
Abstract
The mechanism of metachronous recurrence (MR) after performing endoscopic treatment for early gastric adenocarcinoma (GAC) and eradicating Helicobacter pylori (H. pylori) is unknown. To elucidate the mechanism and risk factors of MR, we analyzed gene expression at multiple locations of the gastric mucosa. We selected each five patients with MR and without MR (control), after early GAC treatment and eradication of H. pylori. Mucosal tissue was collected from four sites in the stomach of each patient as biopsy specimens for mRNA sequencing, gene set enrichment analysis, and microRNA (miRNA) sequencing. We also performed correlation analysis and target prediction on pathways. As a result, endoscopically, the MR group had more intestinal metaplasia and enlarged folds. A total of 384 mRNAs presented changes in expression and 31 gene sets were enriched in the MR group. Immune-related pathways were enriched in the entire stomach, and the IFN-α response had the highest enrichment score. Additionally, 32 miRNAs revealed changes in their expression. Correlation analysis and target prediction with genes in the gene set of IFN-α response revealed that 10 miRNA–mRNA pairs presented a significant correlation. Immune-related pathways with miRNAs in the gastric mucosa after H. pylori eradication may be a risk factor for MR.
Similar content being viewed by others


Introduction
Gastric cancer remains one of the most life-threatening diseases, causing over a million deaths annually worldwide1. Most gastric cancers (approximately 90–95%) are adenocarcinomas (gastric adenocarcinoma; GAC)2. With the spread of endoscopic technology, GAC is often detected in its early stages and endoscopic treatment is widely available, providing results comparable to those of surgery while maintaining patient quality of life by preserving the stomach. However, gastric cancer recurs in 5.2–14% of patients after endoscopic treatment, which is a major clinical problem3,4.
The main cause of GAC is mucosal damage caused by chronic infection with Helicobacter pylori (H. pylori), which is found in approximately 63% of gastric cancer cases5,6. The eradication of H. pylori reduces the risk of gastric carcinogenesis and prevents recurrence in patients after endoscopic resection7,8. Thus, endoscopic resection (endoscopic submucosal dissection and endoscopic mucosal resection) and eradication of H. pylori are generally performed in patients with early GAC with H. pylori infection9.
However, there are many cases of recurrence even after the eradication of H. pylori and endoscopic treatment of gastric cancer3,4. Metachronous recurrence (MR) of GAC can occur even when curative endoscopic resection for early GAC is performed and eradication of H. pylori is successful. Therefore, even after curative endoscopic resection of early GAC and eradication of H. pylori, almost all patients undergo regular post-treatment follow-up9. This causes patients to incur a large expense, and its contribution to the social cost also needs to be considered. Therefore, risk factors for MR after H. pylori eradication should be identified, and preventative methods should be developed.
In general, the risk of gastric carcinogenesis includes the degree of endoscopic atrophy in the gastric mucosa and the presence and extent of intestinal metaplasia10,11. This is also true for patients after H. pylori eradication and endoscopic treatment of early GAC12. However, the mechanism of this phenomenon has not yet been elucidated. Recently, genetic and epigenomic analyses have advanced with the widespread use of next-generation sequencers, and various reports on the molecular mechanisms of GAC have been published13,14,15.
MicroRNAs (miRNAs) are small non-coding RNAs that play an important role in post-transcriptional gene regulation in a variety of biological processes and have been reported to be involved in gastric carcinogenesis16,17. However, the gene expression changes, which occur in the gastric mucosa after H. pylori eradication and induce recurrence, along with miRNA changes, are unclear. Additionally, since the inflammation caused by H. pylori progresses heterogeneously in the gastric body, it is necessary to perform a wide range gene expression analysis of the gastric mucosa from each individual to understand the details of gene expression related to MR.
In this study, we performed an integrated analysis of microRNA (miRNA) and messenger RNA (mRNA) expression profiles of the gastric mucosa at multiple locations in patients with MR to elucidate the metachronous recurrence mechanism of GAC. The functions of the extracted genes were clarified by bioinformatics, and the expression profiles of miRNAs and mRNAs were compared with endoscopic findings and other clinical data to identify the regulatory factors of MR.
