
Abstract
Flowers cluster at various spatial scales, so pollinators use information from multiple scales when foraging in natural plant populations. Little is known about the effects of interactions between scales or their relative strength. We examined bumblebee foraging behaviour in a natural population of Salvia nipponica in 10 and 7 patches in 2019 and 2020, respectively. We recorded within-patch factors (display size of racemes and local open raceme densities) and patch-level factors (patch size and distance from the nearest patch) and analysed their relationships with pollinator behaviour. The numbers of visits per raceme and flower were mainly affected by the interaction of patch size and raceme density; they were higher in locations with lower raceme density in larger patches. The ratio of flowers visited to all open flowers in a raceme during a raceme visit, which relates to a bumblebee’s choice to leave a raceme, was mainly affected by the interaction of display size and local open raceme density; in 2019 it was higher in racemes with smaller display sizes, while in 2020 the strength and direction of the relationship depended on the open raceme density. These results suggest that pollinators relied on the sizes of flower clusters at different spatial scales when visiting and leaving racemes and adjusted their responses to the sizes of flower clusters depending on the distances between clusters. Therefore, it is important to evaluate factors at various spatial scales and their interactions to fully understand pollinator behaviour in natural plant populations.
Similar content being viewed by others

Introduction
Pollinator foraging behaviour is important for fitness of pollinators and animal-pollinated plants and hence many studies have examined factors affecting foraging behaviour1,2,3,4. These factors include floral display size (number of simultaneously opening flowers on a plant or an inflorescence)1,4,5,6,7,8,9, local plant (or inflorescence) density around a focal plant4,6,7,10,11,12,13, patch size (size of plant clusters within populations, where a population is defined such that the distances between populations are long and the populations are almost isolated from each other in terms of pollination)9,10,11, and distance between patches2,14,15,16,17. Using signals from these factors, pollinators may make decisions about visiting and leaving plants and patches. Optimal foraging theory18 is based on cost/benefit analyses where obtaining resources such as nectar is a benefit and energy spent foraging is a cost; and it predicts that pollinators should visit and leave plants as to optimize the benefit/cost ratio19,20.
Understanding how pollinators evaluate costs and benefits would increase our understanding of how pollinators decide to visit and then leave a flower, plant, or patch. Previous studies have examined the effects of individual signals on pollinator behaviour. (1) Floral display size and patch size: in general, pollinator visitation rate to a plant (or an inflorescence) increases with increasing floral display size1,4,5,6,7,8,9, possibly because larger patches provide more rewards. Similarly, the visitation rate to a patch2,9,14 and the residence time (the length of time a pollinator remains in a patch)16 increase with increasing patch size. However, the proportion of visited flowers decreases with increasing inflorescence or patch size2,3,4, possibly because of the cost of revisiting already-visited flowers21. (2) Local plant density: higher plant density should decrease cost by decreasing distance between plants. In general, pollinators visit more flowers on a plant in sparse areas4,6,7,10,22,23. On the other hand, the visitation rate per plant is higher in dense areas, potentially increasing benefits at higher plant density4,6,7,11,12,13,24,25,26,27,28. (3) Distance between patches: lastly, increasing distance between patches increases foraging time and cost of movement when pollinators move between patches20.
In natural plant populations, flowers can be grouped at various spatial scales; on inflorescences or racemes within a plant; on a plant cluster within a patch; or in patches with multiple plants. In such heterogenous conditions, pollinators may simultaneously evaluate multiple signals and may assign different levels of importance to different signals. Moreover, different signals may interact in how they affect pollinators. This could be interpreted as pollinators using interactive information obtained from multiple signals. For example, pollinators tend to visit more flowers on larger inflorescences or patches although they tend to visit a smaller proportion of the available flowers10,19. This behaviour led to the concept of ideal free distribution (IFD) of pollinators, i.e., a constant visitation rate on each flower irrespective of the size of flower clusters2,19. The key assumptions of the IFD are that the cost of movement is negligible, and foragers are omniscient about resource distributions. If these assumptions are violated, distribution of pollinators is predicted to deviate from the IFD20,29,30. The cost of movement and the perception of resource distribution may depend on the distance between clusters of resources20,30. Therefore, the size of flower clusters (e.g., floral display and patch size) and the distance between them (e.g., local density and distance between patches) may interact in affecting pollinator behaviour, although no interaction between the effects of patch size and distance between patches on pollinator behaviour was observed in a previous study2. Moreover, because the cost of movement would depend on the spatial scale, local density within a patch and distance between patches may also interact in affecting pollinator visitation rates.
Previous studies have studied the effects of single factors or only a few factors on pollinator behaviour: population (not patch) sizes and average plant densities in the populations31,32,33,34,35,36, local plant density (density of flowering plants around a focal plant)1,6,7,8,12,13,23,25,26,27,37,38,39,40,41,42,43,44,45, or patch size2,16,46. Dauber et al.47 focused on both patch density and size, but the effects of distance between patches and display sizes of individual plants were not examined. Mustajärvi et al.28 studied pollinator behaviour in artificial patches that included two levels of density and size, but studies with more variation in density and size of patches are required because pollinator decision making may become more complicated in more complex foraging sites, possibly including non-linear interactive effects of density and size. In addition, though the effects of distance between patches have been examined15,17,48, interactions of distance between patches with other patch characteristics are poorly understood (but see Fragoso et al.14, which showed that bees used both patch size and distance between patches when selecting a patch).
The purpose of this study is to determine the interactive effects and relative importance of flower clusters in different spatial scales on pollinator behaviour. Therefore, we examined bumblebee foraging behaviour in a natural population of Salvia nipponica. We focused on four factors representing cost and benefit for pollinators: local open raceme density (density of racemes having at least one open flower), patch size, patch distance, and floral display size (number of open flowers on a raceme). We then elucidated their effects and relative strengths, including the interactions among them, on three measures of pollinator behaviour: the number of visits to a raceme, the number of visits to a flower, and the proportion of visited flowers in a raceme.
