
Abstract
Drosophila melanogaster is unique among animal models because it has a fully defined synthetic diet available to study nutrient-gene interactions. However, use of this diet is limited to adult studies due to impaired larval development and survival. Here, we provide an adjusted formula that reduces the developmental period, restores fat levels, enhances body mass, and fully rescues survivorship without compromise to adult lifespan. To demonstrate an application of this formula, we explored pre-adult diet compositions of therapeutic potential in a model of an inherited metabolic disorder affecting the metabolism of branched-chain amino acids. We reveal rapid, specific, and predictable nutrient effects on the disease state consistent with observations from mouse and patient studies. Together, our diet provides a powerful means with which to examine the interplay between diet and metabolism across all life stages in an animal model.
Similar content being viewed by others


Introduction
Nutrition is a major environmental factor that affects animal physiology, metabolism, and influences traits such as survival and fitness1. Understanding how each nutrient contributes to the animal phenotype remains a major challenge in biology due to the complex composition of food. Animal models such as the vinegar fly Drosophila melanogaster permit wide-scale testing of genotype-environment interactions and their molecular basis. These attributes are key in medical and biological sciences due to the shared physiological and metabolic processes in flies and humans2,3,4,5.
D. melanogaster is unique amongst animal models as it has a fully synthetic chemically-defined (holidic) medium allowing for manipulation of individual nutrients6. This diet, named hereon as 100N, has been used extensively to test how nutrients impact complex adult traits such as reproduction, lifespan, behaviour, and host-microbiome interactions6,7,8,9. The annotation “100N” describes our standard (100) diet, which contains the minimal amount of high-quality dietary nitrogen (N) to support maximal adult fecundity and longevity7. However, a major limitation of 100N is that flies reared solely on it (i) are significantly delayed when compared to flies reared on sugar-yeast (SY) diet (100N: 17.95 days to eclosion; SY: 10.55 days at 25 °C), (ii) show reduced survivorship (100N: 85%; SY: 96%), and (iii) are smaller in size upon eclosion (Fig. 1a, Figure S1a), suggesting that 100N is suboptimal for growth and development. To yield a formula suitable for experiments in pre-adult stages we tested a series of supplementations involving amino acids (aa), lipids, carbohydrates, and other metabolites such as carnitine and acetic acid as suggested by metabolome profiles. Here we show that by adding 8% extra total amino acid content and doubling the amount of acetic acid in the original formula, developmental timing is reduced, egg-to-adult survivorship and fat levels in the fat body are restored, and adult body mass upon eclosure is enhanced, without compromising adult lifespan. We further demonstrate an application of the improved formula together with a D. melanogaster model of an inherited metabolic disorder for identifying diets of therapeutic potential.

Optimizing the amino acid content of 100N for Drosophila pre-adulthood. (a), Eclosion kinetics, median egg-to-adult duration in days after-egg-lay (dAEL), mean percentage egg-to-adult survivorship, and male weight upon eclosure (mg) on SY and 100N diets. (b–d), Egg-to-adult duration (dAEL) and egg-to-adult survivorship on 100N supplemented with (b) 0.1%; 0.5%; or 1% of yeast extract (YE), (c) 50% Ser; 50% Gly; 100% Glu; 50% supplementation of all 20 aa (150N); and (d) 50% Ser, Val and Gly, 50% Gly and Ser, 110% Glu, and 8% supplementation of all 20 aa (108N). (a–d), five biological replicates per group of 20 individuals each. For adult male weight upon eclosure at least 30 individuals were weighted per group. (a), Student's unpaired t-test, ****p < 0.0001, *p < 0.05. b-d, One-way ANOVA followed by Tukey’s HSD test; different letters (A, B and C) represent statistically significant differences (P < 0.05).