Material and methods
Patients
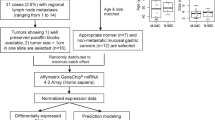
Adults aged ≥ 20 years, who had previously undergone curative endoscopic resection for early GAC at our hospital, Chiba University Hospital, Japan, and underwent esophagogastroduodenoscopy at our hospital between November 1, 2019 and February 29, 2020, were deemed eligible and enrolled accordingly. Curative endoscopic resection is defined pathologically as differentiated mucosal cancer, without ulceration or with ulceration with a maximum diameter of less than 30 mm, without lymphatic or venous invasion, and with negative horizontal and vertical margins9. Patients who had H. pylori successfully eradicated 1 year after their first GAC treatment, had open-type gastric mucosal atrophy according to the Kimura-Takemoto classification18, and had never undergone upper gastrointestinal surgery were enrolled (Fig. 1a). From these, we selected five patients that did (MR group) and did not (control group) have MR after successfully eradicating H. pylori.

Study flow chart (a) Patient eligibility criteria. (b) Biopsies were taken from four different locations: 1. the lesser curvature of the antrum (LA), 2. the greater curvature of the antrum (GA), 3. the lesser curvature of the body (LB), and 4. the greater curvature of the body (GB).
The MR group was defined as the group with patients who underwent curative endoscopic resection for GAC, presented no new GAC during endoscopic follow-up within six months, and presented newly developed GAC (differentiated cancer) in other parts of the stomach during endoscopic follow-up more than 12 months after treatment. In contrast, the control group was defined as the group with patients who did not have MR after more than four years of endoscopic follow-up after curative endoscopic treatment. GAC was finally diagnosed by pathological evaluation of endoscopically resected specimens.
The clinical characteristics, including age, sex, body mass index (BMI), drinking and smoking habits, the time interval between the first occurrence of GAC and biopsy, successful H. pylori eradication to biopsy, and history of GAC at the antrum or body of the patients, were investigated.
This study was reviewed and approved by the Institutional Review Board of the Chiba University School of Medicine. The purpose of this study was explained to all patients. Written and informed consent was obtained from all patients included in the study. All experiments were performed in accordance with relevant guidelines and regulations.
Evaluating endoscopic findings
Modified grading scores for the Kyoto Classification of Gastritis were performed to evaluate the mucosal condition. The Kyoto classification score for gastritis is based on the sum of scores of the five endoscopic findings: Atrophy (A), intestinal metaplasia (IM), enlarged folds (H), nodularity (N), and diffuse redness (DR), which ranges from 0 to 819. Although the IM score is usually determined by observation with white light imaging, we determined the IM score by observations with white light imaging and image-enhanced endoscopy, in a manner different from that of the Kyoto classification score. We summed the A, H, N, and DR scores and the modified IM score into a modified Kyoto classification score. This scoring was performed by three endoscopy experts who read the endoscopic images retrospectively.
Biopsy sites
In each case, mucosal tissue was collected as biopsy specimens from four non-neoplastic gastric mucosal sites during endoscopic follow-up. These four sites were defined by the updated Sydney system20 and other previous reports21. Specimens were obtained from the lesser curvature of the antrum (LA) and from the greater curvature of the antrum (GA), both within 2–3 cm from the pylorus, the middle portion of the lesser curvature of the body between the cardia and angle (LB), and the middle portion of the greater curvature of the corpus (GB) (Fig. 1b).
Extraction of RNA
After the biopsy, samples were placed in a microtube containing All Protect Tissue Reagent (Qiagen, Venlo, Netherlands), stabilized, and stored at -80 °C. Total RNA was extracted using the miRNeasy Micro Kit (Qiagen). Quality of total RNA was assessed before miRNA / mRNA sequencing by the protocol of the analysis service.
Integral analysis for miRNA and mRNA
The extracted total RNA was sequenced for miRNA using the DNBSEQ-G400 (MGI Tech Co., Ltd, Shenzhen, CHINA) and mRNA using the Hiseq 2500 (Illumina, San Diego, CA, USA) by GENEWIZ Japan (Tokyo, JP) (Fig. 2). The Deseq2 package version 1.34.0 was used for expression analysis, and the BiomaRt package was used for mRNA annotation22. Gene set enrichment analysis (GSEA) was used for functional analysis of mRNAs, with the hallmark gene sets from the Molecular Signature Database (MSigDB) 7.423. TargetScan 8.0 was used for miRNA target prediction24. Gene expression information was annotated to the predicted miRNAs and target genes, and the correlation between them was calculated. All authors had access to the study data and had reviewed and approved the final manuscript.