Materials and methods
Plant species
Salvia nipponica Miq. (Lamiaceae) is a perennial herb that grows on the forest floor in the Japanese mainland. Each plant produces one or a few racemes (inflorescences), and each raceme bears up to 50 flowers. The yellow and lip-shaped flowers are adichogamous (i.e., stamens and pistils of a flower ripen at the same time), and each flower has 4 ovules. This species is self-compatible but needs pollinators for seed production3. Most flowers last only one day, and up to 15 flowers open on a raceme on one day. The mean ± SD number of open flowers of each observed raceme was 3.6 ± 1.7. In our study site, flowering occurs from early September through early October. The primary pollinator is Bombus diversus Smith. They visit the flowers mainly for nectar, but sometimes collect pollen3. Flowers of S. nipponica have a special stamen structure which allows only compatibly-sized pollinators (i.e., bumblebees in our study site) to contribute to their pollination3. Other visitors (including small bees, bee flies, and hover flies) do not perform effective pollination (though they try to collect nectar or pollen, their bodies fail to touch anthers, stigmas, or both, and therefore we did not examine them in this study. S. nipponica also reproduces clonally by tuberous roots or creating new roots from stems touching the ground, although it usually does not spread over long distances via clonal reproduction. Because the underground structure and history of clonal reproduction are unknown, we regarded an aboveground cluster of shoots, which might have several racemes, as a ramet (individual plant).
Study site and patches
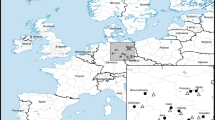
Field work was conducted on the forest floor in two adjacent areas in Kongosawa and Mt. Kagitori National Forests, Miyagi Prefecture, northern Honshu, Japan (Fig. 1). The plant patches selected for this study were on flat areas or gentle slopes in continuously distributed mixed deciduous and coniferous forests dominated by tree species such as Abies firma, Quercus serrata, and Pinus densiflora, or in an adjacent Cryptomeria japonica forest. The elevations of the patches were about 80–120 m. Although there were a few other nectar sources for bumblebees, such as Impatiens textorii and Tricyrtis affinis, around some patches, flowers of S. nipponica were much more abundant than those other nectar sources. The under-canopy vegetation and environmental conditions looked similar among the patches. We selected patches of wild S. nipponica plants of various sizes. We defined a patch as a group of ramets in which the distances between the ramets were within 5 m because 5 m distances were enough to distinguish patches and would prevent the same genets being assigned to different patches since the species does not spread over long distances. Therefore, the minimum distance between the patches was more than 5 m.

A map of patches studied. The study patches were on flat places or gentle slopes in continuously distributing mixed deciduous and coniferous forests dominated by tree species such as Abies firma, Quercus serrata, and Pinus densiflora or in an adjacent Cryptomeria japonica forest. The elevations of the patches were about 80–120 m and the under-canopy vegetation and environmental conditions looked similar. The study site is in two adjacent areas in Kongosawa and Mt. Kagitori National Forests, Miyagi Prefecture, northern Honshu, Japan.
We selected the patches studied as follows. In the middle of September in 2019, we recorded all flowering patches found from the walking trail network running through the study site. We then selected 10 patches so that their patch size distribution showed a wide range and each patch included racemes with multiple display sizes (Table 1; observed display sizes for each patch and year are shown in Supplementary Fig. S1d). We also took care to select patches distributed over a sufficiently small area to allow us to set video cameras and start video recording without large time lags. Many of the patches studied differed between 2019 and 2020 because, in 2020, flowering was early and had ended before we got to the patches or we could no longer find the patches studied in 2019 (Table 1). However, we studied patches nos. 4 and 10 in both years.
For each patch, we recorded the number of reproductive ramets (patch size). Locations of all the flowering patches were recorded by GPS and planar distances from the nearest patch were calculated for the patches used for the pollinator observations. The distances from the nearest patch were 20.5–483 m (Table 1), which is within the reported range of foraging area for bumblebee pollinators (estimated maximum foraging ranges are at least 449–758 m for four bumblebee species49).
Pollinator observations
Observations of pollinators were carried out on 5 and 7 sunny, warm days between 19 and 25 September 2019 and 16–28 September 2020, respectively (Supplementary Table S1). On each observation day, we selected 1 to 5 patches depending on their flowering conditions and locations within the study site; in particular in 2019, we tried to select northern (no. 1–3, 9, and 10) and southern (no. 4–8) patches on alternate days to minimize the possible effects of the difference in observation dates between the locations, while retaining simultaneous observation of nearby patches for convenience. In each patch, we selected 1–9 ramets and 1–11 racemes, aiming to observe racemes of various display sizes (ranging from 1 to 9 flowers) growing in patches of various sizes. In total, we recorded 5–31 racemes on one day and the total numbers of racemes observed were 123 and 56 in 2019 and 2020, respectively. The total number of ramets, racemes, and flowers used for pollinator observations for each year and patch are shown in Table 1 and the number of observed racemes for each observation day and patch are shown in Supplementary Table S1.
Using 4–9 video cameras (GZ-RX, JVCKENWOOD, Kanagawa, Japan), we recorded pollinator visits to each of the selected racemes for 180 min during 0900-1300, when pollinators were foraging. In both years, a raceme was only observed once. The total observation time was 216 h in 2019 and 168 h in 2020. We observed all open flowers on each selected raceme. Because the flowering phenology and the number of available racemes were different between the two years, 1 to 6 racemes belonging to the same ramets were simultaneously observed by single video cameras in 2019, whereas in 2020, single racemes were observed by single video cameras.
After the field observations, we recorded the number of open flowers (display size) for each selected raceme. A flower was classified as open if its corolla opening was wider than 7 mm, which allows pollinators to enter the flower. Ten racemes dropped one or two flowers during the video recording, so the field-measured numbers of open flowers were later adjusted for each pollinator visit using the video recordings for these racemes. Also, the number of other racemes having open flowers within a 1.5 m radius from the focal raceme (local open raceme density) was recorded after the field observations. We selected a 1.5 m radius because it reflected the local flowering condition of the focal raceme (variation in raceme density within this radius was small). When the focal raceme was at the edge of a patch, empty areas were included in the 1.5 m radius as it reflected the local flowering conditions. We used raceme density, instead of flower density, because the mean number of open flowers per raceme had little variation among patches.
