
Abstract
Amyloid β (Aβ) aggregates into two distinct fibril and amorphous forms in the brains of patients with Alzheimer’s disease. Adenosine triphosphate (ATP) is a biological hydrotrope that causes Aβ to form amorphous aggregates and inhibit fibril formation at physiological concentrations. Based on diffracted X-ray blinking (DXB) analysis, the dynamics of Aβ significantly increased immediately after ATP was added compared to those in the absence and presence of ADP and AMP, and the effect diminished after 30 min as the aggregates formed. In the presence of ATP, the β-sheet content of Aβ gradually increased from the beginning, and in the absence of ATP, the content increased rapidly after 180 min incubation, as revealed by a time-dependent thioflavin T fluorescence assay. Images of an atomic force microscope revealed that ATP induces the formation of amorphous aggregates with an average diameter of less than 100 nm, preventing fibrillar formation during 4 days of incubation at 37 °C. ATP may induce amorphous aggregation by increasing the dynamics of Aβ, and as a result, the other aggregation pathway is omitted. Our results also suggest that DXB analysis is a useful method to evaluate the inhibitory effect of fibrillar formation.
Similar content being viewed by others
Introduction
One of the major hallmarks of Alzheimer’s disease (AD) is the formation of senile plaques that primarily contain amyloid β (Aβ) fibrils within the brain. Before the senile plaque formation, diffuse plaques of amorphous Aβ aggregates are found in the brains of individuals with AD1. The fibril forms a β-sheet structure, and the amorphous aggregates are formed from the most toxic oligomers by pathways that are on- and off-pathways, respectively. Their toxicity differs in various reports, probably due to the distinct preparation conditions, which determine the aggregation forms2,3,4. Regardless, the on- and off-pathway aggregates show distinct toxicity, and controlling aggregation to decrease toxicity can be an effective therapy for AD. However, evaluating aggregates is challenging, especially at the early stages, due to the instability of the Aβ monomer and the aggregates.
Several Aβ binding molecules have been reported to reduce oligomer toxicity by accelerating the oligomer into the on-pathway and off-pathway aggregates5,6,7,8. Adenosine triphosphate (ATP) is a hydrotrope that solubilizes hydrophobic molecules at physiological concentrations between 5 and 10 mM9. Aβ misfolding is prevented by ATP at lower concentrations, i.e., 0.5 mM10. Molecular dynamics simulations suggest that the hydrophobic adenosine of ATP interacts with Aβ and that ATP converts the oligomer into the off-pathway11. Furthermore, ATP concentrations decrease in the neocortex of AD patients as AD progresses, suggesting a molecular link between ATP and Aβ12. However, the detailed mechanisms by which ATP induces the off-pathway are unclear.
Diffracted X-ray blinking (DXB) monitors the rotational motions of single protein molecules labeled with gold nanocrystals13. DXB differentiates the dynamics of Aβ42, Aβ40, and Aβ38 on a subnanometer scale and reveals that Aβ42 possesses lower dynamics among the isoforms, especially after 96 h incubation14. Because Aβ42 exhibits a higher tendency to aggregate among the isoforms due to the C-terminal hydrophobic amino acid residues, the decrease in dynamics may represent the stacked molecules on the surface of aggregates. Thioflavin T (ThT) selectively interacts with the β-sheet structure in the fibril forms of Aβ and exhibits an increase in fluorescence15. Thus, ThT is often used as a fluorescent marker for fibril forms of Aβ. Large amorphous aggregates partially contain β-sheet structures, which are detected by ThT16. Off-pathway aggregates with β-sheet structures distinct from the on-pathway fibrils have also been reported3. Here, the effect of ATP on the aggregation of Aβ42 was evaluated by monitoring the dynamics of Aβ42. The increased dynamics of Aβ42 may explain the distinct aggregation form induced by ATP.