Analysis flowchart MR, metachronous recurrence; DEA, differential expression analysis; GSEA, gene set enrichment analysis; NOM p val, nominal p-value.
CIBERSORT
CIBRESORT, a computational approach, was used to analyze the micro-immune environment within the gastric mucosa. Transcriptome data of all samples were analyzed according to the CIBRESORT analysis algorithm to evaluate the dynamics of the 22 kinds of immune cells (naïve B cells naive, memory B cells, plasma cells, CD8+ T cells, naïve CD4+ T cells, resting memory CD4+ T cells, activated memory CD4+ T cells, follicular helper T cells, regulatory T cells, gamma delta T cells, resting NK cells, activated NK cells, monocytes, M0 macrophages, M1 macrophages, M2 macrophages, resting dendritic cells, activated dendritic cells, resting mast cells, activated mast cells, eosinophils, and neutrophils) in the gastric mucosa25.
Statistics
The Wald test was used for gene expression analysis, and the Pearson test was used for correlation analysis using R version 4.1.226. The statistical program SPSS version 21.0 (SPSS Inc., Chicago, IL, USA) was used for statistical analysis of clinical characteristics and endoscopic findings. The chi-square test, Fisher's exact test, and Mann–Whitney U test were used, and a p-value of less than 0.05 was considered statistically significant.
Results
Clinical characteristics and endoscopic grading
A total of five patients with MR and five patients without MR (as controls) were enrolled in this study according to the criteria (Fig. 1a). There were no significant differences in the clinical background between the groups (Table 1). On evaluation by the modified grading scores for the Kyoto classification of gastritis, the MR group had higher scores of IM (intestinal metaplasia) and H (enlarged folds) and a higher overall score than the control (Supplementary Fig. 1).
mRNA profiles of gastric mucosa from patients with MR
In the mRNA expression profiling, total RNAs of gastric mucosa from subjects were examined. All samples measured the A260 /A280 and RNA integrity number (RIN) for quality check [A260 / 280 (median (range)), 2.04 (1.91–2.10); RIN, 7.50 (4.90–9.10))] by the protocol of the analysis service.
FastQC were performed to check the quality of row sequence data by the analysis service. The peak of the average of quality scores was greater than 30, indicating that the sequence data of all samples were highly evaluated. Then, we filtered out from the total sequence data (60,612 rows) those rows that contained gene sequences unregistered in the ensemble gene ID and low-expressed genes for which the total value of all samples was less than 100 reads. The 20,584 mRNAs of sequence data obtained by this filtering were statistically analyzed.
A total of 384 mRNAs revealed expression changes in the comparison of the gastric mucosa at all four sites; 303 revealed an increased expression and 81 revealed a decreased expression in patients with MR compared to the control [adjusted p-value (adj-p) < 0.05, |Log2 fold change| (|LFC|) > 1].
In the greater curvature of the gastric body, 889 mRNAs were differentially expressed; 771 were upregulated and 118 were downregulated in patients with MR compared to the control. In the greater curvature of the antrum, 311 mRNAs revealed changes in their expression; 298 were upregulated and 13 were downregulated in patients with MR compared to the control. In the lesser curvature of the gastric body, three mRNAs revealed changes in their expression; two were upregulated and one was downregulated in patients with MR compared to the control. In the lesser curvature of the antrum, one mRNA was upregulated in patients with MR compared to that in controls.
Hierarchical clustering analysis demonstrated that patients with MR and controls can be generally classified according to the expression patterns of 384 genes that revealed changes in their expression in all four gastric mucosae. Similarly, the expression patterns of 899 genes from mucosae in the greater curvature of the gastric body and 311 genes from mucosae in the greater curvature of the gastric antrum were used to classify patients with MR and controls (Fig. 3).

mRNA profiles and IFN-α response in GSEA (a-c) Heatmap of differentially expressed mRNAs by hierarchical clustering analysis (adjusted p-value < 0.05, |Log2 fold change|> 1). The following was analyzed: (a) all sites combined, (b) greater curvature of the body (GA), and (c) greater curvature of the body (GB). MR-1–5 are patients of the MR (metachronous recurrence) group. N-1–5 are patients of the control (non-MR) group. (d) Enrichment plot of IFN-α response at the analysis of all sites combined, which was the top hit at GSEA. (e) Expression of genes in the gene set of IFN-α response. Panels (a-c) were made using R version 4.1.2 and Pretty Heatmaps package. Panels (d-e) were generated using GSEA software (https://www.gsea-msigdb.org/gsea/index.jsp).