Using the videos of the field observations, we recorded pollinator visits to racemes (raceme visits) and flowers (flower visits), and visited flower ratio. To count raceme visits, for each raceme observed, we counted the number of effective visits during the 3 h of observation. We regarded a visit as effective if a pollinator entered a flower and its body touched the anthers. Pollinator behaviour approaching flowers but not touching the anthers were not counted, and a revisit to a probed flower within a single bout was counted as a new visit. To count flower visits, for each flower within the raceme observed, we counted the number of effective visits during the 3 h of observation. Here, we counted multiple visits to a flower when pollinators re-visited the same flower during single visits to their racemes. To obtain the visited flower ratio, we recorded the numbers of visited and not visited open flowers in each raceme for each pollinator visit to determine the proportion of visited flowers to all open flowers in the raceme for each pollinator visit. Note that the visited flower ratio could be related to decision making about leaving racemes because this measurement involves the number of flowers not visited.
Seed development
We marked all the flowers observed, including both pollinated and not pollinated ones, after pollinator observation. Seeds developed to mature size 2–3 weeks after pollination, and we could classify seeds as matured by their sizes. When seeds matured, we counted the number of seeds developed for each flower of each raceme studied to examine the effects of pollinator visits on seed production. We measured the basal stem diameter of the ramet of each raceme, which was used as an index of the ramet resource status in many previous studies e.g., 50,51,52,53 and possibly affects seed production. Also, individuals of S. nipponica having larger diameters tend to have longer stems (Fig. S2). We determined the seed development ratio (the number of seeds developed/4, where 4 is the number of ovules) for each flower of each raceme.
Data analysis
For all statistical analyses, we applied a generalized linear mixed model (GLMM) using R version 4.3.1 statistical software54 with the lme4 package55. Before modelling, patch size and distance from the nearest patch were log-transformed to weaken collinearity among explanatory variables (Pearson’s correlation coefficients between patch size and distance from the nearest patch before and after log-transformation were 0.79 and 0.48, respectively). After log-transformation, all values of the variance inflation factor (VIF) for all analyses were less than 2.6 (Supplementary Table S2) and hence the collinearity should not have large effects on the analyses. Because there were no evident differences in the measured variables between the north and south patches (Fig. 1, Supplementary Fig. S1), we analysed the data obtained from the north and south patches together.
To examine factors affecting pollinator preferences in visiting racemes, we developed a model in which number of raceme visits (number of pollinator visits to a raceme during its observation) was the response variable. The explanatory variables of the model were floral display size, local open raceme density, patch size, and distance from the nearest patch. Because these explanatory variables might have interactive effects on the response variable, two-way interaction terms of combinations of the four explanatory variables were also included. In addition, to consider yearly differences in the response variable and in the relationships between the explanatory variables including the two-way interactions and the response variable, observation year and interaction terms between year and other explanatory variables were also included. For the patch-level explanatory variables (patch size and distance to the nearest patch), the patch value was assigned to all racemes in the patch. For the number of open flowers, the initial value of each focal raceme at the beginning of pollinator observation was used. For this model, the Poisson error distribution with the log-link function was used.
To examine factors affecting pollinator preferences in visiting flowers, we developed a model in which the number of flower visits (number of pollinator visits to a flower including revisits during its observation) was the response variable. This model used the same set of explanatory variables and interactions as the model above. For the patch-level explanatory variables (patch size and distance from the nearest patch), the patch value was assigned to all flowers in the patch. For the local open raceme density, the value of each focal raceme was assigned for all flowers within it. For the number of open flowers, the value of the raceme bearing the focal flower when the first pollinator visit was observed was used. Note that if no pollinator visits were observed for a flower, the initial number of open flowers of the raceme bearing the flower was used. For the error distribution, the Poisson error distribution with the log-link function was used.
To examine factors affecting pollinator decisions about staying on a raceme or leaving it, we developed models in which the visited flower ratio among the open flowers in a raceme in one pollinator bout was the response variable. For this analysis, the data obtained in the first three bouts observed was used for each raceme and later bouts were discarded to avoid possible effects of reduction in nectar amount. The same explanatory variables used in the models described above were also used for this model. Similarly, the patch-level explanatory variables were assigned to all racemes in the patch. For the number of open flowers in the visited flower ratio model, the value of the focal raceme at the time of visitation was used. For the error distribution, the binomial error distribution with the logit-link function was used.
To avoid estimation failure and to compare relative effects of each explanatory variable, all explanatory variables (display size, local open raceme density, patch size, and distance from the nearest patch) were standardized to have mean = 0 and standard deviation = 1 using the scale function. The standardization was done for each dataset for each analysis after assigning patch-level and raceme-level variables to racemes and flowers so that the standardization did not affect the results of the analyses.
To control the potential dependencies in pollinator behaviour recorded within the same observation days, patches, ramets and racemes, all models included observation day, patch, ramet and raceme as random effects. However, models including all the random effects occasionally produced singular fits. Because there was no consensus about how to deal with singularity55 (please see the documentation of ‘isSingular’ in the lme4 package), we followed the method proposed by Matuschek et al.56 and selected random terms using step-wise likelihood ratio tests with α = 0.2 to avoid singularity. Using the resultant models, the significance of the explanatory variables was tested by type II Wald χ2 tests using the Anova function in the car package57 because the significance of estimated coefficients using z statistics produced by the summary function depends on correspondence of the dummy variable for the year term.
To find which aspects of pollinator behaviour were important for seed production, a GLMM testing relationship between pollinator behaviour and seed development was implemented. The response variable of the model was the seed development ratio of a flower, and the explanatory variables were the number of raceme visits, the average visited flower ratio of the raceme where the flower was located, and the number of flower visits. In addition, basal stem diameter of the ramet of each raceme was included as an explanatory variable which might represent the resource status of individual plants. Because seed development and the relationships between the explanatory variables and seed development could be different between 2019 and 2020, observation year and two-way interaction terms between each explanatory variable and year were also used as explanatory variables. All explanatory variables were standardized using the same method as the other models. To control for potential similarities in seed set within the same patches, ramets, or racemes, the model included patch, ramet, and raceme as random effects. Also, to control potential overdispersion in the response variable, flower was also included as a random effect. After implementing the model with all the random effects, the same method of model selection and hypothesis testing used in the other models was applied.