Results and discussion
To improve the growth and development of D. melanogaster reared from embryo to adulthood on the holidic diet, we made a series of tests varying individual nutrients. As a starting point, before investigating which specific nutrients could be limiting, we examined the effects of a complex mixture, water soluble yeast extract (YE). Yeast is a common component of fly media and vinegar flies are naturally attracted to it10. We enquired whether a component present in YE could improve the development time of flies reared on 100N. Adding 0.1% YE improved development time by 1.5-days, but while further additions progressively improved development time, they also reduced survival indicating nutritional imbalance (Fig. 1b, Figure S1b). YE is a rich source of amino acids (aa), which promote growth and development in many ways. Among these aa, Glutamate (Glu) is extensively utilized for protein synthesis and plays a critical role in central carbon and nitrogen metabolism11. Glycine (Gly) and serine (Ser), besides their protein synthesis function, are vital for cellular redox control and are the major source of one-carbon units, which is essential for various anabolic pathways including nucleotide synthesis12. Branched-chain amino acids (BCAAs), such as valine (Val), play pivotal roles in growth regulation via the activation of the growth-stimulating target of rapamycin (TOR) pathway13,14, as well as in muscle function and immunity15. Thus, we investigated whether individual supplementation of these four amino acids could confer similar benefits without compromising survival. Individually adding 50% more (in comparison to 100N) Ser, Gly, or Glu showed trends but no significant reduction in development time (Fig. 1c, Figure S1c). The only individual aa supplementation tested to produce a significant reduction in development time from egg to adult and without compromising survivorship was Val (Fig. 1c, Figure S1c). As a control we also tested the effect of supplementing 50% of all aa (150N). That, however, was highly detrimental to survival and development rate (Fig. 1c, Figure S1c). Given the positive trends observed with some of the four aa individual supplementations we next investigated whether supplementing the four in different combinations could improve development time (Fig.1d, Figure S1d). Increasing the content of both Gly and Ser by 50%, or even the content of Ser, Val and Gly by 50%, caused no significant improvement. Given Glu is an important nitrogen source, we tested whether 110% extra Glu (equal to the nitrogen content of supplementing 50% extra Ser, Val and Gly), would improve development rate, but again had no effect, suggesting the development delay is not caused by lack of nitrogen. However, as a control diet, an addition of 8% more of all 20 amino acids (108N; equal to the nitrogen content of supplementing 50% extra Ser, Val and Gly) promoted a reduction in egg-to-adult development time by 1.7-days and significantly increased survivorship (98%, Fig. 1d, Figure S1d).
To further improve the formula, we investigated the differences between 108N- and SY-raised Drosophila 3rd instar larvae via untargeted metabolomics (Fig. 2, Table S1,2). We identified 1065 metabolites with a high degree of confidence and found 674 that were significantly different between the two diets. In particular, lipid-, carnitine-, energy metabolism- (Fig. 2b,c), and β-oxidation-associated metabolites (Fig. 2d–f) were all lower in the 108N-raised larvae, suggesting the developmental delay may be due to an energy deficit, possibly correlated to fatty acid (FA) metabolism. Carnitines are involved in the transport of long-chain FA into mitochondria for energy production via β-oxidation16. We therefore attempted supplementing the 108N diet with carnitine, coconut oil (mixture rich in FA species), linolenic acid (polyunsaturated FA) and sucrose (carbon source). None of these additions improved larval performance on the 108N diet, and in many cases proved to be toxic (Figure S2a-h). These results indicate that providing energy-rich metabolites to correct the deficit suggested in the metabolic profile does not enhance the development rate. It is possible that other nutrients remain limiting and/or that fly metabolism relies on nutrients other than sugar and fats as an energy source.

Metabolic profile comparison of 3rd instar larvae reared on SY or 108N. (a), Principal Component Analysis (PCA) of metabolic profiles of third instar larvae reared on SY or 108N. For metabolome analysis a total of five biological replicates were performed per group, each replicate containing 10 five-day-old larvae. (b,c) Log2(fold change) of larval metabolites involved in (b) long-chain FA transport in mitochondria, and (c) energy metabolism significantly affected in 108N in comparison to SY media. Student's unpaired t-test, ****p < 0.0001. (d), Top 50 metabolites, based on fold change, significantly affected by dietary treatment (Student's unpaired t-test, P < 0.05). The cell color represents the z-score, i.e., the standardized scores on the same scale, calculated dividing a score's deviation by the standard deviation in the row. The features are color-coded by row with red indicating high intensity and blue indicating low intensity. (e,f), Enrichment analysis showing the top 20 pathways (based on small molecule pathway database—SMPDB) (e) increased or (f) decreased in larvae reared on 108N in comparison to larvae reared on SY.