Enrichment analysis for mRNA and predicted target genes
Gene set enrichment analysis (GSEA) was performed to elucidate the biological significance of the mRNA expression associated with metachronous recurrence of GAC. In the analysis of all sites, 31 gene sets, which were enriched in patients with MR, and four gene sets, which were dominant in the controls, were extracted (Table 2).
Similarly, GSEA was performed for all sites, and enrichment gene sets were extracted for patients with MR and controls. The interferon-alpha (IFN-α) response gene set had the highest enrichment score at all sites and was extracted as the top hit. The IFN-α response gene set contained 97 mRNAs, of which 26 presented significant expression changes in the expression variation analysis.
miRNA profiles of gastric mucosa from patients with MR
FastQC were performed to check the quality of row sequence data by the analysis service. The peak of the average of quality scores was greater than 30, indicating that the sequence data of all samples were highly evaluated. Then, we filtered out from the total sequence data (3,553 rows) those rows that low-expressed genes for which the total value of all samples was less than 100 reads. The 545 rows of miRNA sequence data obtained by this filtering were statistically analyzed.
To understand the regulatory system of gene expression associated with metachronous recurrence of GAC, RNA sequencing was used to profile miRNA expression. In a comparison using the gastric mucosa at all four sites, a total of 32 miRNAs presented expression changes; 13 presented an increased expression and 19 presented a decreased expression in patients with MR compared to the control [adj-p < 0.05, |LFC|> 1]. In the greater curvature of the gastric body, 13 miRNAs were differentially expressed; four were upregulated and nine were downregulated in patients with MR compared to the control. In the greater curvature of the antrum, one miRNA was upregulated in patients with MR compared to the control. In the lesser curvature of the gastric body, no miRNA presented changes in the gene expression. In the lesser curvature of the antrum, 14 miRNAs presented gene expression changes; 9 were upregulated and 5 were downregulated in patients with MR compared to the control.
Hierarchical clustering analysis revealed that patients with MR and control can be generally classified according to the expression patterns of 32 miRNAs that presented expression changes in all four gastric mucosae (Fig. 4). Similarly, the expression patterns of 13 miRNAs in the greater curvature of the gastric body and 14 genes in the lesser curvature of the gastric antrum were used to classify patients with MR and controls.

miRNA profiles Heatmap of differentially expressed miRNAs by hierarchical clustering analysis (adjusted p-value < 0.05, |Log2 fold change|> 1). Each of the following was analyzed: (a) all sites combined, (b) LA, and (c) GB. MR-1–5 are patients of the MR (metachronous recurrence) group, and N-1–5 are patients of the control (non-MR) group. Panels (a-c) were made using R version 4.1.2 and Pretty Heatmaps package. LA, Lesser curvature of the antrum; GA, greater curvature of the antrum; LB, lesser curvature of the body; GB, greater curvature of the body.
Correlation analysis of miRNAs presenting changes in expression to IFN-α response
To elucidate the transcriptional regulation of the IFN-α response in gastric mucosa, the correlation between 26 genes in the IFN-α response gene set and 32 differentially expressed miRNAs was analyzed. Accordingly, 435 mRNA-miRNA pairs were correlated (|r|> 0.5, p < 0.05, Fig. 5a). Among these pairs, 206 presented a negative correlation and 229 presented a positive correlation between differentially expressed mRNAs and miRNAs. Of the 206 pairs, 10 pairs were predicted to be target gene pairs from the database and were negatively correlated with each other (Fig. 5b). The five pairs (miR-135a-5p/CASP1, miR-340-5p/B2M, miR-29b-2-5p/IFIT2, miR-204-5p/PSMB9, miR-135a-5p/CXCL10, and miR-135a-5p/TMEM140) presented particularly strong negative correlations (r < − 0.6, p < 0.05).

Correlation between mRNA and miRNA in IFN-α response (a) The correlation between 26 genes in the IFN-α response gene set and 32 differentially expressed miRNAs (|r|> 0.5, p < 0.05). (b) Correlation diagram of the mRNA-miRNA pairs that revealed a negative correlation and were predicted to be target gene pairs from the database: The central figure demonstrates miRNA and mRNA expression levels and regression line, and the figures above and to the right of the center demonstrate box plot of the expression levels of miRNA and mRNA in MR and control groups, respectively.
