-3
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