Results
Pollinators observed
The most frequent visitor was Bombus diversus Smith, the only species effective for pollination. This bumblebee visited several flowers in single racemes and represented the entirety of bumblebee visitations for observed racemes. These pollinator conditions were consistent for both years.
Number of raceme visits
We recorded 419 and 575 pollinator visits to racemes in 2019 and 2020, respectively. The mean frequency of visits per raceme in 2020 was about twice as high as in 2019; 3.42 ± 2.45 and 10.26 ± 5.09 visits/3 h (mean ± SD) in 2019 and 2020, respectively.
The model selection procedure kept ramet identity as the sole random factor for the final model of the raceme visits (SD of the random factor = 0.245). The number of raceme visits increased with increasing number of open flowers in a raceme and was higher in 2020 (Fig. 2; Supplementary Table S3). Significant interactions were found between the local open raceme density and patch size, patch size and distance from the nearest patch, and the local open raceme density and year (Supplementary Table S3). The patch size and open raceme density had interactive effects on raceme visits. In both years, the number of raceme visits was higher in larger patches only when the local open raceme density was low and in 2020 the trend was reversed if local raceme density was high (Fig. 3A and B). On the other hand, local raceme density had positive effects on the number of raceme visits in small patches whereas it had negative effects in large patches in both years (Fig. 3C and D). The patch size and distance from the nearest patch had interactive effects on raceme visits (Fig. 4). The number of raceme visits was higher in larger patches only when the nearest patch was closer (Fig. 4A and B). On the other hand, distance from the nearest patch had negative effects for large patches (Fig. 4C and D).

The predicted relationship between the number of raceme visits and the standardized number of open flowers in a raceme in 2019 and 2020. The lines represent the predicted relationships, and the points represents partial residuals. Values of other explanatory variables in the model (i.e., local open raceme density, patch size, and distance from the nearest patch) were fixed at their means when calculating the predictions. Also, because we found significant interaction between year and local raceme density (Appendix S4), the difference between the 2 years could depend on local raceme density.

The interactive effect of patch size and local raceme density on the number of raceme visits in 2019 (A and C) and 2020 (B and D). Lines represent predicted relationships between the standardized patch size (A and B) or local raceme density (C and D) and the number of raceme visits when another explanatory variable is mean – 1 SD (dotted), mean (solid), and mean + 1 SD (dashed). The points represent partial residuals and their colours and symbols show the range of another explanatory variable (circle: \(x \leqq -0.5\); triangle: \(-0.5<x<0.5\); cross: \(x\geqq 0.5\)). Lines are drawn for the same range of another explanatory variable to avoid extrapolation. Values of other explanatory variables which are not shown in the panels were fixed at their means when calculating the predictions. Note that log-transformation was applied for patch size before standardization.

The interactive effect of patch size and distance from the nearest patch on the number of raceme visits in 2019 (A and C) and 2020 (B and D). Lines represent predicted relationships between the standardized patch size (A and B) or distance from the nearest patch (C and D) and the number of raceme visits when another explanatory variable is mean−1 SD (dotted), mean (solid), and mean + 1 SD (dashed). The points represent partial residuals and their colours and symbols show the range of another explanatory variable (circle: \(x \leqq -0.5\); triangle: \(-0.5<x<0.5\); cross: \(x\geqq 0.5\)). Lines are drawn for the same range of another explanatory variable to avoid extrapolation. Log-transformation was applied for patch size and distance from the nearest patch before standardization.
The observed number of raceme visits showed a wider range of responses depending on patch size and local open raceme density (Fig. 3; predicted lines ranged from ca. 2 to 8 in 2019 and from ca. 8 to 18 in 2020) than on the number of open flowers (Fig. 2; it ranged from ca. 3 to 6 in 2019 and from ca. 10 to 13 in 2020) and the interaction between distance from the nearest patch and patch size (Fig. 4; it ranged from ca. 2 to 5 in 2019 and from ca. 8.5 to 10 in 2020).
Number of flower visits
The model selection procedure kept ramet identity as the sole random factor for the final model of the flower visits (SD of the random factor = 0.370). For the number of flower visits, significant interactions were found between local open raceme density and year, patch size and year, local open raceme density and patch size, and patch size and distance from the nearest patch (Supplementary Table S4). In both years, the number of flower visits was higher in larger patches but only when local open raceme density was low (Fig. 5A and B). In 2020, patch size had a negative effect on number of flower visits when local raceme density was high (Fig. 5B). Local open raceme density generally had a negative effect on the number of flower visits in 2019 (Fig. 5C). In contrast, it had negative effects in large patches whereas it had positive effects in smaller patches in 2020 (Fig. 5D). In addition, the number of flower visits was higher in larger patches only if they were closer to other patches in both years (Fig. 6A and B). In both years, flowers in distant patches received smaller numbers of visits in large patches (Fig. 6C and D).

The interactive effect of patch size and local raceme density on the number of flower visits in 2019 (A and C) and 2020 (B and D). Lines represent predicted relationships between the standardized patch size (A and B) or local raceme density (C and D) and the number of flower visits when another explanatory variable is mean −1 SD (dotted), mean (solid), and mean + 1 SD (dashed). The points represent partial residuals and their colours and symbols show the range of another explanatory variable (circle: \(x \leqq -0.5\); triangle: \(-0.5<x<0.5\); cross: \(x\geqq 0.5\)). Lines are drawn for the same range of another explanatory variable to avoid extrapolation. Log-transformation was applied for patch size before standardization.

The interactive effect of patch size and distance from the nearest patch on the number of flower visits in 2019 (A and C) and 2020 (B and D). Lines represent predicted relationships between the standardized patch size (A and B) or distance from the nearest patch (C and D) and the number of flower visits when another explanatory variable is mean −1 SD (dotted), mean (solid), and mean + 1 SD (dashed). The points represent partial residuals and their colours and symbols show the range of another explanatory variable (circle: \(x \leqq -0.5\); triangle: \(-0.5<x<0.5\); cross: \(x\geqq 0.5\)). Lines are drawn for the same range of another explanatory variable to avoid extrapolation. Log-transformation was applied for patch size and distance from the nearest patch before standardization.