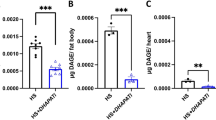
The natural substrate of D. melanogaster larvae is fermenting fruit which is poor in long-chain fatty acids but rich in alcohol and acetate, a short chain FA used in the synthesis of acetyl-CoA17,18. We therefore tested acetic acid (AcOH) supplementation. Doubling the original content of AcOH in the holidic diet (from 0.3% v/v to 0.6% v/v) significantly reduced egg-to-adult time by 1-day compared to 108N, without compromise to survival (Fig. 3a, Figure S2i). Tissue analysis of animals reared on 108N (0.6% AcOH) showed larger adipose tissue (fat body) lipid stores in comparison to 100N-raised larvae, reaching equivalent levels to SY-raised larvae, indicating a general metabolic improvement (Fig. 3b,c). These individuals also attained significantly higher bodyweights as newly-eclosed adults, compared to 100N-raised animals (Fig. 3d), despite reaching adulthood in a shorter time. Importantly, the lifespan of adult flies that had been raised on 108N (0.6% AcOH) was no different to those raised on SY (Fig. 3e). Overall, when compared to the original holidic diet, our adjusted formula improved survivorship to 98% (equivalent to SY-raised D. melanogaster), reduced egg-to-adult time by 2.7 days (Fig. 3f), restored fat levels, and enhanced body mass, without compromising adult lifespan. Adult weight equivalent to SY-raised flies is not reached on 108N (0.6% AcOH), suggesting there is room for future nutritional improvements. This could involve the addition or modulation of vitamins and salts. It also remains possible that we have reached the limit of the synthetic diet environment to yield optimal fly development.

Optimizing the 108N diet for D. melanogaster pre-adulthood. (a), Egg-to-adult duration (dAEL) and egg-to-adult survivorship on 108N containing 0.45%; 0.6%; or 0.75% acetic acid (AcOH, v/v). (b), Lipid storage in the late-larval fat body on 100N, 108N (0.6% AcOH), or SY. Nucleic acids (blue) and lipid droplets (red), 400 × magnification. (c), Percentage of fat body area occupied by lipid droplets. (d), Adult male weight upon eclosion (mg) raised on SY, 100N, or 108N (0.6% AcOH). (e), Lifespan curves of adult flies fed SY food after being raised on either 108N (0.6% AcOH) or SY during their pre-adult period. (f), Eclosion kinetics, egg-to-adult duration (dAEL) and egg-to-adult survivorship on SY, 100N, and 108N (0.6% AcOH) diets. (a) and (f), Five biological replicates per group of 20 individuals each. (c), Five larvae per group (each point represents an average of 5 individual measurements per larva). (d), For adult male weight upon eclosure at least 30 individuals were weighed per group. (e), 5 to 10 replicates per group of 10 female adults each. (a), (c), (d), and (f), One-way ANOVA followed by Tukey’s HSD test; different letters (A, B and C) represent statistically significant differences (p < 0.05). i, Cox Proportional-Hazards modelling (p = 0.472). Data shown in Fig. 1a are repeated in Fig. 3d and f, to compare against 108N (0.6% AcOH).
A synthetic diet on which D. melanogaster can be raised is particularly valuable in the modelling of human metabolic disorders, where phenotypic abnormalities often manifest prior to adulthood19. For example, human disorders such as Phenylketonuria manifest with rapid neurological decline prior to adulthood mitigated by a custom diet with single aa restriction that drastically improves health outcomes20. However, the majority of metabolic disorders lack therapeutic diets21. To demonstrate the potential of our diet as a tool to find new dietary treatments, we applied it in a model of human Branched Chain Amino Acid (BCAA) Transaminase (BCAT2) deficiency, a rare disorder associated with developmental delay, intellectual disability, seizures, and movement defects21. BCAT2 catalyzes the first step in Valine (Val), Leucine (Leu) and Isoleucine (Ile) catabolism22 and patients have high plasma and tissue levels of these aa21 (Fig. 4a).