Results
Dynamic behaviors of Aβ 42
To investigate the dynamic behaviors of Aβ42 in response to the addition of ATP, we obtained DXB measurements to monitor the subnanometer rotational motions of Aβ42. Aβ samples for DXB analysis were prepared based on the method described in a previous study14 with some modifications (Fig. S1A). DXB analyses the dynamical fluctuations of each Aβ42 molecule by calculating the decay constant of the autocorrelation function from the blinking behavior of the diffraction spots (Figs. S1 and S2). Higher decay constant values indicate higher fluctuation of Aβ molecules (Figs. S2 and S3). The addition of ATP resulted in a particularly significant increase in the 0 min decay constant (Fig. 1A), though less than 3 mM ATP showed no significant effect (Table S1). This result corresponded with the knowledge that ATP functions as a hydrotrope to increase the solubility of hydrophobic Aβ429. This effect diminished after 30 min of incubation, suggesting that Aβ42 formed metastable aggregates within 30 min. The median value of the decay constants increased after 1 day of incubation and decreased after 4 days of incubation (Table 1 and Fig. S4). The decreased values may indicate that the aggregates are in a metastable state, as described previously14. The distribution of decay constants was clustered using a Gaussian mixture model, revealing three main components in the distribution (Fig. S5, Table 1). The 2nd component after 1 day of incubation may represent the instability of Aβ42 aggregates, except for that in the presence of ATP.

Box plots of the decay constants. Aβ was incubated at 37 °C in the presence or absence of 10 mM ATP, ADP, or AMP and observed by DXB measurement. The distributions of data in the absence were compared with the absence: ***q < 0.001, **q < 0.01, *q < 0.05, n.s. not significant.
Effect of nucleotides on fibril formation
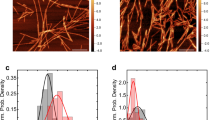
The Aβ monomer was prepared from 26-O-acyliso-Aβ42 (iso-Aβ), which starts aggregating immediately after dissolving in an aqueous solution17. The β-sheet content of the formed aggregates increased for days based on CD spectra (Fig. S6). The fibril formation of iso-Aβ was monitored using the ThT fluorescence assay. ThT fluorescence rapidly increased after 150 min incubation in the absence of ATP (Fig. 2). This result corresponds with the knowledge that nucleus formation during conversion into the fibril form is a rate-limiting step, and the subsequent elongation of fibrils proceeds faster18. The addition of ATP altered the time course of fluorescence and gradually increased the fluorescence. This gentle increase in fluorescence suggests that ATP induces the formation of Aβ42 aggregates with β-sheet structures, similar to polyphenolic flavonoids6. Atomic force microscopy (AFM) analysis revealed that ATP induced the formation of amorphous aggregates with an average diameter of less than 100 nm (Fig. 3) analyzed using ImageJ19. The average diameter of aggregates after 4 days of incubation was smaller in the presence of ATP than in the absence of ATP. Furthermore, ATP seemed to prevent fibril formation since no fibril formation was observed in the tested conditions. The formed amorphous aggregates may be metastable aggregates, including β-sheets, which resulted in the increase in the fluorescence of ThT since amorphous aggregates often partially include β-sheet structures16. ATP seemed to suppress the growth of aggregates until 4 days of incubation, although larger amorphous aggregates with an average diameter of more than 100 nm were observed in the absence after 24 h incubation. Compared to ATP, ADP exhibited a smaller effect on the suppression of fibril formation based on the ThT assay (Fig. 2). AFM images showed fibril formation in the presence of ADP after 24 h (Fig. 3). The AFM images at higher magnification (Fig. S7 and S8) and those in the absence of Aβ (Fig. S8) suggest that these aggregates mainly consist of Aβ.

Fibril formation monitored by ThT. Formation of Aβ aggregates in the presence of 10 mM ATP (Filled triangle), ADP (Open circle) or AMP (Open square) and the absence (Filled circle) were monitored by ThT fluorescence.

AFM images. Aβ aggregates formed in the presence or absence of 10 mM ATP, ADP, or AMP were observed by AFM.
Discussion
ATP can alter the aggregation process of Aβ in the brain since Aβ can accumulate intraneuronally and perform some functions in AD20, although senile plaques mainly consist of extracellular Aβ deposits. The effect of ATP at the early aggregation stage of Aβ42 was demonstrated by the ThT assay, which showed a gradient increase in fluorescence (Fig. 2). This suggests the aggregates formed in the presence of ATP include β-sheet structure which is distinct from those in the other conditions. However, evaluating the aggregation process at the early stage is challenging due to the instability of the aggregates of Aβ42, which has a high tendency to aggregate. DXB monitors the dynamics of Aβ, and the decay constant values decrease as aggregation proceeds based on a previous report14. ATP increased the decay constant values immediately after the addition, suggesting that ATP alters the aggregation process from the early stage. ThT analysis showed that ATP induces aggregates to increase the fluorescence at the early stage. It is plausible that the higher dynamics prevent the formation of aggregates with lower stabilities and, as a result, induce solely stable aggregates. Since various reports have predicted hydrophobic interactions between the adenine moiety of ATP and Aβ21,22, ADP with an adenine moiety should also interact with Aβ to affect the aggregation process. ADP significantly increased Aβ dynamics after 1 day of incubation (Fig. 1 and Fig. S4), and ADP may partially inhibit the on-pathway.