Visited flower ratio among the open flowers in a raceme
The model selection procedure kept raceme identity as the sole random factor for the final model of the visited flower ratio (SD of the random factor = 0.319). The number of flowers effectively visited in a raceme during a single visit (not counting revisits to the same flowers) was 1.54 ± 0.93 (mean ± SD) in 2019 and 1.93 ± 1.30 in 2020. Significant three-way interactions on their effects on the visited flower ratio were found among the number of open flowers, local open raceme density, and year, and among patch size, distance from the nearest patch, and year (Supplementary Table S5). The visited flower ratio was higher in racemes with smaller display sizes whereas local open raceme density had a negligible effect in 2019, and the interaction between display size and raceme density was not strong (Fig. 7A and C). On the other hand, the visited flower ratio was higher in racemes with smaller display sizes if the racemes had higher local open raceme density whereas it was slightly higher in racemes with larger display sizes if the racemes had lower raceme density in 2020 (Fig. 7B). Local raceme density had a positive effect on the visited flower ratio for smaller racemes whereas the opposite relationship was observed for larger racemes (Fig. 7D). Distance from the nearest patch and patch size did not have strong effects on the visited flower ratio in 2019 (Fig. 8A and C) whereas in 2020 it was higher in larger patches that were closer to other patches and smaller patches that were distant from other patches (Fig. 8B and D).

The interactive effect of the number of open flowers and local open raceme density on the visited flower ratio in 2019 (A and C) and 2020 (B and D). Lines represent predicted relationships between the standardized number of open flowers (A and B) or local open raceme density (C and D) and the visited flower ratio when another explanatory variable is mean − 1 SD (dotted), mean (solid), and mean + 1 SD (dashed). The points represent partial residuals and their colours and symbols show the range of another explanatory variable (circle: \(x \leqq -0.5\); triangle: \(-0.5<x<0.5\); cross: \(x\geqq 0.5\)). Lines are drawn for the same range of another explanatory variable to avoid extrapolation.

The interactive effect of patch size and distance from the nearest patch on the visited flower ratio in 2019 (A and C) and 2020 (B and D). Lines represent predicted relationships between the standardized number of patch size (A and B) or distance from the nearest patch (C and D) and the visited flower ratio when another explanatory variable is mean − 1 SD (dotted), mean (solid), and mean + 1 SD (dashed). The points represent partial residuals and their colours and symbols show the range of another explanatory variable (circle: \(x \leqq -0.5\); triangle: \(-0.5<x<0.5\); cross: \(x\geqq 0.5\)). Lines are drawn for the same range of another explanatory variable to avoid extrapolation. Log-transformation was applied for patch size and distance from the nearest patch before standardization.
The observed visited flower ratio showed wider responses depending on the number of open flowers and local open raceme density (estimated slope in Fig. 7; it ranged from ca. 0.15 to 0.75 in 2019 and from ca. 0.3 to 0.75 in 2020) than on patch size and distance (Fig. 8; it ranged from ca. 0.45 to 0.58 in 2019 and from ca. 0.35 to 0.6 in 2020). In particular, the response to the number of open flowers was very strong in 2019.
Seed development ratio
The model selection procedure kept ramet and flower identities as the random factors for the final model of the seed development ratio (SD of the random factor; ramet ID: SD = 0.783; flower ID: SD = 0.570). Among the observed aspects of pollinator behaviour, the interaction between the number of raceme visits and year had the strongest effect on the seed development ratio (Supplementary Tables S6 and S7), though the p value was slightly higher than 0.05; the seed development ratio tended to increase with the number of raceme visits in 2019 but tended to decrease with it in 2020 (Fig. 9).

The predicted relationship between the standardized number of raceme visits and seed development ratio in 2019 and 2020. The lines represent the predicted relationships for the two years, and the points represent partial residuals.
Discussion
Pollinator behaviour was affected by the floral display size and the local flowering conditions including the local raceme density, patch size, and distance from the nearest patch. We found several consistent effects of the local flowering conditions in both years, though there were differences between the 2 years.
Decision making using multiple signals
Raceme visits
The number of raceme visits was affected by both display size (Fig. 2) and interactions between patch level variables (Figs. 3 and 4). Patch size and local open raceme density had larger effects on raceme visits than the number of open flowers (compare changes along the variables in Fig. 3 with Fig. 2). This suggests that the decision making of pollinators about visiting racemes relies more on patch size and local density than on display size. Maybe this is because the benefit of an entire patch (patch size) or the cost of movement among racemes (local density) have larger effects on the foraging efficiency of pollinators than the benefit of individual racemes (raceme size). Also, no significant interaction between display size and patch level variables was detected (Supplementary Table S3). This suggests that pollinators rely on the display size independent of the other patch level signals; maybe visiting racemes having large display sizes improves foraging efficiency of pollinators in any patch level conditions.
The number of raceme visits increased with increasing number of open flowers in the raceme (Fig. 2), consistent with many previous studies1,4,5,6,7,8,9. However, the local flowering conditions also affected the number of raceme visits; one of the consistent results in both years was that the effect of local raceme density on raceme visits depended on patch size: it was higher in the locations with lower local raceme density in larger patches (Fig. 3). The local raceme density may be important in large patches compared to small patches because sub-structures of raceme distributions within patches may be more important in large patches, where the cost of within-patch movement is large, than in small patches. Mustajärvi et al.28 also reported that pollinator visits per plant were higher in sparse populations. However, many other studies6,7,12,13,27 reported that pollinator visits per plant were high in dense areas. Perhaps the interactive effect of patch size and local raceme density we showed can explain the difference in the density dependence of pollinator visits among these studies.
The number of raceme visits was also affected by the interaction between patch size and distance from the nearest patch though the effect was smaller than other explanatory variables (Fig. 4): it was higher in larger patches that are closer to other patches in both years (Fig. 4). This might be because patches within a certain distance are subjects for patch selection; for pollinators that have left a patch, large patches soon found are more attractive compared to small patches soon found, resulting in higher raceme visits in nearby, large patches. Thus, such decisions may be in agreement with previous studies showing frequent visits of pollinators in larger2,15 or closer2 patches. Thus, pollinators may select patches providing higher benefits with lower search costs. On the other hand, no obvious trend was found in the relationship between the number of raceme visits and patch size in patches distant from others (Fig. 4A and B). This suggests that pollinators do not selectively visit patches after long trips from previously visited patches because further trips are costly and there is no assurance of finding better patches.