Applying the pre-adult-optimized synthetic diet in a Drosophila inherited metabolic disorder model. (a), Schematic of branched-chain amino acid (BCAA) metabolism. BCAA are converted into branched-chain α-keto acid (BCKA) by the action of BCAT2, the deficiency of which causes a rare metabolic disorder. (b), Schematic of BcatTG4 allele in D. melanogaster containing the disruption T2A-GAL4 (TG4) cassette. (c), Validation of the BcatTG4 allele, showing Bcat expression is not detected. w1118 are a wildtype control genotype. (d), Egg-to-adult survivorship of BcatTG4 flies and BcatTG4 expressing the human orthologue Bcat2, when reared on 108N (0.6% AcOH). (e), BcatTG4 survivorship contour plots resulting from a 3-way nutrient (Leu, Ile, Val) dose response array (27 diets total, 8–9 replicates performed per diet). (f), Egg-to-adult survivorship of BcatTG4 flies raised on SY, 108N (0.6% AcOH), and 108N (0.6% AcOH) with 50% BCAA content. (d) and (f), five biological replicates per group of 20 individuals each. (c), Student's unpaired t-test, ****p < 0.0001. (d) and (f), One-way ANOVA followed by Tukey’s HSD test. Different letters (A, B and C) represent statistically significant differences (p < 0.001).
We used a D. melanogaster Bcat allele (BcatTG4) that comprises a T2A-GAL4 cassette inserted within the second intron of Bcat, which abolishes expression of the full-length transcript and truncates protein translation23 (Fig. 4b,c). Egg-to-adult survival for BcatTG4 homozygotes on 108N (0.6% AcOH) is 4% and this was rescued to 60% via expression of a human ortholog BCAT2 transgene suggesting fly and human genes are functionally equivalent (Fig. 4d). We next tested BcatTG4 flies and their survival relative to sibling controls in a nutrigenomic array of 27 BCAA-modified diets where each BCAA (Leu, Ile, and Val) was co-varied across three concentrations (50%, 75% and 100%). BcatTG4 flies had highest relative survival when reared on a diet where all three BCAAs were reduced by 50%, while no benefit was observed from restricting one or two BCAA at a time in any combination (Fig. 4e, Figure S3). Reducing Val levels to 50% whilst keeping Leu and Ile at 100% had a drastic negative impact in median egg-to-adult duration, suggesting that maintaining a BCAA balance appears important for development (Figure S3). Tracking BcatTG4 flies developing in isolation on the 50% all BCAA diet (to determine absolute survival, rather than relative to genetic controls) revealed a striking improvement in survivorship (from ~ 5% to 34%, Fig. 4f). These findings mirror diet responses observed in BCAT2 deficiency patients21 and observations from a mouse BCAT2 model24. This highlights the potential of our modified synthetic diet for nutrigenomic studies of fly disease models for which effective diet formulations are lacking. More broadly, our diet expands the study of nutrient-gene interactions to the entire life cycle of Drosophila, thus permitting the exploration of even complex processes such as growth and development.
Materials and methods
Fly stocks and base diet
The synthetic (holidic) diet optimizing experiments were performed on w1118 (Bloomington stock #3605). For experiments involving Bcat mutant flies, BcatTG4 mutants (Bloomington stock #79238) were first outcrossed to w1118 for removal of y1 and then maintained over fluorescently marked balancer chromosome (FM7a, ChFP)25. Rescue with human ortholog was performed by cross with UAS-hBCAT2 (Bloomington stock #84835). BcatTG4/ FM7a, ChFP females were crossed to FM7a, ChFP/Y; UAS-hBCAT2/UAS-hBCAT2 males. First instar larvae without ChFP maker were selected, corresponding to hemizygous males BcatTG4/Y; UAS-hBCAT2/ + . The viability of hemizygous BcatTG4 males containing one copy of UAS-hBCAT2 was then compared to that of hemizygous BcatTG4 males without the UAS-hBCAT2 transgene. Dietary manipulations were conducted using the synthetic diet6,7 (Table S3). The sugar-yeast (SY) diet was prepared as per standard media (Table S4). All stocks of Drosophila melanogaster were maintained at 18 °C on sugar-yeast medium under natural photoperiod conditions. For experiments, flies were reared in incubators at 25 °C and 45% humidity in specific diets as described below.