The dynamics increased after 1 day of incubation (Fig. 1 and S4) which is probably due to changes in their formation during the fibrosis progress. CD spectrum suggests that the content of β-sheet structures continues to increase after 1 day of incubation (Fig. S6). FT-IR spectrum showed a distinct peak of spectrum in the presence of ADP after 1 day of incubation from those in the absence (Fig. S9). These results are consistent with the results of the DXB measurements, showing the significantly higher dynamics with ADP as compared to those in the absence (Fig. 1). DXB can detect the rotational fluctuation of Aβ molecules on the sub-nanometer scale. DXB analysis could be a useful method for the evaluation of the inhibitory effect of fibrillar formation by various molecules.
Methods
Sample preparation for DXB
Aβ was dissolved in 0.1% NH4OH/1 mM DMSO to a concentration of 0.1 mg/mL. ATP, ADP, and AMP (10 mM) were added to the Aβ solution and reacted at 37 °C for 0 min, 30 min, 1 day, and 4 days. Aβ solution without reagent as a control was also measured after the same reaction time at 37 °C as when reagent was added.
The immobilization of Aβ molecules onto the substrate film and the labeling of Aβ molecules with gold nanoparticles were performed based on previous studies14. To immobilize Aβ to the polyimide film by amine coupling, 100 μL of NHS/WCS (Dojindo) with minimum essential medium (MEM) buffer was added to the film after KOH exposure. The films were incubated at room temperature (RT) for 10 min. The Aβ solution was dissolved in activation buffer to a concentration of 70 μg/mL. Then, 100 μL of Aβ solution was added to the film and incubated at RT for 30 min. Blocking solution was then added to the film, and the film was washed with phosphate-buffered saline (PBS) buffer after 30 min of RT reaction.
In the 0 min condition, Aβ molecules were immobilized on the substrate film, and each reagent was added and measured immediately. In the 30 min condition, Aβ was immobilized on the substrate film, left to react at 37 °C for 30 min, and then measured. In the 1 day and 4 day conditions, each reagent was added to the Aβ solution, and Aβ was immobilized on the substrate film after reacting at 37 °C for the specified duration.
Gold nanoparticles for labeling Aβ molecules were prepared according to the following procedure. A 60 nm gold nanoparticle solution (Cytodiagnostics Inc.) was centrifuged at 10,000 g for 5 min at 4 ℃. The supernatant solution was then discarded and 50 μL of PBS was added to it. After dissolving well by pipetting, the solution was added to the film on which the Aβ molecules were immobilized.
To label Aβ with gold particles, 100 µL of N-succinimidyl 3-(2-pyridyldithio)propionate (SPDP) crosslinker dissolved in DMSO was added to the Aβ substrate film and reacted at RT for 60 min. Next, 10 mM EDTA with PBS and 0.3 mM dithiothreitol were added to the film and reacted at RT for 30 min to introduce thiol groups to SPDP. Gold nanoparticles (Cytodiagnostics) with a size of 40 nm were centrifuged at 3000×g for 10 min at 4 °C, and the supernatant solution was discarded and concentrated. To bind the gold nanoparticles and SPDP via disulfide bonds, concentrated gold nanosolution was added to the film and reacted for 120 min at RT. The film was then washed with PBS.
Diffracted x-ray blinking
DXB measurements were conducted using the Photon Factory Advanced Ring AR-NW12 beamline. Time-resolved diffraction images were recorded using a 2D photon-counting detector (PILATUS 2 M, Dectris, Switzerland). Measurements were performed at room temperature. X-ray diffraction images were taken in a series of 2010 images with an exposure time of 50 ms. Time profiles of Au(111) intensity changes were extracted from these images using ImageJ. The time-resolved diffraction intensity in Au(111) was analyzed at each pixel by the autocorrelation function as follows:
where I(t) is the diffraction intensity, the brackets < > indicate the time-averaged value, and τ is the lag time. The ACF curves were fitted to an exponential curve by ACF(t) = Aexp(−Гt) + y, where A is the amplitude, y is the conversion factor, and Г is the decay constant. The decay constants were chosen to satisfy the following conditions: (i) 0 < y, 0 < A and 0 < Г, and (ii) residual values between the fitted and actual ACF curves of less than 1.023,24. These calculations were performed for all pixels. The distribution of the decay constants of Au (111) diffraction was visualized using histograms and box plots to estimate the dynamic behavior of the protein molecules (Fig. S1C).