Flower visits
The number of flower visits showed a similar response to the local flowering conditions as the number of raceme visits; the consistent results in both years were that the number of flower visits was higher in locations with lower local raceme density in larger patches (Fig. 5), and in larger patches that were closer to other patches (Fig. 6). This may be simply because pollinators visit more flowers while frequently visiting racemes.
According to the IFD, the proportions of foraging animals will equal the rates of resource production58. For pollinators, increasing number and decreasing proportion of visited flowers with increasing size of flower clusters often bring equal frequency of pollinator visits to each flower irrespective of cluster size and produce IFD of pollinators2,19. However, our results showed scale-dependent deviation from the IFD; although there was no relationship between raceme display size and flower visits, flower visits were higher in larger patches when the focal patches were close to other patches (Fig. 6). This might be because the cost of movement between clusters of flowers and incomplete knowledge of resource distribution, which are neglected in the original IFD model and may increase with distance, are more significant at larger spatial scales. On the other hand, the observed pattern was different from the prediction of the revised IFD model incorporating suboptimal movement due to incomplete discriminability of benefit, distance-dependent cost of movement, and information uncertainty of resource distribution20. In the revised model, foragers were predicted to use smaller resource patches more frequently than the prediction of the original IFD model when forager movement is suboptimal. Under this condition, increasing information uncertainty shifted the distributions of foragers to more frequent use of smaller patches whereas increasing cost of movement shifted the distribution closer to the prediction of IFD. However, because bumblebees used flowers in larger patches more frequently (Fig. 6), the observed pattern did not match the prediction of the model. Another possibility which could explain the observed pattern would be differences in nectar production in different sized patches; larger patches could be located in favorable environmental conditions and flowers in larger patches could produce more nectar per flower. Although nectar production of S. nipponica does not depend on display size of inflorescences3, its dependence on patch size is unknown. Measuring nectar production in different sized patches may help to understand potential mechanisms underlying the scale-dependent deviation from IFD.
Visited flower ratio
The visited flower ratio may be based on decision making about staying on or leaving racemes, noting that this measurement involves the number of flowers not visited. This ratio was affected both by the number of open flowers and the studied patch conditions, but the number of open flowers and local open raceme density had stronger effects than patch size and distance (compare Fig. 7 and Fig. 8). This suggests that, in the decision making about leaving racemes, pollinators rely more strongly on the benefit of the current raceme and the cost of movement to adjacent racemes than on the total benefit of the current patch or the cost of arrival to the current patch. In addition, interactive effects on the visited flower ratio were found (Figs. 7 and 8, Supplementary Table S5). This suggests that pollinators determine when to leave racemes by balancing several factors relating the costs and benefits rather than relying on a single factor.
The visited flower ratio was higher in racemes with lower numbers of open flowers in 2019 (Fig. 7A). This result agrees with the prediction that, as pollinators have limited memory, pollinators forage in higher proportions of flowers in plants with smaller display sizes because of low probabilities of revisiting already-foraged flowers21. On the other hand, the interactive effect of the number of open flowers and local open raceme density was more evident in 2020: the visited flower ratio was slightly higher in racemes with larger numbers of open flowers in lower local open raceme densities, in contrast to the result in 2019, whereas it was higher in racemes with smaller numbers of open flowers in higher raceme densities, as in 2019 (Fig. 7B). These results in 2020 are consistent with a previous study reporting that the proportion visited (number of visited flowers in a bout/display size) was higher for larger display sizes in sparse patches and was higher for the smallest display sizes in dense patches7. The difference between the years might be due to pollinators being more abundant in 2020 compared to 2019. In 2020, there might have been more flowers previously foraged by other pollinators, reflecting increasing competition among pollinators. Under such conditions, pollinators may need to adjust their departure timing based on the number of open flowers in the current raceme relative to the cost of movement to adjacent racemes (i.e., local density). Note that because there were only two patches observed in both years, the difference between the years might be partly confounded by the spatial difference between the patches.
Although their effects were weaker than the other variables, patch sizes and distances also affected the visited flower ratio; it was higher in larger and distant patches in 2019 though the effect of distance alone was negligible among the observed patches (Fig. 8A and C), whereas it was higher both in smaller distant patches and in larger patches that were closer to other patches in 2020 (Fig. 8B and D). Note that the number of raceme visits also depended on patch size and distance from the nearest patch in a manner similar to the visited flower ratio; it was higher in larger patches slightly closer to other patches in 2019 whereas it was higher in smaller and distant patches and larger patches that are closer to other patches in 2020 (Fig. 4). Therefore, one possible explanation for the observed patterns is that pollinators adjusted their foraging behaviour in response to competition with other pollinators, which should be strong if the number of raceme visits is large. In patches with higher numbers of raceme visits, the benefit of movement to other racemes would decrease and the benefit of staying on the current raceme would increase because the probability of visiting already pollinated racemes would be higher. Therefore, the visited flower ratio of pollinators would be higher in patches with higher numbers of raceme visits.
Effects of pollinator behaviour on seed production
The interaction between the number of raceme visits and year had the strongest effect on the seed development ratio, a measurement of effective pollen deposition (Supplementary Table S6); it tended to increase with the number of raceme visits in 2019 but tended to decrease with it in 2020 (Fig. 9). These contrasting results might be due to greater abundance of pollinators in 2020 than in 2019; it is possible that raceme visits enhanced seed production simply due to enhanced pollen deposition in 2019, whereas pollen deposition was saturated for seed production in 2020. However, it is unclear why raceme visits had negative effects on seed production in 2020. Miyake and Sakai1 showed that larger racemes of S. nipponica received higher numbers of raceme visits and geitonogamy, but outcrossing rates were still higher for larger racemes in the same study site. Therefore, the negative effect might not be caused by inbreeding depression by geitonogamy. Other factors such as resource depletion by nectar consumption and style damage by pollinators59 might be responsible for this negative effect.