Testing holidic diet improvements
The modifications tested with the holidic diet include manipulation of amino acid levels, supplementation with yeast extract (Bacto™ Yeast Extract, Thermo Fisher Scientific), sucrose (Sigma-Aldrich, S1888), organic virgin coconut oil (Macro Wholefoods Market), Linolenic acid (L2376-500MG, Merck), L-Carnitine hydrochloride (Merck, C0283-5G), and acetic acid (Fisher, A/0400/PB15). The solutions of yeast extract, sucrose, acetic acid, and carnitine were prepared in Milli-Q water. Appropriate volumes of these solutions, with the exception of Linolenic acid and coconut oil, were added to the solid food surface and left for 24 h in a 25 °C incubator to allow for complete diffusion. 300 µL was added to each vial as we found that this is the maximum volume of liquid that would be fully absorbed and diffuse into the solidified holidic diet within a 24 h period and allowed for concentrations where amino acids remained soluble. For Linolenic acid and coconut oil, appropriate volumes were added to the holidic diet prior to setting and homogenized with help of a magnetic stirrer before dispensing into vials. Diets with varied amino acid concentrations were prepared by modifying the base holidic diet recipe (Table S3).
Survival and developmental timing
Embryos were collected from population cages containing approximately 200 females and 50 males (5 to 10 days old) laying for 6 h on apple juice agar plates supplemented with yeast paste. Embryos were then washed with distilled water and transferred to a new plate not containing yeast paste and left for 24 h to hatch. Twenty first instar larvae were transferred to each vial for the different diets. For each diet, at least five replicate vials were tested (n = 100 individuals). Vials were monitored every 12 h for a period of 20 days and the numbers of pupae and adults were recorded. For creating the pupariation kinetics or eclosion kinetics curves, data were visualized as the cumulative percentage of individuals at each life stage as a function of time (days) from egg lay. Box plots depicting each biological replicate data point were also generated to represent the median time of development (in days) and mean survival (in percentage) for egg-to-pupa and egg-to adult stages. Significant differences in median time of development and mean survival were analyzed using multiple comparison one-way ANOVA followed by Tukey’s HSD test, where each group was compared against each other group within an experiment (GraphPad Prism 9). Groups with different letters (“A”, “B”, etc.) indicate they are significantly different from one another, while groups with the same letter are indistinguishable (p-values are shown in Figure legends).
Metabolic profiling
Five replicates of 10 five-day-old larvae from each genotype and diet condition were collected, washed in PBS, blotted dy, and weighed. Samples were then transferred to 1.5 mL safe lock microtubes tubes (Eppendorf), flash frozen in liquid nitrogen, and stored at -80 °C. Thawed larvae were homogenized using a disposable pestle in 20 µL of ice-cold extraction solvent consisting of 2:6:1 chloroform:methanol:water with 2 µM of (CHAPS, CAPS, PIPES and TRIS) acting as internal standards. Once ground additional solvent was added to a final ratio of 20 µL of extraction solvent per mg of larvae. Samples were vortexed for 30 s and then sonicated in an ice-water bath for 10 min and centrifuged at 4 °C (22,000 × g for 10 min). The supernatant was then transferred to a glass vial for LC–MS based metabolomic analysis. 20 µL of each extract were combined to make a pooled quality control (PQC) sample.
LC–MS was performed using a Dionex Ultimate 3000 UHPLC coupled to an QExactive Plus mass spectrometer (Thermo Scientific). Samples were analyzed by hydrophilic interaction liquid chromatography (HILIC) following a previously published method26. The chromatography utilized a ZIC-pHILIC column 5 µm, 150 × 4.6 mm with a 20 × 2.1 mm ZIC-pHILIC guard column (both Merck Millipore, Australia) (25 °C). A gradient elution of 20 mM ammonium carbonate (A) and acetonitrile (B) (linear gradient time-%B: 0 min-80%, 15 min-50%, 18 min-5%, 21 min-5%, 24 min-80%, 32 min-80%) was utilized. Flow rate was maintained at 300 μL/min. Samples were stored in the autosampler (6 °C) and 10 μL was injected for analysis. MS was performed at 70,000 resolution operating in rapid switching positive (4 kV) and negative (− 3.5 kV) mode electrospray ionization (capillary temperature 300 °C; sheath gas flow rate 50; auxiliary gas flow rate 20; sweep gas 0; probe temp 120 °C). Samples were randomized and processed in a single batch with intermittent analysis of PQC samples to ensure reproducibility. For accurate metabolite identification, a standard library of ~ 500 metabolites was analyzed before sample testing and accurate retention time for each standard was recorded. This standard library also forms the basis of a retention time prediction model used to provide putative identification of metabolites not contained within the standard library27.