Because the distributions of the decay constant did not show a Gaussian distribution, the distributions were statistically analyzed with the nonparametric Wilcoxon rank sum test.
To evaluate whether the distribution of decay constants contains multiple distribution components, we clustered the distribution components by a Gaussian mixture model with the EM algorithm. The analysis was performed using R scripts. Based on the likelihood values, the distributions of decay constants were classified into three Gaussian components (Fig. S5).
Evaluation of fibril formation
The Aβ monomer was prepared by converting iso-Aβ (Peptide Institute, Inc., Osaka, Japan) in phosphate-buffered saline (PBS) consisting of 137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, and 2 mM KH2PO417. The Aβ monomer was preserved in dimethyl sulfoxide (DMSO) prior to use since the conversion proceeded in a few minutes. The aggregation proceeded in a final concentration of 10% v/v DMSO where the structure of Aβ1–40 is little affected25. The β-sheet structures on the surface of the assemblies of Aβ42 were analyzed by the ThT assay26. The fluorescence of 10 μM ThT in the presence or absence of 2.2 μM Aβ42 was monitored at an excitation wavelength of 444 nm and emission wavelength of 485 nm. The solutions were monitored every 30 min and shaken for 5 s prior to each measurement.
Evaluation of aggregate shape
The shapes and sizes of the assemblies were observed by AFM. The solution of the assemblies (10 μL) of 0.1 mg/mL Aβ42 was dripped and spread onto fresh mica and dried by vacuum drying for a few minutes. Measurements were performed in alternating current (AC) mode at room temperature. The sample was measured by a typical resonance frequency of approximately 190 kHz and a spring constant of 4.5 N/m. The sizes of more than 6 assemblies were measured, and the mean and standard deviation were determined.
Data availability
The raw data for Figs. 1, S4, and S5 has been deposited in Figshare repository, https://doi.org/10.6084/m9.figshare.25230083. The data of Figs. 2, S6 and S9, are publicly available in Figshare repository, https://doi.org/10.6084/m9.figshare.25218485. The data that support the findings of this study are available from the corresponding author on reasonable request.
References
Yamaguchi, H. et al. Electron micrograph of diffuse plaques. Initial stage of senile plaque formation in the Alzheimer brain. Am. J. Pathol. 135, 593–597 (1989).
Du, W. J. et al. Brazilin inhibits amyloid β-protein fibrillogenesis, remodels amyloid fibrils and reduces amyloid cytotoxicity. Sci. Rep. 5, 7992 (2015).
Yasumoto, T. et al. High molecular weight amyloid β1–42 oligomers induce neurotoxicity via plasma membrane damage. FASEB J. 33, 9220–9234 (2019).
Mrdenovic, D. et al. Amyloid β interaction with model cell membranes—What are the toxicity-defining properties of amyloid β?. Int. J. Biol. Macromol. 200, 520–531 (2022).
Ehrnhoefer, D. E. et al. EGCG redirects amyloidogenic polypeptides into unstructured, off-pathway oligomers. Nat. Struct. Mol. Biol. 15, 558–566 (2008).
Gargari, S. A. & Barzegar, A. Simulations on the dual effects of flavonoids as suppressors of Aβ42 fibrillogenesis and destabilizers of mature fibrils. Sci. Rep. 10, 16636 (2020).
Wong, H. E. et al. A safe, blood-brain barrier permeable triphenylmethane dye inhibits amyloid-β neurotoxicity by generating nontoxic aggregates. ACS Chem. Neurosci. 2, 645–657 (2011).
Yang, A. et al. Attenuation of β-amyloid toxicity in vitro and in vivo by accelerated aggregation. Neurosci. Bull. 33, 405–412 (2017).
Patel, A. et al. ATP as a biological hydrotrope. Science 356, 753–756 (2017).
Coskuner, O. & Murray, I. V. J. Adenosine triphosphate (ATP) reduces amyloid-β protein misfolding in vitro. J. Alzheimers. Dis. 41, 561–574 (2014).
Kurisaki, I. & Tanaka, S. ATP converts Aβ42 oligomer into off-pathway species by making contact with its backbone atoms using hydrophobic adenosine. J. Phys. Chem. B 123, 9922–9933 (2019).