Conclusion
In this study, we found several interactive effects of display size and patch conditions on pollinator behaviour. Thus, in a natural population of flowering plants where patchy sub-structure causes uneven distribution of floral resources, pollinators use multiple signals representing the costs and benefits of raceme- and patch-level conditions and adjust their responses to a certain signal in association with other signals. Moreover, pollinators use different information signals when visiting and leaving racemes; pollinators may use patch-level information when visiting racemes, but raceme-level information when leaving racemes. These results emphasize the importance of evaluating the effects of multiple factors and their interactions on several aspects of pollination behaviour to fully understand optimal foraging strategies of pollinators in natural populations.
Data availability
All data and programs are available at Dryad: https://doi.org/10.5061/dryad.0vt4b8h14
References
Miyake, Y. C. & Sakai, S. Effects of number of flowers per raceme and number of racemes per plant on bumblebee visits and female reproductive success in Salvia nipponica (Labiatae). Ecol. Res. 20, 395–403 (2005).
Ishihama, F. & Washitani, I. Behavior of queen bumblebee pollinators on Primula sieboldii (Primulaceae) in response to different patch sizes and spacing. Plant Species Biol. 22, 167–174 (2007).
Ohashi, K. Consequences of floral complexity for bumblebee-mediated geitonogamous self-pollination in Salvia nipponica Miq. (Labiatae). Evolution 56, 2414–2423 (2002).
Ohashi, K. & Yahara, T. Visit larger displays but probe proportionally fewer flowers: Counterintuitive behaviour of nectar-collecting bumble bees achieves an ideal free distribution. Funct. Ecol. 16, 492–503 (2002).
Mitchell, R. J., Karron, J. D., Holmquist, K. G. & Bell, J. M. The influence of Mimulus ringens floral display size on pollinator visitation patterns. Funct. Ecol. 18, 116–124 (2004).
Klinkhamer, P. G. L. & de Jong, T. J. Effects of plant size, plant-density and sex differential nectar reward on pollinator visitation in the protandrous Echium vulgare (Boraginaceae). Oikos 57, 399–405 (1990).
Grindeland, J. M., Sletvold, N. & Ims, R. A. Effects of floral display size and plant density on pollinator visitation rate in a natural population of Digitalis purpurea. Funct. Ecol. 19, 383–390 (2005).
Makino, T. T., Ohashi, K. & Sakai, S. How do floral display size and the density of surrounding flowers influence the likelihood of bumble bee revisitation to a plant?. Funct. Ecol. 21, 87–95 (2007).
Akter, A., Biella, P. & Klecka, J. Effects of small-scale clustering of flowers on pollinator foraging behaviour and flower visitation rate. PLoS ONE 12, e0187976 (2017).
Goulson, D. Why do pollinators visit proportionally fewer flowers in large patches?. Oikos 91, 485–492 (2000).
Kunin, W. E. Population size and density effects in pollination: Pollinator foraging and plant reproductive success in experimental arrays of Brassica kaber. J. Ecol. 85, 225–234 (1997).
Feldman, T. S. Pollinator aggregative and functional responses to flower density: Does pollinator response to patches of plants accelerate at low-densities?. Oikos 115, 128–140 (2006).
Weber, A. & Kolb, A. Local plant density, pollination and traitfitness relationships in a perennial herb. Plant Biol. 15, 335–343 (2013).
Fragoso, F. P., Jiang, Q., Clayton, M. K. & Brunet, J. Patch selection by bumble bees navigating discontinuous landscapes. Sci. Rep. 11, 8986 (2021).
McCormick, M. L., Aslan, C. E., Chaudhry, T. A. & Potter, K. A. Benefits and limitations of isolated floral patches in a pollinator restoration project in Arizona. Restor. Ecol. 27, 1282–1290 (2019).
Cresswell, J. E. & Osborne, J. L. The effect of patch size and separation on bumblebee foraging in oilseed rape: Implications for gene flow. J. Appl. Ecol. 41, 539–546 (2004).
Delmas, C. E. L., Escaravage, N. & Pornon, A. Massive floral display affects insect visits but not pollinator-mediated pollen transfer in Rhododendron ferrugineum. Plant Biol. J. 16, 234–243 (2014).
MacArthur, R. H. & Pianka, E. R. On optimal use of a patchy environment. Am. Natural. 100, 603–609 (1966).
Dreisig, H. Ideal free distributions of nectar foraging bumblebees. Oikos 72, 161–172 (1995).
Matsumura, S., Arlinghaus, R. & Dieckmann, U. Foraging on spatially distributed resources with sub-optimal movement, imperfect information, and travelling costs: Departures from the ideal free distribution. Oikos 119, 1469–1483 (2010).
Ohashi, K. & Yahara, T. How long to stay on, and how often to visit a flowering plant?: A model for foraging strategy when floral displays vary in size. Oikos 86, 386 (1999).
Pyke, G. H. Foraging in bumblebees: Rule of departure from an inflorescence. Can. J. Zool.-Rev. Can. Zool. 60, 417–428 (1982).
Jing, X., Hong, L. & Rui, Q. Unexpectedly high outcrossing rate in both dense and sparse patches in self-compatible Pedicularis rex (Orobanchaceae). Plant System. Evol. 299, 49–56 (2013).
Pyke, G. H. Optimal foraging in bumblebees: Rule of movement between flowers within inflorescences. Anim. Behav. 27, 1167–1181 (1979).
Nielsen, A. & Ims, R. A. Bumble bee pollination of the sticky catchfly in a fragmented agricultural landscape. Ecoscience 7, 157–165 (2000).
Bosch, M. & Waser, N. M. Effects of local density on pollination and reproduction in Delphinium nuttallianum and Aconitum columbianum (Ranunculaceae). Am. J. Bot. 86, 871–879 (1999).
Field, D. L., Ayre, D. J. & Whelan, R. J. The effect of local plant density on pollinator behavior and the breeding system of Persoonia bargoensis (Proteaceae). Int. J. Plant Sci. 166, 969–977 (2005).
Mustajärvi, K., Siikamäki, P., Rytkönen, S. & Lammi, A. Consequences of plant population size and density for plant-pollinator interactions and plant performance: Plant-pollinator interactions. J. Ecol. 89, 80–87 (2001).
Åström, M. Travel cost and the ideal free distribution. Oikos 69, 516–519 (1994).