Metabolic profiling data analysis
Acquired LC–MS/MS data was processed in an untargeted fashion using open source software IDEOM, which initially used msConvert (ProteoWizard)28 to convert raw LC–MS files to mzXML format and XCMS to pick peaks to convert to .peakML files29. Mzmatch was subsequently used for sample alignment and filtering30. Metabolites were identified based on accurate mass (< 2 ppm) and comparison of their retention time against that determined for compounds in the standard library or predicted on the basis of their physiochemical characteristics27. Only metabolites that were identified with a level of confidence equal to or greater than 6 in IDEOM were used for downstream functional and statistical analyses, using MetaboAnalyst 5.031. Global metabolic variations due to genotype and diet were visualized using PCA. One-way ANOVA [false discovery rates (FDR) < 0.05] was used to identify significant changes in metabolite levels. A heatmap was created to visualize the 50 topmost affected metabolites (based on fold change).
Assessment of lipid environment by confocal microscopy
The effects of SY, 100N and 108N supplemented with 0.6% AcOH on the lipid storage of wandering third instar larvae were assessed using fluorescence microscopy. Fat bodies were dissected in PBS (Ambion) and subjected to lipid staining with Nile Red N3013 Technical grade (Sigma-Aldrich) and nucleic acid staining with DAPI (Sigma-Aldrich). Tissues were fixed in 4% PFA (Electron Microscopy Science) for 20 min and stained with 0.5 µg/mL Nile Red and 10 µg/mL DAPI in PBS + 0.1% Triton X-100 for 15 min. After staining, samples were washed in PBS and slide were mounted in Vectashield (Vector Laboratories). Microscopies were performed in CV1000 Disc Confocal Microscope (Olympus Life Science) at 400 × magnification. Images were analyzed using ImageJ software. The percentage of area occupied by lipid droplets per fat body samples was averaged from three independent measurements of randomly chosen sections of 2500 µm2. A total of five biological replicates were assessed for each dietary condition, each replicate consisting of a single tissue from a single larva. Results were analyzed using One-way ANOVA followed by Tukey’s HSD test (GraphPad Prism 9).
Male weight upon eclosure
The adult weight upon eclosure of male flies reared on SY, 100N, or 108N + 0.6% AcOH was assessed using a SE2 Ultramicrobalance (Sartorius, Goettingen, Germany). At least 30 flies were weighed upon eclosure. Results were analyzed using One-way ANOVA followed by Tukey’s HSD test (GraphPad Prism 9).
Adult lifespan
The survival of flies reared from egg-to-adult on 108N (0.6% AcOH) was compared to those reared on the SY diet. Upon eclosion, female adults from either diet were transferred onto SY diet and survivorship was checked every other day, at the same time when flies were transferred to fresh media. Five to ten replicates per group of ten flies each were performed. Fly survival was recorded using the software DLife32 and Cox Proportional-Hazards modelling was used for analysis (R, package survival). Difference in survival between groups was analysed using a linear model (base R, “lm”) and post-hoc comparisons from the emmeans package.
BCAA-modified diet array
Large numbers of developmentally synchronized embryos obtained from multiple cages containing BcatTG4/FM7a, ChFP flies were collected and washed in distilled water, and transferred to 15 mL tubes containing PBS. Once the embryos had settled (1 min), excess PBS was removed. Using a micropipette set to 5 µL and a tip cut to widen the bore, small volumes of embryos were aspirated and then dispensed into vials onto the medium surface. This was performed for each of the 27 modified synthetic diets tested with varied levels (100%, 75% and 50%) of leucine, isoleucine and valine replicated at least 8 times (Table S4). Vials were checked daily for 30 days and the total number of adult flies were counted with the survival proportion of BcatTG4 (BcatTG4/ total flies) calculated. The linear mixed model (R package: lme4) was fitted to analyse the effects of diet, genotype, and their interaction on mean BcatTG4 survival proportion (number of BcatTG4 flies out of total), with replicates treated as a random effect.