Characteristics of immune microenvironment in the gastric mucosa from patients with MR
The microenvironment in background gastric mucosa of the MR group was analyzed using CIBERSORT to predict immunocytes associated with continued IFN-α/γ response. Seven of the 22 immune cell populations showed significant differences between the MR and control groups (p < 0.05, Supplementary Fig. 2). Four immunocytes (follicular helper T cells (p = 0.0013), gamma delta T cells (p = 0.0028), activated NK cells (p = 0.0054), and M1 macrophages (p = 0.0029)) were significantly increased in the MR group compared to the control group. Three immunocytes (CD8 + T cells (p = 0.01), regulatory T cells (p = 0.0072), and mast cells (p = 0.0003)) were significantly decreased in the MR group when compared with that in the control group.
Discussion
We performed an integrated and comprehensive analysis of endoscopic findings and miRNA and mRNA profiles of the gastric mucosa of MR patients after eradication of H. pylori. It showed that the gastric mucosa of MR patients shows enlarged folds and specific mRNA and miRNA profiles within the tissue. Then, pathway analysis showed that immune-related pathway, such as IFN-α signaling pathway, were altered in the gastric mucosa of MR patients. Furthermore, miRNAs correlated with the expression of mRNAs in the IFN-α signaling pathway and were predicted to target these mRNAs.
First, inflammatory response such as IFN-α, IFN-γ, TNF-α, and IL-6 were elevated in MR patients. In previous reports, these inflammatory cytokines were also found in the blood of gastric cancer patients and were thought to be associated with chronic inflammation due to H. pylori infection.27. However, all patients with MR in this study had been eradicated of H. pylori. Therefore, it is very interesting that these inflammatory cytokine-associated molecules are upregulated in our patients with MR. Furthermore, endoscopic findings revealed enlargement of the greater curvature folds in patients with MR, even after H. pylori eradication. Enlarged folds are known to be a risk factor for gastric carcinogenesis, especially in patients with current H. pylori infection28, however usually improve shortly after H. pylori eradication29. Thus, enlarged folds after eradication of H. pylori suggests the possibility of a prolonged immune response in the gastric mucosa, which would normally be suppressed by eradication, and supports our mRNA profiling. It is an important issue for the further study to search for the cause of the inflammatory reaction in the gastric mucosa of MR patients after eradication of H. pylori. However, the aim of this study was to perform an integrated miRNA–mRNA analysis, and it was difficult to add sufficient samples for causal validation.
Representatively, we focused on the "IFN-α response" gene set, which was the top hit in GSEA. This gene set contained 27 differentially expressed mRNAs. The relationship of those 27 mRNAs to gastric cancer was discussed based on previous reports. Caspase-1 (CASP-1) is an inflammasome induced by type 1 interferon and regulates the secretion of cytokines and chemokines; IL-1β is a typical cytokine regulated by CASP-1, whose overexpression has been reported to induce gastric inflammation and carcinogenesis30,31. In addition, CASP-1 is reported to have both pro-inflammatory and regulatory effects in H. pylori infection32 and may be involved in the persistence of inflammation after eradication. Chemokine (C-X-C motif) ligand 10 (CXCL10) has been implicated in the promotion of gastric cancer invasion and carcinogenesis33,34. Moreover, CXCL10 is also reported to be induced by type I interferons35. Beta 2-microglobulin (B2M) binds to human leukocyte antigen (HLA) class 1, a transmembrane protein, and HLA expression in gastric cancer has been implicated in prognosis36.
To predict the biological functions of 32 MR-specific miRNAs, we analyzed the correlation between IFN-α-related genes and their expression. Interestingly, the expression of miRNAs and IFN-α-related genes revealed a strong correlation. Thus, we hypothesize that some of the MR-specific miRNAs are involved in the regulation of IFN-α-related genes. Particularly, miR-29b-2-5p, miR-135a-5p, miR-204-5p, and -340-5p have predicted target genes in IFN-α related genes and are strongly correlated with them. miR-135a-5p targets CASP1, CXCL10, and transmembrane protein 140 (TMEM140), which are known to be involved in the proliferation of gastric cancer cells and the expression of IFN-α. miR-135a-5p reportedly suppresses gastric cancer cell proliferation and migration in gastric cancer tissues37,38. miR-340-5p targets beta 2-microglobulin, albeit reportedly involved in the regulation of gastric cancer development and apoptosis39,40. These findings suggest that miRNAs with variable expression may be regulators of MR-induced chronic inflammation.