Sims, N. R. et al. Metabolic processes in Alzheimer’s disease: Adenine nucleotide content and production of 14CO2 from [U-14C] glucose in vitro in human neocortex. J. Neurochem. 41, 1329–1334 (1983).
Sekiguchi, H. et al. Diffracted X-ray blinking tracks single protein motions. Sci. Rep. 8, 17090 (2018).
Chang, J. et al. Dynamic observations of various oligomers in amyloid β isoforms using laboratory diffracted X-ray blinking. Biochem. Biophys. Rep. 31, 101298 (2022).
Biancalana, M. & Koide, S. Molecular mechanism of Thioflavin-T binding to amyloid fibrils. Biochim. Biophys. Acta 1804, 1405–1412 (2010).
Herzberg, M. et al. Probing the secondary structure of individual Aβ40 amorphous aggregates and fibrils by AFM-IR spectroscopy. Chembiochem 21, 3521–3524 (2020).
Sohma, Y. et al. Design and synthesis of a novel water-soluble Aβ1-42 isopeptide: An efficient strategy for the preparation of Alzheimer’s disease-related peptide, Aβ1-42, via O-N intramolecular acyl migration reaction. Tetrahedron Lett. 45, 5965–5968 (2004).
Faller, P. et al. Role of metal ions in the self-assembly of the Alzheimer’s amyloid-β peptide. Inorg. Chem. 52, 12193–12206 (2013).
Schneider, C. A., Rasband, W. S. & Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675 (2012).
LaFerla, F. M. & Green, K. N. Intracellular amyloid-β in Alzheimer’s disease. Nat. Rev. Neurosci. 1, 19 (2012).
Dec, R., Puławski, W. & Dzwolak, W. Selective and stoichiometric incorporation of ATP by self-assembling amyloid fibrils. J. Mater. Chem. 9, 8626–8630 (2021).
Pal, S. & Paul, S. ATP controls the aggregation of Aβ16−22 peptides. J. Phys. Chem. B 124, 210–223 (2020).
Mio, K. et al. X-ray-based living-cell motion analysis of individual serotonin receptors. Biochem. Biophys. Res. Commun. 529, 306–313 (2020).
Kuramochi, M. et al. Dynamic motions of ice-binding proteins in living Caenorhabditis elegans using diffracted X-ray blinking and tracking. Biochem. Biophys. Rep. 29, 101224 (2022).
Garzon-Rodriguez, G., Sepulveda-Becerra, M., Milton, S. & Glabe, C. G. Soluble amyloid Abeta-(1–40) exists as a stable dimer at low concentrations. J. Biol. Chem. 272, 21037–21044 (1997).
Amdursky, N., Erez, Y. & Huppert, D. Molecular rotors: What lies behind the high sensitivity of the thioflavin-T fluorescent marker. Acc. Chem. Res. 45, 1548–1547 (2012).
Acknowledgements
We are grateful to the staff of the Photon Factory for their assistance in data collection, which was approved by the Photon Factory Program Advisory Committee.
We thank M. Hikita for help with the DXB experiment setup in the Photon Factory Advanced Ring (PF-AR). This work was supported by JST ACT-X JPMJAX22B7, the Top Runners in Strategy of Transborder Advanced Researches (TRiSTAR) program conducted as the Strategic Professional Development Program for Young Researchers by the MEXT, and Foundation for Interaction in Science & Technology, and Nakatani Foundation for Advancement of Measuring Technologies in Biomedical Engineering. Experiments were also performed with the approval of the, AR-NW12A of the PF-AR with the approval of KEK (Proposal No. 2023G132).
Author information
Authors and Affiliations
Contributions
M.K. and K.Y. (Nihon Univ.) wrote the main manuscript text, M.K. and H.T. prepared Figs. 1, S1, S2, S3, S4 and S5 and Table 1 and S1, K.Y. (Nihon Univ.), M.N. and T.K. prepared Figs. 2, 3, S7, S8, and S9., K.Y. (Nihon Univ.), T.T., N.P., and K.Y. (Kyoto Univ.) prepared Figure S6. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kuramochi, M., Nakamura, M., Takahashi, H. et al. Adenosine triphosphate induces amorphous aggregation of amyloid β by increasing Aβ dynamics. Sci Rep 14, 8134 (2024). https://doi.org/10.1038/s41598-024-58773-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-58773-6
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