Real time PCR
The expression of Bcat was measured in BcatTG4 homozygous flies and w1118 control flies. A total of five whole flies were used for a single biological replicate and three replicates were taken from each genotype. RNA isolation with TriSure (Bioline) was performed following the manufacturer’s instructions. RNA purity and concentration were evaluated by spectrophotometry (NanoDrop® ND-1000, NanoDrop Technologies). RNA samples were then diluted to 1 µg/µL and stored at − 80 °C. cDNA was generated using the Superscript III reverse transcriptase (NEB). RT-qPCR analysis was performed in a 7500 Real Time PCR System (Applied Biosystems) using the SYBR® Green Reagent protocol (Applied Biosystems) and the following pairs of primers: Bcat Forward 5’ CCGCCTATTCACCGATCACA3’; Bcat Reverse 5’ TACGCCTTCATGCCCTCAAA3’; RpL11 5’ CGATCCCTCCATCGGTATCT3’; RpL11 Reverse 5’ AACCACTTCATGGCATCCTC3’. Relative expression was calculated using the 2^(-∆∆Ct) method and RpL11 expression as a housekeeping gene. The results were analyzed by the Shapiro–Wilk normality test (p > 0.05) to test for normality, before comparing the transcript levels between the two treatment groups using two-tailed Student's t-tests (p < 0.05 (GraphPad Prism 9).
Data availability
All data generated or analysed during this study are included in this published article and its supplementary information files except for the raw metabolomics dataset which can be found at the MetaboLights repository upon publication, www.ebi.ac.uk/metabolights/MTBLS9556.
References
Simpson, S. J. & Raubenheimer, D. The Nature of Nutrition A Unifying Framework from Animal Adaptation to Human Obesity (Princeton University Press, 2012).
Chatterjee, N. & Perrimon, N. What fuels the fly: Energy metabolism in Drosophila and its application to the study of obesity and diabetes. Sci. Adv. 7, 1–15 (2021).
Kim, S. K., Tsao, D. D., Suh, G. S. B. & Miguel-Aliaga, I. Discovering signaling mechanisms governing metabolism and metabolic diseases with Drosophila. Cell Metab. 33, 1279–1292 (2021).
Shin, M., Copeland, J. M. & Venton, B. J. Drosophila as a model system for neurotransmitter measurements. ACS Chem. Neurosci. 9, 1872–1883 (2018).
Ugur, B., Chen, K. & Bellen, H. J. Drosophila tools and assays for the study of human diseases. Dis. Model. Mech. 9, 235–244 (2016).
Piper, M. D. W. et al. A holidic medium for Drosophila melanogaster. Nat. Methods 11, 100–105 (2014).
Piper, M. D. W. et al. Matching dietary amino acid balance to the in silico-translated exome optimizes growth and reproduction without cost to lifespan. Cell Metab. 25, 610–621 (2017).
Leitão-Gonçalves, R. et al. Commensal bacteria and essential amino acids control food choice behavior and reproduction. PLOS Biol. 15, e2000862 (2017).
Henriques, S. F. et al. Metabolic cross-feeding in imbalanced diets allows gut microbes to improve reproduction and alter host behaviour. Nat. Commun. 11, 4236 (2020).
Becher, P. G. et al. Chemical signaling and insect attraction is a conserved trait in yeasts. Ecol. Evol. 8, 2962–2974 (2018).
Yelamanchi, S. D. et al. A pathway map of glutamate metabolism. J. Cell Commun. Signal. 10, 69–75 (2016).
Ducker, G. S. & Rabinowitz, J. D. One-carbon metabolism in health and disease. Cell Metab. 25, 27–42 (2017).
Zhang, S. et al. Regulation of mTORC1 by amino acids in mammalian cells: A general picture of recent advances. Anim. Nutr. 7, 1009–1023 (2021).
Blank, H. M. et al. Branched-chain amino acid synthesis is coupled to TOR activation early in the cell cycle in yeast. EMBO Rep. 24, 1–14 (2023).
Zhang, S., Zeng, X., Ren, M., Mao, X. & Qiao, S. Novel metabolic and physiological functions of branched chain amino acids: a review. J. Anim. Sci. Biotechnol. 8, 10 (2017).
Flanagan, J. L., Simmons, P. A., Vehige, J., Willcox, M. D. & Garrett, Q. 2010 Role of carnitine in disease. Nutr. Metab. 7, 30 (2010).