Additionally, along with these inflammation-related molecules, the p53-related gene cluster also presented changes, suggesting that oncogenic genetic changes also occur in the gastric mucosa of patients with MR.
On the other hand, the CIBERSORT analysis showed that some immunocytes in the gastric mucosa of the MR group fluctuated. It was reported that follicular helper T cells, one of the immunocyte increased in the MR group in our study, contributed to the continuation of interferon-γ-mediated chronic inflammation and carcinogenesis in the gastric mucosa infected with H. pylori41. Similarly, various associations between groups of immunocytes that exhibit fluctuations and gastric cancer have been reported42,43,44. While this study cannot clarify how those fluctuations are related to the enhanced IFN-α/γ response in the MR group and to gastric cancer recurrence, it is an important finding that elucidates the mechanism of gastric cancer recurrence.
In order to accurately elucidate the relationship of these IFN-α-related mRNAs and miRNAs to MR, a longitudinal analysis using a larger number of specimens is necessary. In this study, four gastric mucosal biopsies were taken from individuals, which pose a high patient burden. Therefore, it is desirable to verify these findings in a renewed study design.
In this study, we found enlargement of the greater curvature folds with upregulation of immune-related molecules even after eradication of H. pylori in MR patients. We hypothesize that these changes indicate ongoing inflammation of the gastric mucosa in MR patients, and that this ongoing inflammation induces carcinogenesis.
Data availability
RNA-seq data for this study have been deposited in the Gene Expression Omnibus (GEO) under accession number GSE190459.
References
Wong, M. C. S. et al. Global incidence and mortality of gastric cancer, 1980–2018. JAMA Netw. Open. 4, e2118457. https://doi.org/10.1001/jamanetworkopen.2021.18457 (2021).
Rawla, P. & Barsouk, A. Epidemiology of gastric cancer: Global trends, risk factors and prevention. Prz. Gastroenterol. 14, 26–38. https://doi.org/10.5114/pg.2018.80001 (2019).
Nasu, J. et al. Characteristics of metachronous multiple early gastric cancers after endoscopic mucosal resection. Endoscopy. 37, 990–993. https://doi.org/10.1055/s-2005-870198 (2005).
Nakajima, T. et al. Metachronous gastric cancers after endoscopic resection: How effective is annual endoscopic surveillance?. Gastric. Cancer. 9, 93–98. https://doi.org/10.1007/s10120-006-0372-9 (2006).
Uemura, N. et al. Helicobacter pylori infection and the development of gastric cancer. N. Eng. J. Med. 345, 784–789. https://doi.org/10.1056/NEJMoa001999 (2001).
Parkin, D. M. The global health burden of infection-associated cancers in the year 2002. Int. J. Cancer. 118, 3030–3044. https://doi.org/10.1002/ijc.21731 (2006).
Ford, A. C., Yuan, Y. & Moayyedi, P. Helicobacter pylori eradication therapy to prevent gastric cancer: Systematic review and meta-analysis. Gut. 69, 2113–2121. https://doi.org/10.1136/gutjnl-2020-320839 (2020).
Lee, Y. C. et al. Association between Helicobacter pylori eradication and gastric cancer incidence: A systematic review and meta-analysis. Gastroenterology. 150, 1113-1124.e5. https://doi.org/10.1053/j.gastro.2016.01.028 (2016).
Japanese Gastric Cancer Association. Japanese gastric cancer treatment guidelines 2018. 5th ed. Gastric Cancer 2021;24(1):1−21
Sekikawa, A. et al. Gastric atrophy and xanthelasma are markers for predicting the development of early gastric cancer. J. Gastroenterol. 51, 35–42. https://doi.org/10.1007/s00535-015-1081-0 (2016).
Reddy, K. M., Chang, J. I., Shi, J. M. & Wu, B. U. Risk of gastric cancer among patients with intestinal metaplasia of the stomach in a US integrated health care system. Clin. Gastroenterol. Hepatol. 14, 1420–1425. https://doi.org/10.1016/j.cgh.2016.05.045 (2016).
Kaji, K. et al. Grading of atrophic gastritis is useful for risk stratification in endoscopic screening for gastric cancer. Am. J. Gastroenterol. 114, 71–79. https://doi.org/10.1038/s41395-018-0259-5 (2019).