Kennedy, M. & Gray, R. D. Can ecological theory predict the distribution of foraging animals? A critical analysis of experiments on the ideal free distribution. Oikos 68, 158–166 (1993).
Bernhardt, C. E., Mitchell, R. J. & Michaels, H. J. Effects of population size and density on pollinator visitation, pollinator behavior, and pollen tube abundance in Lupinus perennis. Int. J. Plant Sci. 169, 944–953 (2008).
Mayer, C., Michez, D., Chyzy, A., Brédat, E. & Jacquemart, A.-L. The abundance and pollen foraging behaviour of bumble bees in relation to population size of whortleberry (Vaccinium uliginosum). PLoS One 7, e50353 (2012).
Klank, C., Pluess, A. R. & Ghazoul, J. Effects of population size on plant reproduction and pollinator abundance in a specialized pollination system. J. Ecol. 98, 1389–1397 (2010).
Somme, L., Mayer, C. & Jacquemart, A.-L. Multilevel spatial structure impacts on the pollination services of Comarum palustre (Rosaceae). PLoS ONE 9, e99295 (2014).
Kirchner, F. et al. Effects of local density on insect visitation and fertilization success in the narrow-endemic Centaurea corymbosa (Asteraceae). Oikos 111, 130–142 (2005).
Waites, A. R. & Ågren, J. Pollinator visitation, stigmatic pollen loads and among-population variation in seed set in Lythrum salicaria: Population size and pollination. J. Ecol. 92, 512–526 (2004).
Heinrich, B. Resource heterogeneity and patterns of movement in foraging bumblebees. Oecologia 40, 235–245 (1979).
Schmitt, J. Flowering plant density and pollinator visitation in Senecio. Oecologia 60, 97–102 (1983).
Bosch, M. & Waser, N. M. Experimental manipulation of plant density and its effect on pollination and reproduction of two confamilial montane herbs. Oecologia 126, 76–83 (2001).
Essenberg, C. J. Scale-dependent shifts in the species composition of flower visitors with changing floral density. Oecologia 171, 187–196 (2013).
Nattero, J., Malerba, R., Medel, R. & Cocucci, A. Factors affecting pollinator movement and plant fitness in a specialized pollination system. Plant Syst. Evol. 296, 77–85 (2011).
Kunin, W. E. Sex and the single mustard: Population-density and pollinator behavior effects on seed-set. Ecology 74, 2145–2160 (1993).
Singh, V. K., Barman, C., Mohanty, D. & Tandon, R. Relative contribution of reproductive attributes to the density-dependent effects on fruit-set. AoB PLANTS 10, 141 (2018).
Elliott, S. E. & Irwin, R. E. Effects of flowering plant density on pollinator visitation, pollen receipt, and seed production in Delphinium barbeyi (Ranunculaceae). Am. J. Bot. 96, 912–919 (2009).
Cibula, D. A. & Zimmerman, M. The effect of plant-density on departure decisions: Testing the marginal value theorem using bumblebees and Delphinium nelsonii. Oikos 43, 154–158 (1984).
Sih, A. & Baltus, M. S. Patch size, pollinator behavior, and pollinator limitation in catnip. Ecology 68, 1679–1690 (1987).
Dauber, J. et al. Effects of patch size and density on flower visitation and seed set of wild plants: A pan-European approach. J. Ecol. 98, 188–196 (2010).
Zimmerman, M. Optimal foraging, plant density and the marginal value theorem. Oecologia 49, 148–153 (1981).
Knight, M. E. et al. An interspecific comparison of foraging range and nest density of four bumblebee (Bombus) species. Mol. Ecol. 14, 1811–1820 (2005).
Hendrix, S. D. Compensatory reproduction in a biennial herb following insect defloration. Oecologia 42, 107–118 (1979).
Hendrix, S. D. Reactions of Heracleum lanatum to floral herbivory by Depressaria pastinacella. Ecology 65, 191–197 (1984).
Wada, N. & Uemura, S. Size-dependent flowering behavior and heat production of a sequential hermaphrodite, Symplocarpus renifolius (Araceae). Am. J. Bot. 87, 1489–1494 (2000).
Iwaizumi, M. G. & Sakai, S. Variation in flower biomass among nearby populations of Impatiens textori (Balsaminaceae): Effects of population plant densities. Can. J. Bot. 82, 563–572 (2004).
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/ (2023).
Bates, D., Mächler, M., Bolker, B. & Walker, S. Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67, 48 (2015).
Matuschek, H., Kliegl, R., Vasishth, S., Baayen, H. & Bates, D. Balancing Type I error and power in linear mixed models. JMemL 94, 305–315 (2017).
Fox, J. & Weisberg, S. An R Companion to Applied Regression (Sage, 2019).
Fretwell, S. D. & Lucas, H. L. On territorial behavior and other factors influencing habitat distribution in birds. Acta Biotheor. 19, 16–36 (1969).
Aizen, M. A. et al. When mutualism goes bad: Density-dependent impacts of introduced bees on plant reproduction. New Phytol. 204, 322–328 (2014).
Acknowledgements
We thank Takuya Hasegawa, Saya Shinagawa, Ayaka Momi, and Hirotsugu Niijima for their help during the field work, Jane Glazebrook for English editing, and the three anonymous reviewers for their valuable comments and suggestions. The administrative boundary and the road edge shown in Fig. 1 were obtained from the Digital National Land Information provided by the National Land Information Division, National Spatial Planning and Regional Policy Bureau, Ministry of Land, Infrastructure, Transport and Tourism of Japan (https://nlftp.mlit.go.jp/ksj/) and the river data in Fig. 1 was obtained from the OpenStreetMap project.
Author information
Authors and Affiliations
Contributions
N.M., T.I., and S.S. designed the study and conducted the field work. N.M. and M.O. analysed the data. N.M., M.O., and S.S. wrote the first version of the manuscript. All authors approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Murakoshi, N., Itagaki, T., Oguro, M. et al. Effects of floral display size, local open raceme density, patch size, and distance between patches on pollinator behaviour in Salvia nipponica. Sci Rep 14, 967 (2024). https://doi.org/10.1038/s41598-024-51327-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-51327-w
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