Carvalho, M. et al. Effects of diet and development on the Drosophila lipidome. Mol. Syst. Biol. 8, 600 (2012).
McKenzie, J. A. & McKechnie, S. W. A comparative study of resource utilization in natural populations of Drosophila melanogaster and D. simulans. Oecologia 40, 299–309 (1979).
Mele, S. et al. Drosophila as a diet discovery tool for treating amino acid disorders. Trends Endocrinol. Metab. 34, 85–105 (2023).
Lichter-Konecki, U. & Vockley, J. Phenylketonuria: Current treatments and future developments. Drugs 79, 495–500 (2019).
Knerr, I. et al. Expanding the genetic and phenotypic spectrum of branched-chain amino acid transferase 2 deficiency. J. Inherit. Metab. Dis. 42, 809–817 (2019).
Holeček, M. Branched-chain amino acids in health and disease: metabolism, alterations in blood plasma, and as supplements. Nutr. Metab. 15, 33 (2018).
Lee, P. T. et al. A gene-specific T2A-GAL4 library for Drosophila. Elife 7, 1–24 (2018).
Wu, J.-Y. et al. ENU mutagenesis identifies mice with mitochondrial branched-chain aminotransferase deficiency resembling human maple syrup urine disease. J. Clin. Invest. 113, 434–440 (2004).
Abreu-Blanco, M. T., Verboon, J. M., Liu, R., Watts, J. J. & Parkhurst, S. M. Drosophila embryos close epithelial wounds using a combination of cellular protrusions and an actomyosin purse string. J. Cell Sci. 125, 5984–5997 (2012).
Stoessel, D. et al. Metabolomics and lipidomics reveal perturbation of sphingolipid metabolism by a novel anti-trypanosomal 3-(oxazolo[4,5-b]pyridine-2-yl)anilide. Metabolomics 12, 126 (2016).
Creek, D. J. et al. Toward global metabolomics analysis with hydrophilic interaction liquid chromatography-mass spectrometry: Improved metabolite identification by retention time prediction. Anal. Chem. 83, 8703–8710 (2011).
Chambers, M. C. et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 30, 918–920 (2012).
Creek, D. J., Jankevics, A., Burgess, K. E. V., Breitling, R. & Barrett, M. P. IDEOM: An excel interface for analysis of LC–MS-based metabolomics data. Bioinformatics 28, 1048–1049 (2012).
Scheltema, R. A., Jankevics, A., Jansen, R. C., Swertz, M. A. & Breitling, R. PeakML/mzMatch: A file format, java library, r library, and tool-chain for mass spectrometry data analysis. Anal. Chem. 83, 2786–2793 (2011).
Pang, Z. et al. MetaboAnalyst 5.0: narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 49, W388–W396 (2021).
Linford, N. J., Bilgir, C., Ro, J. & Pletcher, S. D. Measurement of lifespan in Drosophila melanogaster. J. Vis. Exp. https://doi.org/10.3791/50068 (2013).
Acknowledgements
We thank Bloomington Drosophila Stock Center fly stocks, Ozdros for stock importation, Christen Mirth, Richard Burke, and members of the Johnson and Piper labs for helpful discussions. This work is supported by a National Health and Medical Research Council Ideas grant (APP1182330) to MDWP and TKJ and National Institutes of Health grant (5U01HG007530-08) to TKJ. TKJ is an Australian Research Council Future Fellow (FT220100023). JL and SM are supported by Australian Government Research Training Program Scholarships.
Author information
Authors and Affiliations
Contributions
F.M., A.Q., M.W., C.K.B. and T.K.J. performed the experiments. F.M., S.M., J.L., T.L.F., C.K.B. and R.S. analysed the data. F.M. generated all figures with assistance from J.L. F.M., T.K.J. and M.D.W.P. interpreted the data and wrote the manuscript. T.K.J. and M.D.W.P. conceived, acquired funding, and supervised the study. All authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Martelli, F., Quig, A., Mele, S. et al. A defined diet for pre-adult Drosophila melanogaster. Sci Rep 14, 6974 (2024). https://doi.org/10.1038/s41598-024-57681-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-57681-z
Keywords
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.

{kind=link}
{kind=link}
{kind=link}