Cancer Genome Atlas Research Network. Comprehensive molecular characterization of gastric adenocarcinoma. Nature. 513, 202–209. https://doi.org/10.1038/nature13480 (2014).
Sakuta, K. et al. Somatic alterations and mutational burden are potential predictive factors for metachronous development of early gastric cancer. Sci. Rep. https://doi.org/10.1038/s41598-020-79195-0 (2020).
Asada, K. et al. Demonstration of the usefulness of epigenetic cancer risk prediction by a multicentre prospective cohort study. Gut. 64, 388–396. https://doi.org/10.1136/gutjnl-2014-307094 (2015).
Song, J. H. & Meltzer, S. J. MicroRNAs in pathogenesis, diagnosis, and treatment of gastroesophageal cancers. Gastroenterology. 143, 35-47.e2. https://doi.org/10.1053/j.gastro.2012.05.003 (2012).
Ishimoto, T. et al. Current perspectives toward the identification of key players in gastric cancer microRNA dysregulation. Int. J. Cancer. 138, 1337–1349. https://doi.org/10.1002/ijc.29627 (2016).
Kimura, K. & Takemoto, T. An endoscopic recognition of the atrophic border and its significance in chronic gastritis. Endoscopy. 1, 87–97. https://doi.org/10.1055/s-0028-1098086 (1969).
Haruma K, Kato M, Inoue K, Murakami K, Kamada T. Kyoto classification of gastritis. 1st ed. Tokyo: Nihon Medical Center; (2017)
Dixon, M. F., Genta, R. M., Yardley, J. H. & Correa, P. Classification and grading of gastritis: The updated Sydney system. he Am. J. Surg. Pathol. 20(10), 1161–1181. https://doi.org/10.1097/00000478-199610000-00001 (1996).
Satoh, K. et al. Biopsy sites suitable for the diagnosis of Helicobacter pylori infection and the assessment of the extent of atrophic gastritis. Am. J. Gastroenterol. 93, 569–573. https://doi.org/10.1111/j.1572-0241.1998.166_b.x (1998).
Durinck, S., Spellman, P. T., Birney, E. & Huber, W. Mapping identifiers for the integration of genomic datasets with the R/Bioconductor package biomaRt. Nat. Protoc. 4, 1184–1191. https://doi.org/10.1038/nprot.2009.97 (2009).
Mootha, V. K. et al. PGC-1alpha-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet. 34, 267–273. https://doi.org/10.1038/ng1180 (2003).
Lewis, B. P., Shih, I. H., Jones-Rhoades, M. W., Bartel, D. P. & Burge, C. B. Prediction of mammalian microRNA targets. Cell. 115, 787–798. https://doi.org/10.1016/s0092-8674(03)01018-3 (2003).
Newman, A. M. et al. Robust enumeration of cell subsets from tissue expression profiles. Nat. Methods. 12, 453–457. https://doi.org/10.1038/nmeth.3337 (2015).
R Core Team. R: A Language and Environment for Statistical Computing; 2021. https://www.R-project.org/. Vienna, Austria: R Foundation for Statistical Computing
Sánchez-Zauco, N. et al. Circulating blood levels of IL-6, IFN-γ, and IL-10 as potential diagnostic biomarkers in gastric cancer: a controlled study. BMC Cancer. 17, 384. https://doi.org/10.1186/s12885-017-3310-9 (2017).
Nishibayashi, H. et al. Helicobacter pylori-induced enlarged-fold gastritis is associated with increased mutagenicity of gastric juice, increased oxidative DNA damage, and an increased risk of gastric carcinoma. J. Gastroenterol. Hepatol. 18, 1384–1391. https://doi.org/10.1046/j.1440-1746.2003.03192.x (2003).
Kato, M. et al. Changes in endoscopic findings of gastritis after cure of H. pylori infection: multicenter prospective trial. Dig. Endosc. 25, 264–273. https://doi.org/10.1111/j.1443-1661.2012.01385.x (2013).
Malireddi, R. K. & Kanneganti, T. D. Role of type I interferons in inflammasome activation, cell death, and disease during microbial infection. Front. Cell Infect. Microbiol. 3, 77 (2013).
Tu, S. et al. Overexpression of interleukin-1β induces gastric inflammation and cancer and mobilizes myeloid-derived suppressor cells in mice. Cancer Cell. 14, 408–419. https://doi.org/10.1016/j.ccr.2008.10.011 (2008).
Hitzler, I. et al. Caspase-1 has both proinflammatory and regulatory properties in helicobacter infections, which are differentially mediated by its substrates IL-1β and IL-18. J. Immunol. 188, 3594–3602. https://doi.org/10.4049/jimmunol.1103212 (2012).
Zhou, H., Wu, J., Wang, T., Zhang, X. & Liu, D. CXCL10/CXCR3 axis promotes the invasion of gastric cancer via PI3K/AKT pathway-dependent MMPs production. Biomed. Pharmacother. 82, 479–488. https://doi.org/10.1016/j.biopha.2016.04.069 (2016).
Chen, X., Chen, R., Jin, R. & Huang, Z. The role of CXCL chemokine family in the development and progression of gastric cancer. Int. J. Clin. Exp. Pathol. 13, 484–492 (2020).
Kader, M. et al. Interferon type I regulates inflammasome activation and high mobility group box 1 Translocation in hepatocytes during Ehrlichia-induced acute liver injury. Hepatol. Commun. 5(1), 33–51. https://doi.org/10.1002/hep4.1608 (2020).
Park, Y. et al. Clinicopathologic significance of human leukocyte antigen class I expression in patients with stage II and III gastric cancer. Cancer Immunol. Immunother. 68, 1779–1790. https://doi.org/10.1007/s00262-019-02410-z (2019).
Wu, H. et al. MiR-135a targets JAK2 and inhibits gastric cancer cell proliferation. Cancer Biol. Ther. 13, 281–288. https://doi.org/10.4161/cbt.18943 (2012).
Xie, Y., Li, F., Li, Z. & Shi, Z. miR-135a suppresses migration of gastric cancer cells by targeting TRAF5-mediated NF-κB activation. Onco. Targets Ther. 12, 975–984. https://doi.org/10.2147/OTT.S189976 (2019).
Yu, J. et al. miR-340 inhibits proliferation and induces apoptosis in gastric cancer cell line SGC-7901, possibly via the AKT pathway. Med. Sci. Monit. 23, 71–77. https://doi.org/10.12659/msm.898449 (2017).
Xiao, C. et al. miR-340 affects gastric cancer cell proliferation, cycle, and apoptosis through regulating SOCS3/JAK-STAT signaling pathway. Immunopharmacol. Immunotoxicol. 40, 278–283. https://doi.org/10.1080/08923973.2018.1455208 (2018).
Zhang, H. et al. Proinflammatory follicular helper T cells promote immunoglobulin G secretion, suppress regulatory B cell development, and correlate with worse clinical outcomes in gastric cancer. Tumour. Biol. 39, 1010428317705747. https://doi.org/10.1177/1010428317705747 (2017).
Wang, J. et al. Tumor-infiltrating γδT cells predict prognosis and adjuvant chemotherapeutic benefit in patients with gastric cancer. Oncoimmunology. 6, e1353858. https://doi.org/10.1080/2162402X.2017.1353858 (2017).
Saito, H., Takaya, S., Osaki, T. & Ikeguchi, M. Increased apoptosis and elevated Fas expression in circulating natural killer cells in gastric cancer patients. Gastric. Cancer. 16, 473–479. https://doi.org/10.1007/s10120-012-0210-1 (2013).
Yuan, X. L. et al. Gastric cancer cells induce human CD4+Foxp3+ regulatory T cells through the production of TGF-β1. World J. Gastroenterol. 17, 2019–2027. https://doi.org/10.3748/wjg.v17.i15.2019 (2011).
Author information
Authors and Affiliations
Contributions
H.O., K.O., and R.N. conceived the study. A.N. and H.O. collected and preserved biopsy specimen of gastric mucosa and extracted RNA. R.K. and R.N. analyzed the RNA data. N.A., K.O., and T.M. evaluated endoscopic findings. C.G., S.T., R.H., A.K., T.I., W.S., T.K., K.K., Y.O., T.T., and K.S. provided valuable discussions. A.N. and R.N. wrote the main manuscript text. T.C., J.K., N.K. administrated this project. All authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Nagashima, A., Okimoto, K., Nakagawa, R. et al. Investigation of risk factors for metachronous recurrence in patients with early gastric adenocarcinoma by miRNA–mRNA integral profiling. Sci Rep 13, 19661 (2023). https://doi.org/10.1038/s41598-023-47000-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-47000-3
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
